
碳酸盐鲕粒包壳结构研究综述*
2020-02-27 12:32宋文天刘建波
古地理学报 2020年1期
宋文天 刘建波,2
1北京大学地球与空间科学学院,北京 100871 2造山带与地壳演化教育部重点实验室(北京大学),北京 100871
1 概述
碳酸盐鲕粒是广泛分布于现代和古代碳酸盐沉积物中的颗粒,因其独特的结构特征及其在古海洋、古环境恢复方面的广泛应用而备受关注(Sandberg,1983;Strasser,1986;Opdyke and Wilkinson,1990;Lietal., 2015;Diaz and Eberli, 2019)。鲕粒包壳结构在现代沉积和古代地层中存在多种类型,原始矿物组成也存在差异,保存程度不尽一致,被广泛应用于古环境和海洋化学性质的研究(Loreau and Purser,1973;Landetal., 1979;Sandberg,1983;Strasser,1986;Opdyke and Wilkinson,1990)以及成岩过程分析(Wilkinson and Landing,1978;Wilkinsonetal., 1985;Chow and James,1987;Hird and Tucker,1988;Algeo and Watson,1995)。
中国碳酸盐鲕粒研究材料丰富,在中元古界至古近系中广泛分布(如余素玉等,1987;柳永清等,1999;吴熙纯,2009;齐永安等,2014)。国内鲕粒包壳结构的研究晚于国外,大致始于20世纪80年代(王英华等,1983;张秀莲,1984),研究内容主要集中于古代鲕粒方面,现代鲕粒和鲕粒实验模拟(如赵震等,1984;周瑶琪等,2017)等方面的研究相对薄弱。基于中国的研究材料,国内诸多学者识别出了古代鲕粒包壳的原生结构和次生结构,并逐渐接受国外主流观点(王英华等,1983;张秀莲,1984;韦龙明,1995;沙庆安和江茂生,1998;金瞰昆,1998)。自2000年以来,古代鲕粒的包壳结构多被应用于沉积微相和古环境分析(如刘伟和张兴亮,2010;侯恩刚等,2014;Tianetal., 2015;姬国锋等,2016;Huangetal., 2017),且巨鲕的包壳结构研究也逐渐受到重视(如Lietal., 2013,2015;Tangetal., 2015;Tanetal., 2018)。
现代鲕粒包壳具有多种原生结构类型,并具有不同的矿物成分、生长模式和形成环境。古代鲕粒的包壳虽然经历了不同程度的成岩作用,但常能够保存可识别的原生结构(Sandberg,1983;Tucker,1984;Wilkinsonetal., 1985;Algeo and Watson,1995)。对鲕粒包壳结构的准确识别和分类,是根据鲕粒特征开展古气候、古环境、古海洋研究的关键。目前,鲕粒包壳结构的研究尚存在一些问题: 对现代鲕粒包壳原生结构的认识尚未形成统一体系,一些术语定义不明确;对于现代鲕粒结构术语的理解存在偏差,导致对古代鲕粒包壳原生结构的特征识别和分类不尽准确。
文中通过综述前人对现代鲕粒的研究成果,归纳了各类包壳原生结构的主要特征,并明确提出在对包壳进行分类时应当对包壳的结构组合进行识别。此外,还总结了古代鲕粒的各类包壳结构,分析了中国鲕粒研究中存在的问题,并以华南下奥陶统鲕粒为例,对常见于古代鲕粒包壳中的原生结构进行了识别。
2 鲕粒包壳结构的研究历史
国际上,鲕粒包壳结构及其成因的研究历史大致可划分为3个阶段。
2.1 早期研究阶段(1855-1960s)
Sorby(1879)最早对鲕粒进行了深入观察,他在现代鲕粒中识别出了同心状结构(concentric structure),在古代鲕粒中识别出了放射状结构(radial structure)。之后的学者也从现代和古代鲕粒中分别识别出这2类包壳结构(如现代巴哈马滩,Illing,1954;Newelletal., 1960)。该时期,大多数学者认为同心状包壳通常为文石质,而放射状包壳为方解石质,且构成放射状包壳的方解石晶体是在成岩过程中由原始文石晶体转变而来的,包壳结构随着矿物成分的变化从同心状转变为放射状(Eardley,1938;Carrozi,1962),即: 现代鲕粒包壳的同心状结构是鲕粒生长过程中形成的、未经成岩作用改造的原生结构,而现代和古代鲕粒包壳的放射状结构是经成岩作用改造而成的次生结构(Eardley,1938;Carrozi,1962)。
2.2 深入研究阶段(1960s-1990s)
自上世纪60年代开始,对现代鲕粒包壳结构的研究取得了更多进展。大盐湖、得克萨斯巴芬湾、波斯湾、大堡礁等地的现代鲕粒被广泛报道和研究(Rusnack,1960;Halley,1977;Landetal., 1979;Medwedeff and Wilkinson,1983)。Kahle(1974)通过 X 射线衍射方法确定了大盐湖鲕粒包壳中的放射状晶体实为文石,而非次生方解石。与此同时,多项模拟实验人工合成了具有放射状、同心状包壳的鲕粒,进一步证实这2种结构均可形成于鲕粒的原始生长阶段(Donahue,1969;Suess and Fütterer,1972;Daviesetal., 1978;Fergusonetal.,1978)。自此,现代鲕粒的放射状、同心状结构均为包壳原生结构的观点逐渐成为学界主流认识(Rusnack,1960;Freeman,1962;Bathurst,1967;Loreau and Purser,1973)。具有不同类型包壳结构的鲕粒的地理分布表明,水动力条件是包壳结构的重要控制因素(Rusnack,1960;Loreau and Purser,1973;Davies and Martin,1976;Landetal., 1979)。
与此同时,对古代鲕粒包壳原生和次生结构的研究更为深入。Sandberg(1975)通过比较不同矿物在成岩过程中的形态演化,推断古代鲕粒中的放射状包壳结构应为原生结构。之后的研究表明,依据放射状晶体粒度、粗细变化可以推断包壳结构的次生变化过程,根据晶体在不同细层中长度的差异可判断包壳形成过程中磨蚀强度的变化(Tucker,1984;Chow and James,1987;Helleretal., 1980;Bates and Brand,1990)。此外,古代鲕粒中2种常见的包壳结构——晶粒结构(sparry/blocky structure)和泥晶结构(也被称为 micritic structure,但应与原生微晶纹层加以区分)——被归入次生包壳结构,其中晶粒结构可能为溶蚀—充填作用形成的,泥晶结构可能是微生物泥晶化作用的结果(Sandberg,1975;Wilkinson and Landing,1978;Wilkinsonetal., 1985)。根据文石和方解石的稳定性差异,研究者推测保存原生放射状结构包壳的原始矿物成分为稳定性较强的高镁方解石或方解石,溶蚀—充填作用形成的晶粒结构包壳的原始成分可能为稳定性差的文石,并认为这2类包壳在地层中的分布反映了地质历史时期海洋化学性质的演化(Friedman,1964;Sandberg,1983;Majoretal., 1988;Opdyke and Wilkinson,1990;Algeo and Watson,1995)。
2.3 微生物作用研究阶段(2000s至今)

近年来,一些粒径超过2imm、以深色泥晶结构为主的古代巨鲕中的微生物化石或活动遗迹被发现,并被作为鲕粒微生物成因的直接证据(如Trower and Grotzinger, 2010;Liu and Zhang,2012;Lietal., 2013;Lietal., 2017)。需要指出的是,多数巨鲕的包壳是以含有微生物化石的泥晶层为主体的,形成模式与现代鲕粒包壳截然不同(Troweretal., 2010;Lietal., 2013,2017),其本质上是一种微生物颗粒,与核形石相近,与鲕粒不同。此外,有些微生物化石或遗迹并不出现在典型的鲕粒包壳结构中,而是出现在古代鲕粒包壳表面或同心圆状细层之间的泥晶层中。因此,典型鲕粒的微生物成因机制尚有待进一步的研究。
3 现代鲕粒包壳的原生结构特征及其成因
前人对现代鲕粒包壳进行了详细研究,描述了各类包壳结构的结晶学和矿物学特征(Rusnack,1960;Loreau and Purser,1973;Marshall and Davies,1975;Sandberg,1975;Davies and Martin,1976)。然而,现代鲕粒包壳原生结构研究结果多散布在各研究阶段的文献中,部分术语定义不明确,不同学者对术语的理解和用法也有所不同,结构分类方案未形成统一体系。这些问题不但影响到现代鲕粒的包壳结构研究,而且在古代鲕粒的包壳结构研究中亦造成了混淆。因此,在对古代鲕粒包壳结构进行综述前,首先需要对现代鲕粒包壳原生结构研究开展综合分析。
根据组成包壳的碳酸盐晶体的形态、排列方向等特征,现代鲕粒包壳结构可分为放射状和同心状2类,并形成 8种结构组合类型。
3.1 放射状结构
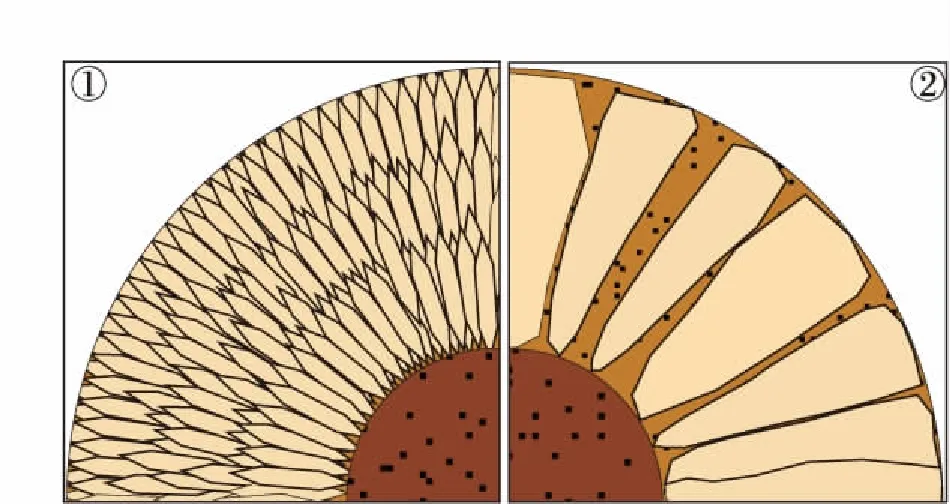
1—放射纤维状结构;2—放射板状结构图 1 现代鲕粒包壳中的放射状结构Fig.1 Radial structures in modern ooids cortices
现代鲕粒包壳的放射状结构是指晶体沿近垂直于核心表面或纹层的方向排列(图 1,表 1)(Rusnack,1960;Freeman,1962;Loreau and Purser,1973;Kahle,1975;Sandberg,1975)。这些晶体多为单向延伸,呈半自形或他形,具体表现为纤维状、针状、片状、棒状、板状等形态(Loreau and Purser,1973;Sandberg,1975;Davies and Martin,1976;Halley,1977;Landetal., 1979)。晶体粒度分布范围较广,在 0.1×1iμm 与 50×150iμm 的区间内连续分布(Loreau and Purser,1973;Halley,1977;Landetal., 1979)。其中,粒径较小的晶体通常呈半自形纤维状、针状(图 1-1)(Loreau and Purser,1973;Landetal., 1979);粒径较大晶体通常为他形棒状、板状,有时存在向鲕粒外侧增粗的现象,末端可呈一个平直的截面(图 1-2)(Sandberg,1975;Halley,1977)。放射状结构由此可分为放射纤维状(图 1-1)和放射板状(图 1-2)2个亚类。根据晶体横断面形态、晶体侧面在酸蚀后显示出类似生长纹的结构,可以推断这些较大晶体可能是由众多次级小晶体构成的集合体(Sandberg,1975;Halley,1977)。晶体长轴有时并不严格垂直于核心表面或纹层,相对垂向可能有 20°~30° 夹角(Sandberg,1975)。
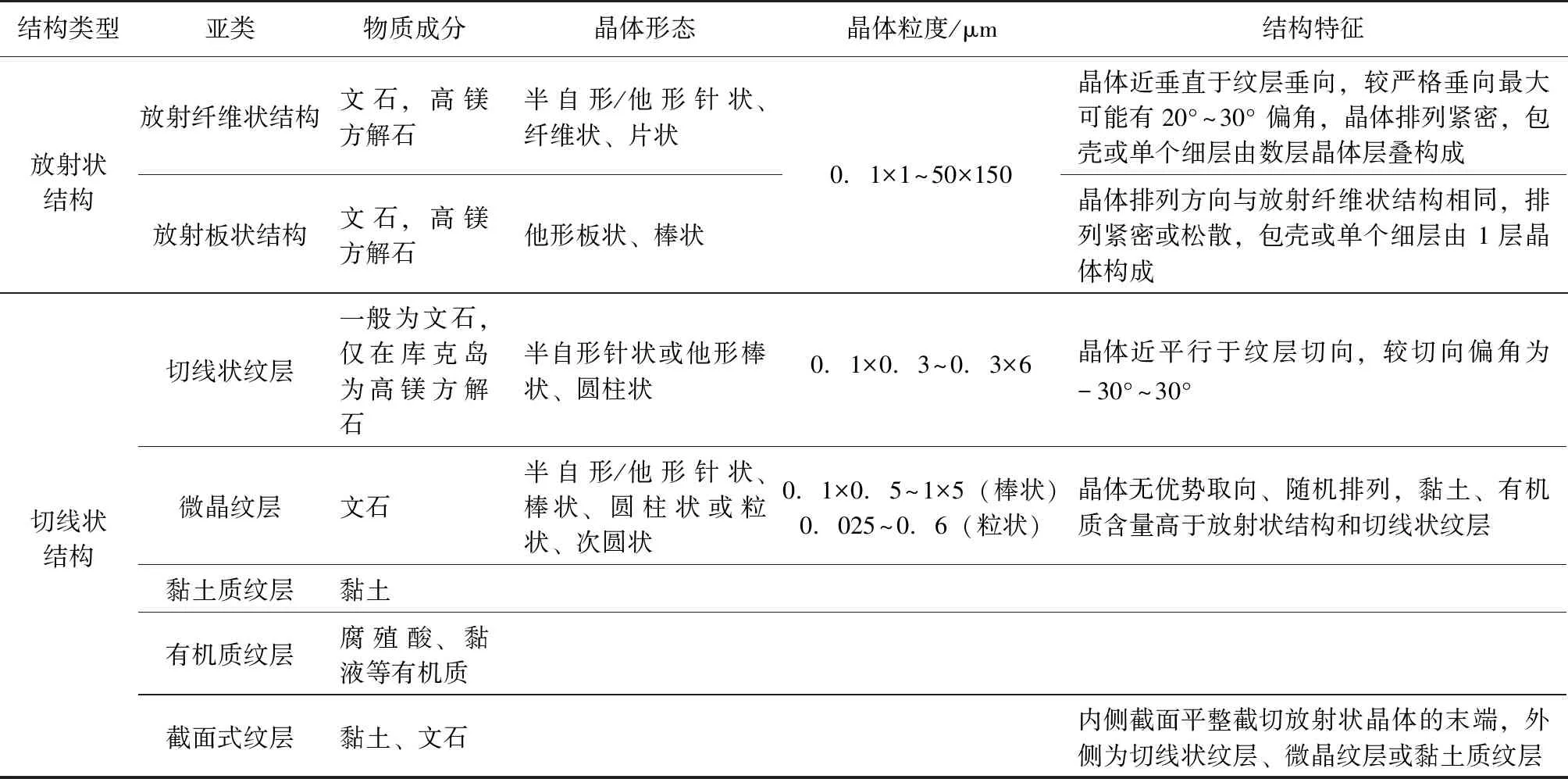
表 1 现代鲕粒包壳结构(据以下文献总结: Newell et al., 1960;Loreau and Purser,1973;Sandberg,1975;Marshall and Davies, 1975;Davies and Martin,1976;Halley,1977;Land et al., 1977;Simone,1981;Kahle,2007;Rankey and Reeder,2009等)Table 1 Cortical structures of modern ooids(Summarized from the following references: Newell et al., 1960;Loreau and Purser, 1973;Sandberg,1975;Marshall and Davies,1975;Davies and Martin,1976;Halley,1977;Land et al., 1977;Simone,1981; Kahle,2007;Rankey and Reeder,2009;etc.)
中、小粒径的放射状晶体一般紧密排列为束状或扇状(图 1-1),而较大粒径的放射状晶体既可能紧密排列,也可能松散排列,晶体间的间隙由文石泥及黏土、有机质等杂质所充填(图 1-2)(Sandberg,1975)。由粒径较小的晶体紧密排列而成的晶体束之间,也可能存在被文石泥、黏土等充填的间隙(Sandberg,1975)。原生的放射状晶体一般不会延伸至其两侧纹层或鲕粒外边界之外(Marshall and Davies,1975),单个放射状细层或放射状包壳可能由1层粒径较大的板状晶体构成,也可能由数层粒径较小的纤维晶体层叠而成(图 1-1,1-2)(Sandberg,1975;Halley,1977)。放射状晶体成分通常为高镁方解石或文石,颜色较浅,晶体间隙中富集细粒充填物处颜色较深(图 1-2)(Sandberg,1975;Halley,1977;Simone,1981)。由于晶体 c 轴方向通常与长轴方向近一致,放射状结构在正交光下呈现出“黑十字”(Sandberg,1975;Marshall and Davies,1975;Halley,1977)。
根据人工合成鲕粒的实验结果,放射状结构应为晶体在颗粒表面自然生长的结果(Suess and Fütterer,1972;Daviesetal., 1978;赵震等,1984)。实验结果和现代放射状鲕粒的分布表明,放射状结构可能形成于相对平静的水体(Rusnack,1960;Loreau and Purser,1973;Daviesetal.,1978;Landetal., 1979)。也有少数学者认为放射状结构的形成环境可能是高盐度水体或淡水环境(Friedmanetal., 1973;Halley,1977),但该假说无法解释在大堡礁正常盐度海水中发现的放射鲕(Davies and Martin,1976)。
部分具有放射状结构的包壳呈现出脑状外形(Carrozi,1962;Kahle,1974)。这种脑状包壳由放射状晶体或晶体束及其侧边之间富含黏土等杂质的隐晶质充填物构成,鲕粒边界处晶体束向外凸出、充填物向内凹陷,使得鲕粒外形起伏不平(Carrozi,1962;Kahle,1974)。脑状鲕的成因可能是放射状晶体增生导致其侧边之间的充填物体积减小、形成内凹形态(Carrozi,1962;Halley,1977),或放射状晶体形成之后,充填物以“U 形”形态在其间隙中沉积(Halley,1977)。少数学者认为充填物的内凹形态可能与微生物的侵蚀作用有关(Kahle,1974)。
3.2 同心状结构
鲕粒中的同心状结构由较薄的同心状纹层构成。不同类型的纹层相间排列在包壳中,根据其结构、成分等差异,纹层可划分为切线状、微晶、有机质、黏土质、截面式 5种类型。
1)切线状纹层(tangential lamina)。纹层由长轴与纹层切向近平行的晶体构成(图 2-1,表 1)(Illing,1954;Rusnack,1960;Duguidetal., 2010;Heartyetal., 2010;Diazetal., 2017)。这些晶体通常为单向延伸,呈终端圆钝的棒状、圆柱状等他形形态或终端尖锐的针状等半自形形态(Loreau and Purser,1973;Halley,1977;Landetal.,1979;Folk and Lynch,2001;Kahle,2007)。晶体长度为0.3~6iμm,直径为 0.1~0.3iμm(Loreau and Purser,1973;Halley,1977;Kahle,2007)。晶体长轴通常并不严格平行于纹层切向,与切向呈-30°~30° 夹角(图 2-1)(Loreau and Purser,1973)。切线状纹层的成分一般为文石(Loreau and Purser,1973;Sandberg,1975;Halley,1977;Simone,1981),高镁方解石质的切线状纹层仅见于库克岛(Rankey and Reeder,2009)。纹层颜色较浅,可能混有微量的黏土、有机质等杂质(Loreau and Purser,1973;Sandberg,1975;Halley,1977)。晶体 c 轴方向往往与长轴方向一致,纹层在正交光下呈现出“黑十字”(Loreau and Purser,1973;Sandberg,1975;Halley,1977;Landetal., 1979)。
对于切线状纹层成因的认识尚有争论。较强的水动力搅动有可能使棒状或圆柱状晶体在鲕粒表面呈切线状排列,由此有些学者认为与放射状结构相比,切线状纹层形成于更强的水动力环境中(Rusnack,1960;Loreau and Purser,1973;Landetal., 1979),且各种包壳类型鲕粒的沉积环境分布特征(Rusnack,1960;Loreau and Purser,1973;Landetal., 1979)、鲕粒的人工合成实验结果(Daviesetal., 1978)都提供了支持证据。不过,也有学者认为切线状纹层的形成可能与晶体矿物成分有关,文石晶体相对易碎,在动荡水体中更容易被改造为切线状结构,而脆性相对较弱的高镁方解石晶体则更易保留放射状结构(Landetal., 1979)。
2)微晶纹层(micritic lamina)。构成纹层的晶体粒度较细,且晶轴无优势取向、随机排列(图 2-2)(Newelletal., 1960;Rusnack,1960;Loreau and Purser,1973;Landetal., 1979;Medwedeff and Wilkinson,1983)。晶体一般为与切线状结构类似的针状或棒状晶体(图 2-2),或粒径 0.025~0.6iμm 的次圆状、粒状晶体(Loreau and Purser,1973;Medwedeff and Wilkinson,1983;Kahle,2007)。微晶的分布往往不会延伸到其所在纹层之外(图 2-2)(Landetal., 1979)。微晶的成分为文石,文石之间的黏土、有机质等杂质含量较切线状晶体、放射状晶体之间更多,且分布更为弥散(Rusnack,1960;Newelletal., 1960;Loreau and Purser,1973)。在部分鲕粒中,微晶层分布不连续,呈不完整的同心圆状(Newelletal., 1960)。纹层整体颜色一般较深(Rusnack,1960)。
原生微晶纹层的成因目前尚无定论,现有的假说包括晶体在较快的碳酸盐沉积速率下难以形成优势取向(Rusnack,1960),或晶体在介于放射状结构、切线状结构之间的扰动强度下难以形成优势取向等(Medwedeff and Wilkinsion,1983)。
3)有机质纹层。通常由腐殖酸或黏液构成,颜色较深(图 2-3)(Shearmanetal., 1970;Marshall and Davies,1975)。
4)黏土质纹层。由黏土矿物构成,颜色较深(图 2-3)(Sandberg,1975)。
5)截面式纹层。纹层内侧为截切放射状晶体的平直截面(图 2-4,2-5,2-6)(Sandberg,1975;Troweretal., 2017)。部分截面附近富集黏土,构成截面—黏土质纹层(图 2-4)(Sandberg,1975)。部分截面外侧发育微晶质纹层,构成截面—微晶质纹层(图 2-5)(Sandberg,1975)。还有部分截面外侧发育切线状纹层,构成截面—切线状纹层(图 2-6)(Sandberg,1975)。
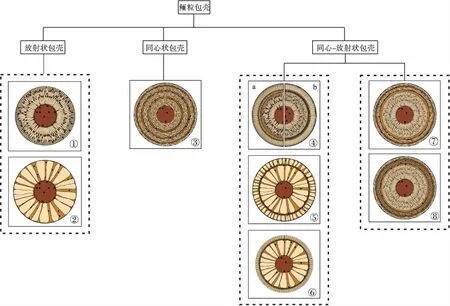
1—放射纤维状包壳;2—放射板状包壳;3—同心状包壳;4—同心—放射纤维状包壳: a—同心状结构为截面式纹层,b—同心状结构为微晶质纹层、有机质纹层或黏土质纹层;5—同心—放射板状包壳层;6—同心—放射板状—放射纤维状包壳;7—同心—放射状包壳,由内部放射纤维状包壳、外部同心状包壳(含切线状纹层)构成;8—同心—放射状包壳,由内部放射纤维状包壳、外部同心状包壳(含切线状纹层、微晶纹层)构成图 3 现代鲕粒包壳的结构组合类型Fig.3 Structure assemblage types of modern ooids cortices
有机质纹层、黏土质纹层、截面式纹层可能形成于鲕粒中碳酸盐晶体生长停滞的时期,现代鲕粒中有机质纹层两侧的14C 年龄相差可达 500~3000 a(Duguidetal., 2010)。沉积速率变化、溶液化学性质变化、磨蚀作用以及阻碍结晶的物质(如一些疏水性低分子物质、Mg2+等)在晶体表面吸积等,均可能导致晶体生长停滞(Sandberg,1975;Daviesetal., 1978;Duguidetal., 2010),在停滞时期鲕粒表面可能加积文石微晶、有机质、黏土等形成纹层(Sandberg,1975)。磨蚀作用可能会使同心状结构中出现平直的截面,并与微晶、黏土等共同构成截面式纹层(Sandberg,1975;Troweretal., 2017)。
3.3 结构组合类型及其特征
现代鲕粒包壳可能并不是由单一的原生结构构成,而是呈现出多种结构的组合,因此鲕粒包壳的分类应当以结构组合类型为依据。根据是否发育放射状或同心状结构,可将包壳结构分为 3大类: 放射状、同心状、同心—放射状(图 3)。各大类包壳可根据放射状、同心状结构的特征进一步细分,如图 3 所示。
放射状包壳呈连续的放射状结构(图 3-1,3-2)(Flügel,2010),可根据放射状晶体的粒度、形态进一步划分为放射纤维状包壳和放射板状包壳。同心状结构由切线状纹层和其他类型的纹层相间排列而成(Newelletal., 1960;Rusnack,1960)。同心—放射状包壳中同时出现同心状结构和放射状结构(Sandberg,1975;Halley,1977;Medwedeff and Wilkinson,1983)。部分同心—放射状包壳中,同心状结构将放射状结构分隔在若干细层中,这些包壳同样可根据放射状晶体的粒度、形态划分为同心—放射纤维状包壳(图 3-4)和同心—放射板状包壳(图 3-5),以及内部为放射状板状结构、外部为放射纤维状结构的同心—放射板状—放射纤维状包壳(图 3-6)(Sandberg,1975;Medwedeff and Wilkinson,1983)。也有一些同心—放射状包壳由内侧的放射状包壳和外侧的同心状包壳共同构成(图 3-7,3-8)(Medwedeff and Wilkinson,1983)。由于切线状纹层以及被同心状结构分隔的放射状细层颜色较浅,而其他同心状纹层颜色较深,故同心状包壳和同心—放射状包壳常常呈现出亮层、暗层交替排列的形态(Medwedeff and Wilkinson,1983;Rankey and Reeder,2009)。
4 古代鲕粒包壳结构特征
古代鲕粒的包壳通常呈现出与现代鲕粒类似的原生结构类型和结构组合类型,放射状结构、同心状结构是常见的原生结构类型,但同心状结构中的切线状纹层极少有发现(Sandberg,1975)。由于成岩作用的影响,部分古代鲕粒包壳由次生包壳结构组成。
4.1 放射状结构
古代鲕粒包壳的放射状结构亦由纤维状晶体或板状晶体构成,放射状晶体粒度分布区间较广(Tucker,1984)。放射纤维状晶体排列紧密,放射板状晶体排列紧密或松散,松散排列的晶体之间充填泥晶、黏土等深色细粒物质(Tucker,1984;Chow and James,1987)。
4.2 同心状结构
古代鲕粒包壳的同心状结构包括与现代鲕粒相似的截面式纹层、微晶纹层、黏土质纹层等类型(Tucker,1984)。同心状结构有时因相邻纹层中的放射状晶体增生而被破坏,可能局部消失,也可能仅有少量残留(Strasser,1986;Chow and James,1987;Cantrell,2004)。切线状纹层极少在古代鲕粒中发现(Sandberg,1975;Wilkinson and Landing,1978;Algeo and Watson,1995)。
4.3 次生结构
古代鲕粒包壳中最为常见的次生结构是晶粒结构和泥晶结构(Wilkinson and Landing,1978;Wilkinsonetal., 1985;Algeo and Watson,1995)。晶粒结构也被称为砖块状(bricklike)结构、粒状镶嵌(mosaic)结构等,由粒状方解石构成,包壳可能整体完全转化为晶粒结构,也可能局部转化为晶粒结构、局部保留同心状结构或放射状结构(Wilkinson and Landing,1978;Wilkinsonetal., 1985)。晶粒结构的成因可能是原生文石被溶蚀后形成溶蚀孔,方解石以胶结物的形态充填其中(Sandberg,1975;Tucker,1984;Wilkinsonetal., 1985)。泥晶结构由分布于包壳内部、局部或完全取代原生结构的泥晶构成(Wilkinson and Landing,1978;Wilkinsonetal., 1985;Flügel,2010),成因则可能是微生物活动导致的泥晶化作用(Margolis and Rex,1971;Flügel,2010)。
部分古代鲕粒包壳中,放射状晶体粒度较粗、保存程度差,呈现出类似亮晶、微亮晶的形态,可能是由粒度较细的原生放射纤维状晶体在成岩过程中转化而成(Wilkinsonetal., 1985;Chow and James,1987;Algeo and Watson,1995)。在成岩过程中,放射状结构中的原生高镁方解石晶体可能转化为方解石,其原生形态可能随着成岩程度的提高而不断变化,原生结构保存程度逐渐减小,如放射状晶体穿透同心状结构的现象随着成岩作用的进行而增多,最终有可能使同心状结构完全消失,而且放射状晶体在成岩序列中可以呈现出由较细晶体到较粗晶体的演化趋势(Chow and James,1984;Algeo and Watson,1995)。
古代鲕粒包壳中还可能出现白云石等次生矿物(Tucker,1984;Hird and Tucker,1988)。通常粒度较小的自形—半自形菱形白云石晶体出现在包壳放射状晶体的间隙(Tucker,1984;Chow and James,1987;Hird and Tucker,1988),也有一些包壳几乎完全由数个粒度较大的自形—半自形鞍状白云石晶体构成(Tucker,1984;Hird and Tucker,1988)。白云石细晶的形成可能与原生的高镁方解石成分有关,而鞍状白云石则是在埋藏后通过交代原始矿物形成(Tucker,1984;Chow and James,1987;Hird and Tucker,1988)。
4.4 结构组合类型
古代鲕粒包壳中常见的结构组合类型为放射状包壳和同心—放射状包壳(图 3-1,3-2,3-4,3-5,3-6)。同心状包壳极少见,原因是构成同心状结构必要的切线状纹层在古代鲕粒中极少发现(Tucker,1984;Strasser,1986;Bates and Brand,1990)。与现代鲕粒类似,放射状包壳呈连续的放射状结构。同心—放射状包壳由黏土质纹层、微晶纹层、截面式纹层和被纹层分隔在细层中的放射状结构构成。
古代鲕粒包壳结构与现代鲕粒包壳结构的差异源于原生矿物成分稳定性的不同(Sandberg,1975;Wilkinsonetal., 1985)。现代鲕粒包壳切线状纹层的原生矿物成分通常为文石,放射状结构的原生矿物成分通常为文石或高镁方解石(Marshall and Davies,1975;Sandberg,1975;Halley,1977;Landetal., 1979)。由于文石稳定性差,在成岩过程中会转化为方解石,由文石构成的切线状纹层或放射状结构可能随之转化为次生的晶粒结构(Friedman,1964;Sandberg,1975;Wilkinsonetal., 1985;Budd,1988)。而由稳定性较好的高镁方解石、方解石构成的放射状结构会不同程度地保存下来(Friedman,1964;Chow and James,1987;Algeo and Watson,1995)。
5 中国古代鲕粒包壳结构研究存在的问题及实例
5.1 存在的问题
中国鲕粒包壳研究相对起步较晚,20世纪80年代开始有学者研究鲕粒包壳结构(王英华等,1983;张秀莲,1984;余素玉等,1987),并在古代鲕粒中识别出放射状、同心状等原生结构以及晶粒结构、泥晶结构等次生结构(王英华等,1983;张秀莲,1984;韦龙明,1995;沙庆安和江茂生,1998;金瞰昆,1998)。2000年以后,古代鲕粒包壳结构受到越来越多学者的重视,包壳结构类型常被作为指示沉积环境的指标之一(如刘伟和张兴亮,2010;侯恩刚等,2014;金鑫等,2015;姬国锋等,2016)。
与国外碳酸盐鲕粒研究成果相比,国内鲕粒包壳结构研究尚存在一些普遍性问题。
1)对包壳结构的识别不够精细,对其成分的认识存在误区,部分研究者对同心状结构,尤其是切线状纹层的认识存在误区。有国内研究报道了古代鲕粒保存的切线状纹层,但由于缺乏纹层内晶体沿切线方向排列的确切证据,目前还难以将这些纹层归为切线状纹层。同时,虽然多数学者认识到放射状结构由放射状排列的晶体组成,但对放射状结构细分类型特征的描述多不明确,比如未区分放射纤维状结构和放射板状结构。另外,部分国内研究对放射状晶体的矿物组成认识不一致,或推断为方解石,或推断为难以在古代鲕粒中保存的高镁方解石、文石等,缺乏可靠的判断依据。
2)对包壳结构组合类型认识不充分。国内研究者一般将包壳划分为同心状包壳、放射状包壳、同心—放射状包壳。事实上,古代鲕粒包壳最常见的原生结构组合类型应为放射状包壳和同心—放射状包壳,同心状包壳因其重要组构切线状纹层难以在地层中保存而无法识别(Sandberg,1975;Budd,1988)。部分包壳因可以观察到交互出现的暗色和浅色同心层而被划分为同心状包壳。与现代鲕粒包壳结构比较,较薄的暗色层应为同心状纹层。有研究描述浅色层具有晶粒状次生结构,原生结构无法识别,其中部分较厚的浅色层可与被同心状结构所分隔的放射状细层对比,不应被笼统归为同心状纹层。
3)对于包壳结构及其组合类型的古环境解释存在问题。鲕粒包壳的同心状结构常被学者认为指示水动力较强的形成环境。然而,现代鲕粒研究和人工合成鲕粒实验结果表明,放射状包壳可能形成于静水环境(Rusnack,1960;Loreau and Purser,1973;Sandberg,1975;Daviesetal., 1978;Landetal., 1979)。同心状结构存在多种类型,具切线状纹层的包壳可能形成于强扰动环境(Rusnack,1960;Loreau and Purser,1973;Sandberg,1975;Daviesetal., 1978;Landetal., 1979),而黏土质、有机质、截面式同心状结构的形成,则可能由磨蚀、加积等多种沉积作用形成,因此不能笼统地认为同心状结构可以指示扰动较强的环境(Sandberg,1975;Daviesetal., 1978;Helleretal., 1980;Troweretal., 2017)。由此可见,由于对古代鲕粒包壳结构认识不充分,在使用鲕粒包壳结构讨论古环境时可能产生错误解读。
5.2 研究实例
近年来,笔者对国内外多个地区的古代鲕粒包壳结构开展了专题研究,其中华南下奥陶统多个层段产出的鲕粒具有保存良好的包壳结构。华南下奥陶统鲕粒主要出现在亮晶鲕粒灰岩和泥晶鲕粒灰岩中,在亮晶似球粒灰岩、亮晶生屑灰岩和泥晶生屑灰岩中亦有少量分布,对应碳酸盐缓坡浅潮下带沉积环境(孙永超等,2016)。Liu 等(2011)发现湖北兴山地区下奥陶统鲕粒包壳原生结构保存良好,依据现代鲕粒包壳研究成果,在其中识别出放射状结构和同心状结构。
在对这些鲕粒的包壳结构特征进行细致描述的基础上,笔者在华南下奥陶统鲕粒中识别出与现代鲕粒类似的放射状和同心—放射状2种包壳类型(图 3-1,3-2,3-4,3-5)。这说明上文所建立的鲕粒包壳结构分类方案在中国古代鲕粒研究中是适用的(该研究的详细内容将另文发表)。

1—放射纤维状包壳,包壳呈连续的放射状结构,晶体呈较细的纤维状,单偏光;2—图4-1中矩形内区域,放射纤维状晶体排列紧密,少量深色细粒物质弥散分布其间,包壳由数层晶体层叠构成,单偏光;3—放射板状包壳,包壳呈连续的放射状结构,晶体呈较粗的板状,单偏光;4—图4-3中矩形内区域,放射板状晶体排列松散,晶体间由深色细粒物质填充,包壳由 1层板状晶体构成,单偏光图 4 湖北兴山古洞口剖面下奥陶统鲕粒的放射状包壳Fig.4 Non-concentric cortices of ooids,without any concentric structure but show radial structures, from the Lower Ordovician of Gudongkou section at Xingshan,Hubei Province

1—同心—放射纤维状包壳,包壳内发育同心状结构,将放射状结构分隔在不同细层中;2—图5-1中矩形内区域,细层内放射状晶体呈纤维状,同心状结构为有断口的黏土质纹层(黄色虚线),或内侧截切放射状晶体(红色箭头)、外侧发育细粒层(红色虚线)的截面—黏土质纹层,正交偏光;3—同心—放射板状包壳,包壳内发育同心状结构,将放射状结构分隔在不同细层中,鲕粒外侧 1个同心层内部呈晶粒结构(红色箭头),单偏光;4—图5-3中矩形内区域,细层内放射状晶体呈板状, 同心状结构为局部有断口的黏土质纹层(黄色虚线),正交偏光图 5 湖北宜昌黄花场剖面下奥陶统鲕粒的同心—放射状包壳Fig.5 Concentric cortices of ooids,with concentric structures and layers which show radial structure, from the Lower Ordovician of Huanghuachang section at Yichang,Hubei Province
1)放射状结构由长轴近垂直于细层或核心表面的晶体构成(图 4,图 5)。这些放射状晶体可以粒度较小、呈纤维状外形(图 4-1;图 5-1),也可以粒度较大、呈板状外形(图 4-3;图 5-3)。其中纤维状晶体排列相对紧密,深色黏土在晶体之间仅有零星分布(图 4-2;图 5-2)。纤维状晶体在同心—放射状包壳的放射状细层中或放射状包壳中通常层叠发育,长度小于细层/包壳的厚度(图 4-2;图 5-2)。板状晶体排列可能紧密也可能松散,松散排列的晶体之间富含黏土、泥晶等深色细粒物质(图 4-4;图 5-4)。单个细层/包壳内通常仅发育 1层板状晶体,板状晶体的长度等于细层/包壳的厚度(图 4-4;图 5-4)。
2)同心状结构将放射状结构分隔在细层中,常见的有 2种: 黏土质纹层和截面—黏土质纹层。黏土质纹层由黏土构成,颜色较深,呈同心圆状分布在相邻纹层之间,局部有断口、呈断续分布(图 5-3,5-4)。截面—黏土质纹层内侧为截切纹层中放射状晶体的同心圆状平整截面,外侧为深色黏土质纹层(图 5-1,5-2)。
3)根据放射状晶体的粒度,可将放射状包壳划分为放射纤维状(图 4-1)和放射板状(图 4-3) 2个亚类。同心—放射状包壳也可分为同心—放射纤维状(图 5-1)和同心—放射板状(图 5-3) 2个亚类。
6 结论
1)鲕粒包壳结构的准确识别是鲕粒成因和环境研究的关键。根据对现代鲕粒研究的历史综述和现状分析,划分出现代鲕粒的 3个研究阶段,综合分析了现代鲕粒包壳原生结构的类型和特征,讨论了包壳结构组合类型对鲕粒研究的重要性。
2)受成岩作用影响,古代鲕粒包壳中的原生结构可能被晶粒结构、泥晶结构等次生结构取代,也可能以不同的保存程度保留在包壳中。放射状结构和同心状结构均常见于古代鲕粒包壳中,但同心状结构中的切线状纹层极少能在成岩作用中保留下来。古代鲕粒常见的包壳结构组合类型为放射状包壳和同心—放射状包壳。
3)中国鲕粒包壳研究起步晚,但国内学者已对中元古界至古近系中的鲕粒进行了大量研究,识别出了放射状结构、同心状结构等包壳原生结构。国内鲕粒包壳结构研究尚存在对放射状结构识别不精确、对同心状结构和切线状纹层等存在认知误区、对包壳结构组合类型认识不充分等问题。在借鉴现代鲕粒包壳结构研究成果的基础上,笔者以华南下奥陶统鲕粒为例,细致描述了古代鲕粒中放射状结构、同心状结构等包壳原生结构的特征,区分出放射状包壳、同心—放射状包壳等包壳结构组合类型,展示了古代鲕粒包壳原生结构研究的重要科学意义。
致谢在野外工作和论文撰写过程中,得到了北京大学地球与空间科学学院王海峰、闫振、陈宇轩、李家腾、董轶婷、兰漪涟、王艺凝、王凯、裴进峰等同学的宝贵意见和建议。北京大学造山带与地壳演化教育部重点实验室贾秋月老师制作本研究所用岩石薄片,北京大学造山带与地壳演化重点实验室 SEEL(SEM-EDS-EBSD-CL)实验室张波副教授和张磊同学等在包壳显微结构的扫描电镜和EDS研究方面提供了宝贵的建议。在此一并感谢!
猜你喜欢
福利中国(2022年3期)2022-07-20
核科学与工程(2021年4期)2022-01-12
音乐天地(音乐创作版)(2021年7期)2021-10-13
中国核电(2021年2期)2021-06-04
都市(2020年8期)2020-09-06
河北画报(2020年3期)2020-06-24
河北画报(2020年5期)2020-06-10
中国金属通报(2020年1期)2020-04-23
小资CHIC!ELEGANCE(2018年1期)2018-02-02
中国CT和MRI杂志(2017年1期)2017-01-11
