
硫醇转移酶基因敲除小鼠模型的建立及其白内障形成机制
2020-03-23 07:19:20MarjorieLou
国际眼科杂志 2020年3期
张 婕,严 宏,Marjorie F.Lou
0 引言
年龄相关性白内障可由多种因素诱发,包括氧化损伤、紫外线辐射、高血糖、吸烟及遗传等。氧化损伤是其中最重要的发病因素。研究证实,在年龄相关性白内障的晶状体中,含巯基的抗氧化物和谷胱甘肽(glutathione,GSH)水平急剧下降,含巯基的蛋白质广泛被氧化,形成蛋白质与GSH形成的二硫化物(protein-GSH mixed disulfides,PSSG)或蛋白质与蛋白质形成的二硫化物(protein-protein mixed disulfides,PSSP);细胞质和细胞膜间的大量结合蛋白转化为不可溶性蛋白;细胞膜内大部分脂质被氧化,以上这些都是导致白内障发生的原因[1]。
硫醇转移酶(thioltransferase,TTase)也称为谷氧还蛋白1(glutaredoxin 1,Grx1),它是巯基-二硫化物的氧化还原酶家族中一个非常重要的成员。1974年首次在大鼠肝细胞发现[2],随后在大肠杆菌E.coli中发现[3],并且得到广泛的研究。哺乳动物中Grx两种最主要的同工酶,TTase(Grx1)主要存在于细胞质中[4-5],Grx2存在于线粒体中[6],Grx2的功能尚未完全阐明。TTase通过断裂晶状体蛋白质氧化形成的二硫键(PSSG)使硫醇化的蛋白质脱硫醇,保持晶状体蛋白和膜蛋白的还原状态,从而阻止它们交联失活,同时可以通过调节酶活性中心的巯基来修复氧化损伤的酶,从而防止白内障的产生[4-5]。本研究通过建立硫醇转移酶基因敲除小鼠模型,观察其晶状体形态和生化方面随年龄的改变,探讨TTase在晶状体氧化还原系统中的生理作用及在年龄相关性白内障发生发展过程中的重要性。
1 材料和方法
1.1材料主要实验试剂:REDExtract-N-Amp Tissue PCR试剂盒、胎牛血清、GSH含量测定试剂盒(美国Sigma公司),BCA蛋白浓度测定试剂盒(美国Pierce公司),抗PSSG单克隆抗体(美国ViroGeng公司),抗GAPDH单克隆抗体、抗Beta-actin单克隆抗体、山羊抗鼠IgG2a(美国Santa Cruz公司),His-band试剂盒(美国QIAGEN公司),其他试剂均为国产分析纯试剂。主要仪器设备:裂隙灯(美国Zeiss公司)、离心机(美国Eppendorf公司),PCR仪、电泳仪、电转仪、化学发光成像仪(美国Bio-Rad公司),紫外分光光度计(美国Beckman Coulter公司)。
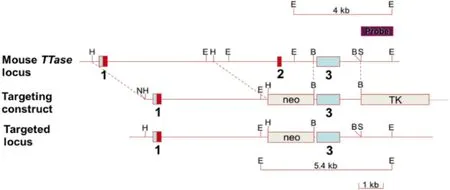
图1小鼠TTase基因敲除过程。
1.2方法
1.2.1建立TTase基因敲除小鼠模型TTase基因敲除过程见图1,该过程是与南京大学-南京生物医药研究院合作完成。小鼠TTase基因序列包括3个外显子,第1、2外显子为蛋白质编码区,且第2外显子为GSH结合部位,第3外显子为3’-非编码区。从129SV小鼠文库中克隆出TTase基因组片段,用外源性载体序列(Neo)替换,敲除片段包含了第2个外显子和第1、2内含子的部分片段。打靶载体用Not I酶切线性化后,转入R1胚胎干细胞[来源(129SVx129SVJ)F1],经过筛选,得到阳性克隆。将筛选成功的克隆细胞植入C57BL/6小鼠的囊胚细胞,进行至少两代遗传,由杂合子(TTase+/-)[遗传背景为(129SVx129SVJ)F1和C57BL/6]小鼠交配后产杂合子、野生型和基因敲除型三种基因型小鼠,采用PCR的方法对其基因型进行鉴定,最后得到TTase基因敲除型小鼠(TTase-/-)[7]。实验动物在清洁级层流动物房内进行饲养繁殖,小鼠笼、水、食物、垫料等均经消毒处理。采用1只公鼠和1只母鼠交配繁殖后代。母鼠的孕期大约是20d,生育小鼠发育良好。本研究过程中动物管理符合动物保护条例,并经伦理委员会批准。
1.2.2小鼠基因型鉴定采用REDExtract-N-Amp Tissue PCR试剂盒进行小鼠基因组DNA抽提,并采用PCR扩增的方法对所有小鼠基因型进行鉴定。
1.2.3裂隙灯观察小鼠晶状体混浊情况分别取不同年龄(1、4、9、12、16月龄)TTase基因敲除型和野生型小鼠,0.5%托吡卡胺和0.5%去氧肾上腺素眼液联合散瞳,15min后在裂隙灯下观察每只小鼠双眼晶状体混浊程度,并根据晶状体混浊分级系统II(LOCSII)[8]进行分级,分析不同基因型小鼠随年龄增长白内障发生情况。
1.2.4小鼠晶状体可溶性蛋白质的提取与定量小鼠处死后,立即取出眼球,分离晶状体并称重,置于干冰上冷冻干燥,-80℃保存待用。加组织裂解液于1~2mL匀浆器中,将晶状体置于匀浆器中匀浆,并置于冰上,重复研磨使组织尽量碾碎,裂解30min后,用移液管将裂解液移至1.5mL离心管中,在4℃下13000r/min离心10min,离心2次,取上清置于-80℃保存。采用BCA蛋白浓度测定试剂盒检测蛋白质的总浓度。
1.2.5小鼠晶状体GSH含量的测定取晶状体组织匀浆液于1.5mL离心管中,加等体积20%三醋酸纤维素(TCA)混合,使终浓度为10%,在4℃下13000r/min离心10min,离心2次,取上清备用。按试剂盒说明书测定晶状体GSH含量。
1.2.6 Western blot检测小鼠晶状体PSSG蛋白质的表达将提取的晶状体组织匀浆液进行12% SDS-PAGE凝胶电泳、转膜及脱脂牛奶封闭,之后于4℃冰箱内与一抗(1∶1000抗PSSG单克隆抗体)孵育过夜,室温漂洗后给予二抗(1∶4000 HRP标记的山羊抗鼠IgG2a)封闭液中孵育1h,再次漂洗后加入配置好的显色液,于发光成像仪Versadoc进行化学发光及成像并进行蛋白条带的光密度扫描。
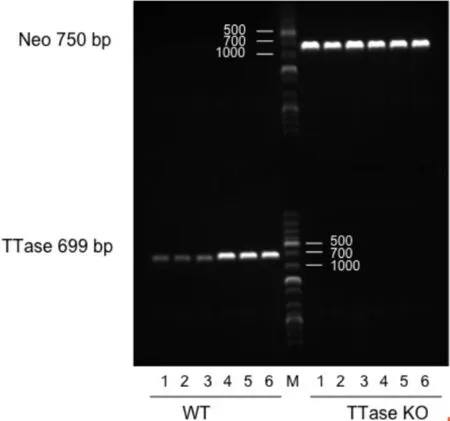
图2小鼠基因型PCR鉴定结果M:Marker;1~6:小鼠编号。WT:野生型;TTase KO:TTase基因敲除型。
1.2.7免疫共沉淀法检测形成PSSG的蛋白质每管中加入20μL晶状体组织匀浆液,加入RIPA缓冲液至100μL,加入5μg抗体(抗Beta-actin/GAPDH单克隆抗体),在混匀器上4℃匀速旋转过夜。每管中加入50μL Beta-actin/GAPDH beads,在混匀器上4℃匀速旋转2h。将免疫沉淀后的溶液于4℃ 2500r/min离心10min,收集上清,加入500μL RIPA缓冲液洗涤beads,4℃ 2500r/min离心10min,弃上清,共洗涤3次。在洗涤完毕的beads中加入2×SDS加样缓冲液,于100℃煮沸3min后进行Western blot检测(方法同前)。
1.2.8重组人晶状体TTase的分离纯化及脱硫醇作用检测选取转化的携有重组表达质粒pET23a(+)的大肠杆菌单菌落于5mL LB(Amp+)培养液中,37℃振荡培养过夜后,测量600nm下吸光光度值,收集4 OD/mL细胞,转移至100mL LB(Amp+)培养液中32℃振荡培养,待吸光光度值达0.6 OD/mL (2~3h)加入IPTG,42℃诱导6h,4℃下3000r/min离心30min收获菌体。同时收取少量表达产物进行全蛋白12% SDS-PAGE分析。离心收集沉淀加入5mL BugBuster和5μL Nucleas悬浮,室温下缓慢摇匀10min,避免气泡。4℃下15000r/min离心30min收集上清。上清经His-band纯化试剂盒进行洗脱。将重组液超滤浓缩至2~3mL,-80℃保存。采用12% SDS-PAGE凝胶电泳分析重组蛋白的纯化过程。取晶状体组织匀浆液,分别与还原型GSH(终浓度为5、10mmol/L)或纯化的重组人晶状体TTase(recombinant human lens TTase,RHLT)0.1U在30℃水浴反应30min。反应后加入5×SDS加样缓冲液,于100℃煮沸3min后进行Western blot检测(方法同前)。
表1 两种基因型小鼠晶状体GSH含量
2 结果
2.1小鼠基因型鉴定及生长情况不同基因型小鼠基因鉴定的PCR电泳结果见图2,TTase基因敲除型小鼠扩增出第一对引物(Neo),野生型小鼠扩增出第二对引物(TTase)。TTase基因敲除型小鼠和野生型小鼠生长状况良好,目前为止最长观察1mo发现,两种基因型小鼠在体质量、习性及生长发育方面均非常相似。
2.2不同基因型小鼠白内障发生情况在TTase基因敲除型和野生型小鼠中,白内障的发生均随着年龄增加而增加,混浊类型主要为核性,见图3。在TTase基因敲除型小鼠中,晶状体混浊最早从4月龄发生,可见晶状体核点状混浊,9月龄晶状体核混浊明显加重;而野生型小鼠晶状体混浊最早从9月龄发生,12月龄明显加重。16月龄时两种基因型小鼠晶状体均发展为成熟期白内障。
4月龄TTase基因敲除型小鼠白内障发病率为26%(10/38眼),野生型小鼠晶状体均透明(0/22眼);9月龄TTase基因敲除型小鼠白内障发病率增加至78%(22/28眼),野生型为40%(8/20眼);12月龄TTase基因敲除型小鼠晶状体均发生混浊(18/18眼),野生型发病率为50%(16/32眼);16月龄两种基因型小鼠晶状体均出现明显混浊(TTase基因敲除型20/20眼;野生型12/12眼)。
2.3不同基因型小鼠晶状体GSH含量变化观察不同年龄(1、4、9、16月龄)TTase基因敲除型和野生型小鼠晶状体中GSH含量发现,两种基因型小鼠晶状体GSH含量均随着年龄增加而降低,且9月龄TTase基因敲除型小鼠晶状体中GSH含量明显低于野生型,差异有统计学意义(P<0.05),见表1,图4。
2.4不同基因型小鼠晶状体PSSG蛋白质的表达观察不同年龄(1、4、9、16月龄)TTase基因敲除型和野生型小鼠晶状体PSSG蛋白质的表达发现,两种基因型小鼠晶状体中PSSG蛋白质的表达均随着年龄增加而升高,且形成的PSSG蛋白质主要集中在分子量37~50kDa和100kDa以上。比较两种基因型小鼠晶状体PSSG蛋白质表达灰度值发现,4月龄(P<0.05)和9月龄(P<0.01)TTase基因敲除型小鼠晶状体中高分子PSSG(100kDa以上)蛋白质表达明显高于同年龄野生型小鼠(图5)。
2.5免疫共沉淀法鉴定形成PSSG的蛋白质Western blot可见PSSG条带主要集中在分子量37~50kDa,与骨架蛋白Beta-actin(42kDa)和GAPDH(37kDa)分子量相似,因此利用免疫共沉淀法检测形成PSSG的蛋白质。取组织裂解液分别用抗Beta-actin和抗GAPDH进行免疫沉淀反应,免疫沉淀物经SDS-PAGE凝胶电泳后转膜,并用抗Beta-actin和抗GAPDH抗体进行Western blot。结果发现,在免疫沉淀产物中可以检测到Beta-actin和GAPDH,证实形成氧化产物PSSG中的蛋白质包含Beta-actin和GAPDH(图6)。

图3裂隙灯观察不同年龄两种基因型小鼠晶状体混浊情况1月龄TTase基因敲除型和野生型小鼠晶状体均透明;4月龄基因敲除型小鼠晶状体开始发生核点状混浊,而野生型小鼠晶状体仍然保持透明;9月龄基因敲除型小鼠可见核性混浊及空泡和小水裂,而野生型小鼠晶状体仅可见轻度的混浊;12~16月龄两种基因型小鼠晶状体混浊均加重,逐渐发展为成熟期白内障,包括皮质和后囊下混浊。红色箭头示小鼠晶状体核混浊部位。

图4不同年龄两种基因型小鼠晶状体GSH含量WT:野生型;TTase KO:TTase基因敲除型。aP<0.05vs野生型。

图5Western blot分析不同基因型小鼠晶状体PSSG蛋白质表达A:Western blot检测结果;B:不同基因型小鼠晶状体PSSG 37~50kDa蛋白条带光密度扫描统计图;C:不同基因型小鼠晶状体PSSG 100kDa以上蛋白条带光密度扫描统计图。WT:野生型;TTase KO:TTase基因敲除型。aP<0.05,bP<0.01vs野生型。
图6免疫共沉淀法鉴定形成PSSG的蛋白质A:免疫共沉淀鉴定Beta-actin;B:免疫共沉淀鉴定GAPDH。Pellet:免疫沉淀物;Supernatant:上清;WT:野生型;TTase KO:TTase基因敲除型。
2.6重组人晶状体TTase的脱硫醇作用取9月龄TTase基因敲除型小鼠晶状体裂解液,分别与5、10mmol/L GSH或同时加入纯化的RHLT 0.1U反应,形成的PSSG可被还原剂GSH还原,而加入纯化的TTase后,更多硫醇化的PSSG可被还原,证明TTase具有脱硫醇作用(图7)。
3 讨论
晶状体的氧化损伤是年龄相关性白内障形成的重要因素[1,9]。晶状体中有多重抗氧化屏障,其中包括2种主要蛋白质/酶修复系统:(1)GSH依赖的TTase/Grx系统,通过切断晶状体蛋白质和GSH硫醇化形成的二硫化物(PSSG)维持晶状体的还原状态[10];(2)NADPH依赖的硫氧还蛋白(thioredoxin,Trx)系统,可以有效还原蛋白质和蛋白质之间形成的二硫键(PSSP)[11-12]。这些抗氧化系统可以独立或联合发挥作用,保持晶状体的氧化还原平衡,从而防止白内障的形成。本研究通过建立TTase基因敲除小鼠模型,观察其白内障的发生情况,进一步探讨TTase在白内障发病中的保护机制。
图7Western blot分析TTase的脱硫醇作用control:TTase基因敲除小鼠晶状体裂解液,未加入 GSH或纯化的TTase。
小鼠TTase基因序列包括3个外显子,其中GSH结合氨基酸位点编码于第2外显子,本研究通过敲除第2外显子建立TTase基因敲除小鼠模型。Western blot检测结果显示,TTase基因敲除型小鼠晶状体上皮细胞及其他组织如脑、心、肾均无TTase表达[7]。目前观察TTase基因敲除型小鼠在体质量、习性及生长发育方面与野生型小鼠都非常相似。两种基因型小鼠白内障的发生均呈年龄相关性,且混浊部位表现为核性混浊。TTase基因敲除型小鼠中白内障最早从4月龄开始发生,9月龄明显加重;而野生型小鼠最早从9月龄开始发生,12月龄时明显加重。至16月龄时,两种基因型小鼠晶状体均发生明显混浊。结果提示TTase基因敲除可以加快年龄相关性白内障的发生。
GSH 是晶状体中重要的还原物,其可以通过一系列代谢反应和抗坏血酸不断产生[13]。GSH的损失是发生老化和白内障形成的重要标志之一[1]。比较两种基因型小鼠晶状体GSH含量发现,GSH均随年龄增加而降低,且9月龄TTase基因敲除型小鼠晶状体中GSH含量明显低于野生型,分析与两种基因型小鼠白内障的发生相关,9月龄TTase基因敲除型小鼠白内障的发生率明显高于同年龄野生型小鼠。结果提示TTase基因敲除可以加速GSH的损失。
本课题组前期进行细胞和动物实验研究发现,TTase等抗氧化酶在紫外线(UV)照射后表达上调,说明TTase在抵抗晶状体蛋白质免受紫外线等氧化损伤方面发挥重要作用[14-16]。研究发现,在年龄相关性白内障(包括核性、皮质性、混合性)患者的晶状体中,TTase的活性明显下降[17]。TTase的重要功能之一是通过脱硫醇作用来还原氧化损伤后的蛋白质或酶形成的PSSG,从而修复其功能或活性。PSSG的形成可以使蛋白质结构发生改变,且晶状体中PSSG的积聚可以引起蛋白质和蛋白质之间二硫键的形成和交联,最终导致白内障的发生[18]。在本研究中两组基因型小鼠晶状体PSSG均随年龄增加而增加,且9月龄TTase基因敲除型小鼠晶状体中PSSG表达明显高于同年龄野生型小鼠,主要为高分子聚合物,分析与白内障的发生情况相关。
本研究发现,硫醇化形成PSSG的蛋白质之一为肌动蛋白(actin),其是一条由多肽链构成的分子量为42kDa的球型分子,与肌球蛋白结合构成微丝后有多种生物学功能,如构成细胞骨架、维持细胞形状;参与细胞运动、分泌等活动;参与细胞内信号的传递;参与蛋白质的合成等。大量研究表明,肌动蛋白因含有大量巯基而易被氧化损伤[19]。另一种被硫醇化的蛋白质为GAPDH(37kDa),它在糖酵解途径中对氧化损伤最敏感。GAPDH有4个亚基,每个亚基都包括4个巯基(-SH),其中2个巯基位于活性中心(Cys-149),而活性中心的氧化会导致其酶活性的丧失[20]。由于晶状体属于无氧代谢,这种代谢在晶状体中尤为重要。Xing等[21]发现晶状体上皮细胞与低浓度H2O2孵育后,GAPDH可以和GSH被硫醇化生成PSSG。抵抗氧化损伤引起的细胞毒性及发生永久性修饰,这种硫醇化反应也对GAPDH及其他酶起到保护作用[22]。本研究表明,形成硫醇化的肌动蛋白及GAPDH可被还原剂GSH还原。而加入纯化的TTase后,更多硫醇化的PSSG被还原。证实了这些条带是形成二硫化物的蛋白质,证明TTase通过断裂氧化形成的二硫键,使晶状体中硫醇化的蛋白质脱硫醇,阻止它们交联失活,从而使晶状体蛋白以及膜蛋白保持还原状态,防止白内障的产生。
综上所述,本研究通过建立TTase基因敲除小鼠模型,发现TTase基因敲除可以加速小鼠白内障的发生。晶状体中PSSG的聚积随年龄的增加而增加,且与晶状体的混浊程度相关。这种形成二硫化物的蛋白质主要是肌动蛋白和GAPDH,且可被GSH联合RHLT还原,证实了TTase的脱硫醇作用。
猜你喜欢
中国实用医药(2021年7期)2021-04-16 11:21:38
科学生活(2019年7期)2020-01-01 08:28:02
中医眼耳鼻喉杂志(2019年2期)2019-04-13 05:23:52
养殖与饲料(2019年10期)2019-02-25 14:52:37
石油石化绿色低碳(2019年6期)2019-01-14 01:16:20
山东畜牧兽医(2018年3期)2018-04-26 09:10:34
中国塑料(2016年8期)2016-06-27 06:35:02
当代化工研究(2016年7期)2016-03-20 16:22:03
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
中外医疗(2015年5期)2015-08-29 01:54:42
