
克氏原螯虾生长的损伤性补偿效应
2020-03-08 12:53徐韵苓陈友明王山黄成黄亚红
江苏农业科学 2020年21期
徐韵苓 陈友明 王山 黄成 黄亚红

摘要:为研究断螯对克氏原螯虾生长的影响,以亚成体克氏原螯虾为试验材料,研究了克氏原螯虾失去大螯后的个体存活率与大螯再生、体质量、摄食量和饵料转化效率等的生长状态变化。结果表明,失去大螯后的亚成体螯虾在单养条件下存活率与全肢螯虾无显著性差异(P>0.05),但在非单养条件下存活率显著大于全肢螯虾(P<0.05);螯虾在断螯后有近80%的个体可实现大螯再生,且摄食量、体质量增长率极显著大于全肢螯虾(P<0.01);雌性断螯螯虾饵料转化效率显著大于全肢螯虾(P<0.05)。综上所述,克氏原螯虾断螯后摄食量较多、生长较快,且雌性断螯螯虾饵料利用效率比全肢螯虾高,同时断螯的发生可以有效降低螯虾打斗的死亡率。本研究从生理学和行为学角度对螯虾断螯后的损伤性补偿生长进行了讨论分析,以期对克氏原螯虾的基础理论研究和生产实践提供参考。
关键词:克氏原螯虾;断螯再生;补偿效应;摄食量;体质量增长
中图分类号:S966.12 文献标志码:A 文章编号:1002-1302(2020)21-0207-04
克氏原螯虾(Procambarus clarkii)属甲壳纲、十足目、螯虾科,因其具有个体大、生长快、肉质鲜美和营养丰富等特点[1],已成为养殖户和消费者极其青睐的特色水产品种。但是克氏原螯虾性情凶猛、生性好斗,且种内斗争较为常见[2],在养殖过程中经常出现因相互打斗导致大螯缺失甚至个体死亡等现象。本研究发现一个有趣的情况,在大多数的养殖生产中,无论是选择幼虾还是选择繁育后代的亲虾,虾农普遍认为肢体完整的个体更好;在理论研究中,无论是关于克氏原螯虾繁殖行为的研究,比如张丽等研究的婚配后雄性螯虾的择偶行为,或是关于社会环境与行为方面的研究[3],如王陈路等研究的不同地位螯虾对社会环境改变的行为反应,又或是关于螯虾的食性研究[4],如陈翔宇等研究的食性对螯虾格斗行为的影響[5],在其选择实验动物时都着重强调选用的是螯肢完整的个体,认为断螯个体是不健康的或处于弊端的劣势个体。但在实际生产中有人认为螯肢完整与否并无影响,也有人认为螯肢残缺个体反而更好,那么克氏原螯虾断螯后,个体生长实际情况到底如何呢?
国内外关于生物体断肢现象的研究,如在节肢动物门中蛛形纲的蜘蛛类和甲壳纲的蟹类的断肢研究中发现,断肢后机体发育会延长[6];招潮蟹(Uca arcuata)能通过延长下一次的蜕壳期而使断肢在蜕壳时重新长出[7];谭梁飞等发现,中华真地鳖(Eupolyphaga sinensis)断肢会延长蜕皮间期时间,且断肢数量越多、断肢期日龄越大,间期时间越长,说明断肢会导致生长周期变长,从而保证足够的时间促进机体修复[8];断肢也会延缓甲壳类蜕壳,以便促进断肢恢复,如在甲壳纲十足目的种类中就发现断肢会延长蜕壳间期,使断肢再生后再进行蜕皮[9];锯缘青蟹(Scylla paramamosain)、陆地蟹等在发生肢体缺失现象后,其恢复会受到温度、光周期和动物发育状况等因素影响[10-11]。
目前,关于克氏原螯虾断螯后机体变化等的具体研究未见报道,本研究以亚成体克氏原螯虾为试验材料,通过去螯操作,研究断螯对螯虾生长状况等的影响,不仅可以丰富克氏原螯虾基础生物学研究,同时对克氏原螯虾养殖业也有一定的指导作用。
1 材料与方法
1.1 材料
本试验于2018年5—9月在江苏省淡水水产研究所扬中基地进行。试验用虾来源于江苏省淡水水产研究所扬中基地,试验前将虾置于基地内水泥池暂养3 d,待虾生长稳定后再进行试验。试验时,从同一批螯虾中选出一批无病无伤、附肢完整、活力强的亚成体螯虾,平均体质量为(3.41±0.68) g,根据试验需要分别置于规格为48 cm×27 cm×28 cm 的长方体塑料箱和上口直径 17.5 cm、下底直径13.0 cm、高10.0 cm的圆台形塑料容器中进行饲养观察,饲养期间投喂嘉吉牌河蟹饲料(SC/T 1078—2004)。
1.2 方法
1.2.1 试验设计 试验分为2个部分,第一部分为单养试验,每个碗状容器饲养1尾克氏原螯虾。将试验用虾进行编号、称量体质量,随机分为2组:试验组、对照组各60尾(雌雄各半)。第二部分为非单养试验,每个长方体容器放置2尾性别相同的螯虾。将试验用虾进行编号、称量体质量,随机分为2组:试验组、对照组各40尾(雌雄各半)。
试验组克氏原螯虾的大螯用人工压力法去掉大螯后再次称质量记录体质量。所有螯虾分别放置在各组相应的实验容器中饲养。每天定时投喂饲料并在2 d投喂前记录死亡数、摄食量后,换掉 1/2 的水,清理残余饲料同时观察残肢的生长恢复情况。试验期间保持各个容器的养殖环境基本一致。单养试验进行45 d,非单养条件试验进行30 d。
1.2.2 指标测定与计算 体质量增长率、饵料转化效率和死亡率计算公式如下:
(3)死亡率的百分比检验公式为:
式中:m0、mt分别为试验开始时和结束时的螯虾体质量,t为试验天数(d);F为摄食总量;a为试验组死亡数;A为试验组样本容量;b为对照组死亡数;B为对照组样本容量。百分比检验结果,u>1.96,P<0.05(表示差异显著);u>2.58,P<0.01(表示差异极显著)。
1.3 数据处理
试验结果采用“平均数±标准差(x±s)”表示,利用Excel 2013、SPSS 25.0软件进行独立样本t检验分析,显著性水平设为0.05。另外还采用百分比检验分析。
2 结果与分析
2.1 单养分析
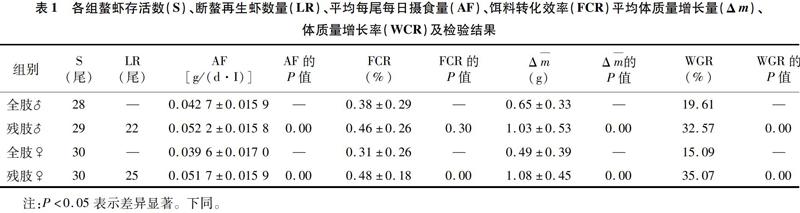
螯虾存活尾数(S,Survival)、断螯再生尾数(LR,Limb Regeneration)、平均摄食量[AF,Average Food Intake(g/(d·I))]、饵料转化效率(FCR,Food Conversion Ratio)和平均体质量变化(Δm)、体质量增长率[WGR,Weight Growth Rate(%)]等情况及检验结果见表1。
2.1.1 存活与再生 由表1可知,试验过程中雄性全肢螯虾存活28尾,雄性残肢螯虾存活29尾;雌性全肢及残肢螯虾均无死亡,这表明断螯对螯虾死亡率并无影响。亚成体残肢螯虾最终存活59尾,螯肢再生的共有47尾,占存活数量的比例为79.67%。
2.1.2 摄食量分析 由表1可知,残肢螯虾摄食量大于全肢螯虾。比较试验组和对照组间的平均摄食量情况,双侧检验结果表明,亚成体克氏原螯虾残肢螯虾摄食量极显著大于全肢螯虾(P<0.01),说明亚成体螯虾在断螯后摄食量较全肢螯虾多。
2.1.3 饵料转化效率 由表1可知,残肢螯虾饵料转化效率高。双侧检验结果表明,雄性残肢螯虾饵料转化效率与雄性全肢无显著差异(P=0.30>0.05),雌性残肢螯虾饵料转化效率显著高于雌性全肢螯虾(P=0.00<0.05)。综上所述,断螯会提高雌性克氏原螯虾的饵料转化效率。
2.1.4 体质量增长量、体质量增长率 由表1可知,残肢螯虾的平均体质量增长量和体质量增长率皆比全肢螯虾大。比较试验组和对照组间体质量情况,双侧检验结果表明,残肢螯虾体质量增长量极显著大于对照组全肢螯虾(P<0.01)。考虑到全肢螯虾最终体质量包含大螯质量而残肢螯虾没有,所以以体质量增长率为标准衡量体质量变化情况更为合理,双侧检验结果表明,残肢螯虾体质量增长率极显著大于对照组全肢螯虾(P<0.01),表明断螯促进了克氏原螯虾体质量的增加,试验组残肢螯虾出现了超补偿生长。
2.2 非单养条件下存活试验
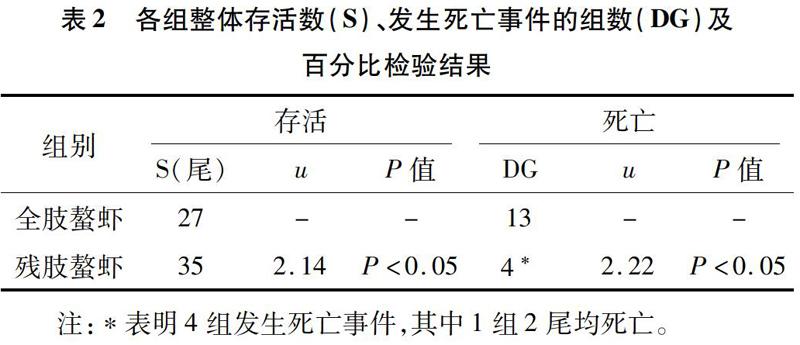
在非单养条件下,为探究断螯对克氏原螯虾的影响,采取2尾螯虾共养模式,观察残肢螯虾、全肢螯虾在社会社区压力和断螯损伤双重压力的应激状态下的存活率。试验以发生死亡的组数和死亡个体数量作为衡量指标,通过百分比检验比较显著性差异。因为螯虾生性好斗,在非单养条件下存在同类相残相食现象,所以无法准确测定螯虾的摄食量,非单养试验将不再探究螯虾的摄食量和体质量变化情况。试验各组存活螯虾尾数(S)、发生死亡组数(DG,Died Group)及相应百分比检验结果见表2。
2.2.1 死亡率统计与检验 由表2可知,试验组螯虾有4组出现死亡事件,对照组有13组出现死亡事件,对其进行百分比检验,u=2.22>1.96,P<0.05,即有显著性差异,说明试验组发生死亡事件的概率显著小于对照组。且死亡13尾克氏原螯虾中有6尾打斗致残,说明是经过激烈打斗致死的,统计个体死亡情况,对非单养条件下试验的死亡率数据进行百分比检验,残肢螯虾与全肢螯虾死亡率百分比检验结果为u=2.14>1.96,P<0.05,即有显著性差异,说明非单养条件下残肢螯虾比正常螯虾死亡率要低。
由表2可知,非单养条件下残肢螯虾及全肢螯虾死亡率分别与表1中单养条件下残肢螯虾及全肢螯虾死亡率进行百分比检验,结果分别为u=2.21>1.96(P<0.05)和u=3.98>2.58(P<0.01)(表2未显示该数据)。这说明只是断螯压力不会对螯虾存活产生应激刺激,生存不会受到影响(单养条件下全肢与残肢螯虾死亡率无显著性差异);但非单养条件下在社会社区压力和断螯双重压力应激下,特别是社会竞争压力的增大显著,甚至极显著(全肢螯虾死亡率对比检验)提高了螯虾的死亡率。
3 讨论与展望
3.1 存活与再生
在单养试验中,残肢螯虾与全肢螯虾死亡率无显著差异,这表明螯虾并不会因断螯而增加死亡率。在本研究和已有文献对克氏原螯虾探讨中均发现,克氏原螯虾存在自切现象[2],即螯虾的大螯在外力胁迫下会从关节处自我切断,这可能是螯虾为防止血液外流、扩大伤口感染,避免因断螯而导致死亡,这种现象是大多数节肢动物在漫长进化过程中形成的自我保护机制[11]。而在非单养条件下,残肢组死亡率显著低于全肢组,这可能是因为克氏原螯虾性格凶猛,生性好斗[2],断螯有效避免了螯虾因为利用大螯相互打斗而导致死亡。非单养条件下螯虾死亡率显著大于单养条件,表明只有断螯不会对螯虾存活产生应激刺激,其生存不會受影响(单养条件下全肢与残肢螯虾死亡率无显著性差异),但在非单养条件下社会社区压力和断螯双重压力应激特别是社会竞争压力的增大,显著甚至极显著地提高了螯虾死亡率。但在社会压力下,断螯的螯虾存活率又显著高于全肢螯虾,说明断螯提高了螯虾社会竞争力,有效降低了螯虾因为相互打斗而导致的死亡率,断螯可有效提高存活率。
本研究中有近80%残肢螯虾重新长出大螯,但在中华绒螯蟹(Eriocheir sinensis)断肢研究中残肢蟹全部长出完整肢[12],作者猜测可能是因为中华绒螯蟹和克氏原螯虾断肢再生恢复机制有所不同,也可能是本次试验周期不够长,不足以让全部螯虾完成断螯再生。
3.2 断螯对摄食影响
本试验中克氏原螯虾虽然缺失了大螯,在一定程度上减少了运动的消耗量,但是通过螯虾摄食量数据分析发现,大螯的缺失反而提高了残肢螯虾的摄食量。生物界不同生物在断肢后会有不同的摄食策略来应对大螯损失的能量代价,如对多数昆虫的研究中特别是普列热蝽,在肢体发生残缺后会大量进食,进入蜕皮期通过蜕皮来促进断肢再生[13],本试验结果与之类似。但在尾鳍缺失对3种鲤科鱼类影响的研究中结果相反[14],3种鲤科鱼类在尾鳍缺失后体内摄食代谢模式会减少,而把更多的能量分配到运动上,补偿因为尾鳍缺失而造成的运动能力减弱这个现象。考虑到大螯是克氏原螯虾主要的打斗器官,同时大螯的缺失也不会影响克氏原螯虾的运动能力,所以在发生断螯现象后,运动不会更多消耗体内能量。有的动物断肢后摄食量不变,如丽斑麻蜥断尾既不促进也不抑制该种蜥蜴摄食量[15],该试验研究者认为在该种蜥蜴中补偿断尾能量代价对策不是增加摄食,猜测可能是以减少活动来节约体内能量损耗。但考虑到丽斑麻蜥属于爬行纲,它们的断肢再生是属于直接生长出来,而不是像甲壳纲的虾、蟹等,昆虫纲的蜘蛛等以蜕皮促进生长,恢复断肢。因此,本研究认为克氏原螯虾在断肢后恢复断肢的对策基本上是通过增加摄食来实现补偿生长。
3.3 体质量增长及饵料转化效率
生命有机体的损伤性修复是一种生长过程。克氏原螯虾在受到断螯胁迫后,以大量摄食对策来弥补损失大螯的能量代价实现快速生长,这种机制与饥饿性损伤补偿生长类似。补偿生长分为超补偿生长、完全补偿生长、部分补偿生长和不补偿生长4种类型[16]。在克氏原螯虾对多重周期饥饿研究中发现,克氏原螯虾可以在胁迫环境下产生补偿生长[17]。克氏原螯虾在经历断螯胁迫后迅速适应,体质量增加、平均体质量增长量和体质量增长率较全肢螯虾呈极显著差异,残肢螯虾表现出超补偿生长,这与对海星研究的结果类似,残肢海星会极显著增加体质量,以尽快恢复觅食能力,同时恢复攻击的武器[18]。断螯刺激克氏原螯虾大量积累能量,外在表现就是体质量增加,然后将能量分配至损失的断螯处,从而恢复断螯,对狼蛛研究中也发现类似结果[19],同时狼蛛会减少其他方面能量消耗,集聚体内能量到下一个蜕皮期,以减轻体质量和减小体型为代价来恢复断肢。
在本研究中雌性残肢克氏原螯虾的饵料转化效率显著大于全肢螯虾组,说明雌性残肢螯虾对饲料的利用率和转化率提高了。在摄食量相同情况下,雌性断螯螯虾的体质量增加量较多。笔者猜测可能是亚成体雌性螯虾为了即将到来的生殖活动,所以要更加高效地储存能量同时还要促进性腺的进一步发育。当生物体遭遇外力胁迫特别是不可抗的强大因素时都会选择在最后将体内能量更多分配到生殖活动上,从而繁衍后代。如淡水蟹在断肢刺激后会促进卵巢的发育[20],说明在遭遇外界不良环境时会将更多的能量倾向于生殖活动上。本试验中则体现在雌性螯虾断螯后的体质量超补偿性增加。
3.4 展望
亚成体克氏原螯虾对断螯产生的机体应激机制还不完全清楚,下一步还可以探究体内哪些激素分泌和相关基因表达以及微小核酸作用于摄食量、体质量等,从而揭示其内部变化。成体克氏原螯虾的生长虽然已经定型,但是在受到断螯损伤胁迫后机体是否会产生相应应激状态还未可知,也可探究成年螯虾在断螯后的生理生化状态。
参考文献:
[1]何金星,窦 寅,唐建清,等. 饲料添加螺旋藻对克氏原螯虾生长性能的影响[J]. 江苏农业科学,2009(2):209-211,250.
[2]王 庆. 克氏原螯虾繁育机制及养殖生态学研究[D]. 南京:南京师范大学,2012:2-44.
[3]张 丽,谢 春,唐建清,等. 婚配后雄性克氏原螯虾的择偶行为[J]. 南京大学学报(自然科学版),2014,50(6):890-894.
[4]王陈路,曾小翠,沈佳琰,等. 不同地位克氏原螯虾对社会环境改变的行为反应[J]. 动物学杂志,2015,50(4):555-562.
[5]陈翔宇,王陈路,黄 成,等. 食性对克氏原螯虾格斗行为的影响[J]. 饲料工业,2014(增刊1):117-120.
[6]González M,Barría E. Effect of autotomy and regeneration of the chelipeds on growth and development in Petrolisthes laevigatus (Guérin,1835) (Decapoda,Anomura,Porcellanidae)[J]. Crustaceana,2008,81(6):641-652.
[7]Holland C A,Skinner D M. Interactions between molting and regeneration in the land crab[J]. The Biological Bulletin,1976,150(2):222-240.
[8]譚梁飞,朱 芬,熊 强,等. 中华真地鳖断足再生对发育的影响[J]. 昆虫知识,2007,44(1):101-104.
[9]Donald L M. Interactions between limb regeneration and molting in decapod crustaceans[J]. American Zoologist,2001,41(3):399-406.
[10]Jie Gong,Yu Kun,Ling Shu,et al. Evaluating the effects of temperature,salinity,starvation and autotomy on molting success,molting interval and expression of ecdysone receptor in early juvenile mud crabs,Scylla paramamosain[J]. Journal of Experimental Marine Biology and Ecology,2015,464:11-17.
[11]Mattoni C I,García-Hernández S,Botero-Trujillo R,et al. Scorpion sheds ‘tail to escape:Consequences and implications of autotomy in scorpions (Buthidae:Ananteris)[J]. PLoS One,2015,10(1):e0116639.
[12]岳武成,陈 娇,慈元吉,等. 断肢再生对中华绒螯蟹蜕壳、生长及相关基因表达的影响[J]. 浙江大学学报(农业与生命科学版),2016,42(4):502-508.
[13]王孟卿,彩万志. 昆虫肢体再生的研究进展[J]. 昆虫知识,2004,41(2):127-131.
[14]付 成,曹振东,付世建. 尾鳍缺失和再生对三种鲤科鱼类游泳运动能力和摄食与运动代谢交互作用的影响[C]//中国海洋湖沼学会鱼类学分会、中国动物学会鱼类学分会2012年学术研讨会.兰州,2012.
