
氢气生物学作用的生物酶基础
2020-02-27 06:08马雪梅张鑫谢飞赵鹏翔张昭仪杨张晓康马胜男李秦剑吕宝北刘梦昱YAOMawulikplimiAdzavon孙学军李英贤
生物技术进展 2020年1期
马雪梅, 张鑫, 谢飞, 赵鹏翔, 张昭, 仪杨, 张晓康, 马胜男, 李秦剑, 吕宝北, 刘梦昱, YAO Mawulikplimi Adzavon, 孙学军, 李英贤
1.北京工业大学生命科学与生物工程学院, 北京 100124;
2.上海交通大学氢科学中心, 上海 200240;
3.中国航天员科研训练中心, 北京 100094
氢气生物学的研究历史很短,它一直被认为是生理惰性气体,曾经作为配气在潜水中使用。1969年Levitt等[1]报道人的肠道菌会产生氢气,99%的氢气在大肠中主要由肠道厌氧细菌产生,食物可以使肠道产氢效率提高7~30倍,但那时氢气被作为代谢废气对待。1975年在人类历史上首次发现氢气的医学效应,美国学者Dole等[2]在Science报道,连续14天吸入8个大气压氢气能够明显减少小鼠皮肤癌,然而由于高压氢气难于控制,这篇文章并未引起广泛关注。日本医科大学Ohsawa等[3]于2007年在NatureMedcine上发表论文,证明吸入少量氢气(2%)可以有效保护缺血再灌注造成的大鼠脑损伤。由于氢气是一种为人们熟知的气体,反应的产物只有水,至今尚未发现对人体的毒性作用,氢气的疾病治疗作用意味着这种气体可能在医疗上具有广泛潜在的应用前景,因此这一研究迅速受到国际生物医学领域的广泛关注。大量的研究证明氢气对逾百种疾病具有潜在的治疗作用,如各类缺血再灌注损伤、动脉硬化和糖尿病等代谢类疾病、神经退行性疾病症状的改善、肿瘤防治、抗炎和戒毒等[4]。氢气对植物也有广泛的生物学效应,包括在植物抗盐、抗铝毒、镉毒、汞毒、缓解百草枯带来的氧化损伤等非生物胁迫反应中具有保护作用[5]。
氢气生物学虽然快速发展,但是关于氢气发挥生物学作用的机理一直都有争论。目前主要有三种假说:一是被广泛接受的氢气选择性清除毒性自由基的假说;二是生物酶假说,作者团队首先报道了氢气促进乙酰胆碱酯酶活性[6],随后证明氢气可以作用于辣根过氧化物酶(HRP)、线粒体复合物Ⅰ等生物酶(未发表数据,2018年氢分子生物医学分会学术年会大会报告),氢气与生物大分子的作用值得关注;三是信号分子假说,由于缺乏明确的直接作用靶点,该假说不作为本文讨论的重点。
1 氢气发挥生物学作用的机理
1.1 氢气清除自由基假说
Dole等[2]认为氢气具有还原性,氢与羟自由基(hydroxyl radical,·OH)的反应是治疗肿瘤的基础;Ohsawa等[3]提出氢气发挥功能的基础是选择性清除·OH和过氧亚硝酸根(peroxynitrite,ONOOH)等毒性自由基。之后的很多研究都证明在氢气的作用过程中有自由基的减少,与活性氧相关的SOD、CAT、GSSH、MDA等也发生变化,氢气的抗氧化作用得到广泛认可。但是,氢气的抗氧化作用是直接与自由基反应清除已经产生的自由基还是减少自由基的生成?目前被广泛接受的是氢气选择性清除自由基的理论,但本团队认为这一理论很难成立,主要是基于现有的研究并不能充分证明氢气与自由基在生理条件下能够直接反应。
1.1.1氢气与自由基反应常数 细胞内能够与·OH发生反应的成分,如膜脂、GSSH等,不仅含量远大于氢气,而且反应常数(1×1010M-1·s-1)也远远大于·OH和氢气的反应常数(4.2×107M-1·s-1) 。因此,氢气与·OH的反应常数太低并不足以支持其优先于体内大量存在的高反应常数的底物发生反应,特别是在氢浓度不高的情况下[7]。
1.1.2无细胞体系中氢气与羟自由基的反应
Ohsawa等[2]证明氢气与·OH直接反应的体外体系是芬顿反应,然而芬顿反应在不同[Fe2+]/H2O2比值下·OH产生量会受到氧气浓度的极大影响[8]。本团队采用不同[Fe2+]/H2O2比值,以氮气组为对照,HPF荧光法检测·OH的量(未发表数据,2017年中国医促会氢分子生物医学分会学术年会大会报告),发现在H2O2浓度为1 mmol·L-1([Fe2+]/H2O2为1∶10) 时的HPF荧光信号强度高于H2O2浓度为5 μmol·L-1([Fe2+]/H2O2为20∶1) 时的HPF荧光信号强度,这说明H2O2浓度为1 mmol·L-1时产生了比较多的自由基。但是氢气组的HPF荧光信号与对照组相比没有降低,表明氢气并没有与·OH反应;H2O2浓度为5 μmol·L-1时,氢气组的 HPF荧光信号与对照组相比降低,但是由于加入气体可能影响溶液的含氧量,本团队以通入氮气的溶液作为对照,发现氮气组的HPF信号也降低了(未发表数据,2017年氢分子生物医学分会学术年会大会报告)。说明在此条件下,·OH产量的变化容易受到氧气 浓度的影响。因而我们认为Ohsawa等提出的氢气与·OH直接反应的体外体系由于H2O2的浓度很低,自由基的产生容易受到反应条件的影响,并不能充分证明氢气与·OH发生了反应。Penders等[9]的研究也证明氢气不能通过与ONOOH反应发挥功能。
1.1.3细胞中氢气与羟自由基的反应 Ohsawa等[2]证明氢气清除·OH的三个细胞学实验都是选用 PC12细胞作实验对象,用线粒体呼吸链抑制剂antimycin A诱导产生活性氧(ROS),通过HPF荧光和ESR发现氢气可以减少·OH。在这些实验中,自由基信号的减弱既可能是因为自由基的产生减少了,也可能是因为自由基产生之后被清除了,这些实验结果以及和自由基相关的SOD、CAT、GSSH、MDA等指标的变化并不能分辨自由基产生和清除的区别,因此尚需要更进一步的证据证明。
1.1.4体外超声乳化体系中氢气与羟自由基的反应 另一个证据证明氢与·OH直接反应的研究是氢气对白内障超声乳化治疗导致的角膜损伤的保护作用[10]。超声乳化治疗白内障过程中,超声波诱导羟基自由基的形成,能破坏角膜内皮细胞,研究采用ESR 和HPF荧光检测发现氢能减少·OH,从而减少内皮氧化损伤。超声反应会有空化效应,即会产生很多小的气泡,空化气泡内极高的温度(几千摄氏度)和压力(数百个大气压)导致水分子发生热分解,进而在气泡边界产生·OH,这种高温高压的特殊微环境下,氢气与·OH反应能力极大增强,从而增加了氢气与·OH反应的可能性;同时,加入氢气的排氧效应也可以减少·OH产生[11]。因而,实验有可能观测到加入氢气后·OH的减少。这个研究属于特殊的体外条件下氢气和自由基反应的例子,并不能代表氢气在生理状态下与自由基的反应。
总之,现有的关于生理条件下氢气与自由基直接反应的证据并不充分,多数属于间接证据,无法区分氢气是与自由基直接反应还是影响了自由基的产生。
1.2 生物酶假说
乙酰胆碱酯酶(acetylcholinesterase,AchE)是存在于胆碱能突触间的神经传导中的一种关键酶,该酶能降解乙酰胆碱、终止神经递质对突触后膜的兴奋作用,并保证神经信号在生物体内的正常传递。越来越多的证据表明,AchE在神经再生、神经退行性疾病等方面发挥重要作用[12]。本团队2014年发现氢气可以在体外与AchE作用,提高酶活性[6],这也是到目前为止氢与生物酶直接作用的唯一报道。AchE的活性中心含有由Ser200、His400和 Glu327 组成的催化三联体,即“His-Ser-Glu催化铁三角”(图1)[12],氨基酸之间的连接与电子转移都是靠氢键等次级键,氢气可能在氢键的形成与电子重排中发挥作用。
为了更方便地研究酶与氢气的相互作用,本团队选择辣根过氧化物酶(horseradish peroxidase,HRP)作为研究对象,HRP是以铁卟啉为辅基的血红素蛋白,同类蛋白包括细胞色素氧化酶、过氧化物酶、细胞色素P450超家族、一氧化氮合酶等众多重要的生物酶[13],这类酶的活性中心含有铁卟啉,在含有过氧化氢的溶液中酶的活性中心可以产生氧化性很强的高价铁自由基中间态。如果氢气可以直接与活性中心的高价铁自由基反应,HRP的酶活应该降低。但本团队研究发现,加入氢气后HRP的酶活性升高,荧光光谱和紫外光谱研究发现氢气可以影响酶活性中心的微环境(未发表数据)。因而推测,由于氢气分子很小,其可以在分子表面,也可以进入分子内部,通过氢键等弱相互作用影响酶活性,AchE和HRP酶活的升高可能与此有关。
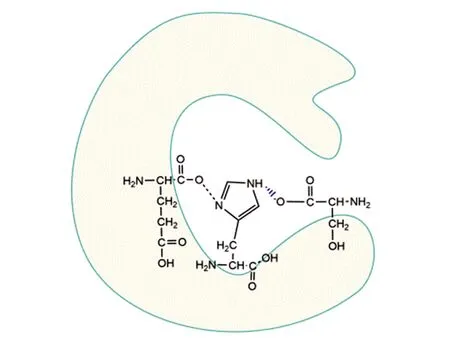
图1 乙酰胆碱酯酶的活性中心[12]
2 高等生物中的氢化酶活性
氢化酶是存在于细菌、古细菌和藻类中的一类金属酶,主要存在于自然界厌氧微生物体内,它能够催化氢气的氧化或者质子的还原这一可逆的化学反应:H2H++H-2H++2e-[14]。
地球大约形成于46亿年前,那时候地球温度很高,从火山中喷出的气体,如水蒸气、氢气、氨、甲烷、二氧化碳、硫化氢等,构成了原始的大气层。地质证据表明生命开始于地球诞生10亿年之后,早期地球的大气中充满氢气,含量达到40%,是一个适宜生命生长的环境,氢化酶在这个时期的原始生命产生过程中被“创造”出来是合乎逻辑的[15]。目前证明,有大量微生物可以利用氢气作为能源,也有很多微生物可以产生氢气,这得益于它们体内的氢化酶。
真核生物的起源有多种理论,Martin等[16]认为氢气是推动真核生物进化的关键因素。他们认为真核生物是由一种厌氧的、依赖氢气的严格自养的古细菌(宿主)和一种能够呼吸、厌氧异养代谢产生分子氢作为废物的真细菌(共生体)共生而产生的,随着地球环境氢气来源的减少,宿主会严格地依赖于异养真细菌共生体,这提供了一种强大的选择力,不可逆转地将共生体和宿主联系起来,如果共生体逃逸,宿主会立即饿死。因此这些宿主紧紧地依附于共生体,并且尽量选择大表面积的宿主细胞形状包围共生体(而不是内吞共生体,因为细菌没有细胞骨架),增加接触面,以便获得更多的H2和CO2。宿主比表面积的增加使其自身受益,但这种环境对共生体的代谢造成损害,也降低了共生体产生氢来为宿主提供燃料的能力。随着适应环境的进化选择,宿主可以利用自身与环境的表面摄入有机质并提供给共生体,宿主对氢的代谢依赖性消失了,厌氧环境的限制也消失,成为异养类型,共生体成为现在线粒体、氢化酶体等的前体。如果这一进化路径是正确的,真核生物共生体的氢气代谢功能可能不会丧失,而是留在线粒体等结构中随时可能因为环境的变化而恢复。
真核生物线粒体的电子传递链是能量代谢的核心部位,复合物Ⅰ(NADH:泛醌氧化还原酶,EC 1.6993)在线粒体以及细菌的呼吸链中广泛存在,可以催化NADH的氧化和泛醌的还原以及质子的泵出。线粒体中,三羧酸循环和脂肪β-氧化等过程的脱氢酶催化反应脱下的2H是由NAD+接受,形成NADH,复合物Ⅰ由基质接受还原型NADH中的2个H和2个e传递给黄素单核苷酸(FMN)形成FMNH2,FMNH2再经一系列铁硫中心(FeS),将电子传递到泛醌(CoQ)。每次传递电子过程同时可偶联将4个质子从内膜基质侧泵到内膜胞质侧,形成膜电位推动ATP的产生。
线粒体复合物Ⅰ由NADH脱氢酶模块(N-module,NuoE、NuoF和NuoG亚基)、醌模块(Q-module,NuoC、NuoI、NuoB和NuoD亚基)和质子泵模块(P-module, NuoA, H, J, K, L, M 和N亚基)组成(图2A),复合物Ⅰ与氢化酶的同源性和进化关系已有许多报道[17-21]。复合物Ⅰ的核心是由14个“核心”亚单位(7个亲水亚单位和7个疏水亚单位)组成的L型复合物,这些亚基从原核生物到真核生物的线粒体都是保守的,真核生物的线粒体复合物Ⅰ含有31个额外的辅助亚单位。形成复合物Ⅰ的14个“核心”亚单位与氢化酶的大/小亚基相比较在结构上高度保守,包括N2簇(复合物Ⅰ中NADH脱氢酶模块末端FeS簇)、质子通道和相似的催化中心结构。N-module的NuoE、NuoF有NADH结合位点、FMN和FeS簇,NuoG与唯铁氢化酶(Fe-only hydrogenase)类似,在复合物Ⅰ中提供FeS簇参与电子传递。存在于许多厌氧或兼性厌氧微生物中的能量转换氢化酶(energy-converting hydrogenase, Ech)与复合物Ⅰ的“核心”亚基序列上高度相似[17]。此外,复合物Ⅰ还与一类膜结合的能将底物氧化和氢还原活化质子转运的镍铁氢化酶相似[18]。醌模块可以接受来自N模块的电子并传递给CoQ,NuoB和NuoD亚基与可溶性镍铁氢化酶(NiFe-hydrogenases)的大小亚基相似,CoQ的结合位点与氢化酶中的NiFe活性位点相对应[19-21]。
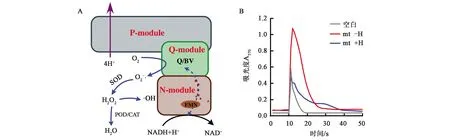
A:线粒体复合物Ⅰ的结构模式图; B:以BV作为电子受体检测氢对离体线粒体复合物Ⅰ电子传递的影响
3 细胞的氢代谢
3.1 线粒体的氢代谢
3.1.1线粒体复合物Ⅰ的产氢活性 高等生物产生氢气在植物中早有报道,1961年Sanadze[22]首次证明植物的叶可以吸收和释放氢气,后来发现小麦、大麦、玉米、拟南芥等都可以产生氢气。虽然有研究证明叶绿体具有产生氢气的能力[23],但是对于线粒体是否能产生氢气还没有报道,线粒体复合物Ⅰ一直被认为失去了氢代谢的能力。结构分析认为复合物Ⅰ可能具有氢代谢的能力,但是一直缺乏实验证据。本团队研究发现,线粒体复合物Ⅰ可以产生氢气,并且产氢的位点在FeS簇末端N2传递电子至CoQ的位置,氢代谢与TCA循环的代谢调节偶联,特别与TCA代谢中间物琥珀酸、ɑ-酮戊二酸、延胡索酸以及CoQ密切相关[24]。
3.1.2氢气影响线粒体复合物Ⅰ的酶活性 复合物Ⅰ催化线粒体内电子由供体NADH传递到CoQ受体的能量转移反应。加入抗霉素A和叠氮化钠分别抑制复合物Ⅲ和复合物Ⅳ的活性,在340 nm检测NADH氧化后峰值的降低确定复合物Ⅰ的活性,溶液中溶入氢气可以排氧,为测试缺氧条件下的酶活性,用氮气处理组作为对照。结果显示,氢气组的酶活性明显高于氮气组(图3),这也表明在缺氧情况下氢对复合物Ⅰ的活性有保护作用(未发表数据,2018年氢分子生物医学分会学术年会大会报告)。
3.1.3氢气可以提高线粒体膜电位 线粒体内膜的质子泵可将基质内质子泵入膜间隙,质子跨膜转运使得线粒体膜间隙积累大量质子,形成跨线粒体内膜的电位 (ΔΨ),简称为线粒体膜电位(mitochondrial membrane potential,MMP)。正常线粒体膜电位是维持线粒体进行氧化磷酸化、产生ATP的前提,人体的ATP有95%为线粒体所提供。线粒体复合物Ⅰ是主要的质子泵,每传递一对电子就会泵出4个质子。本团队测定了多种细胞的线粒体膜电位,细胞在含氢培养基中培养1 h,JC-1染色后检测线粒体膜电位,发现氢处理可以明显增强细胞线粒体的膜电位(图4),结果表明氢促进了质子梯度的形成,点燃了线粒体跨膜电位的“生命之火”,但这一过程并不增加活性氧水平(未发表数据)。
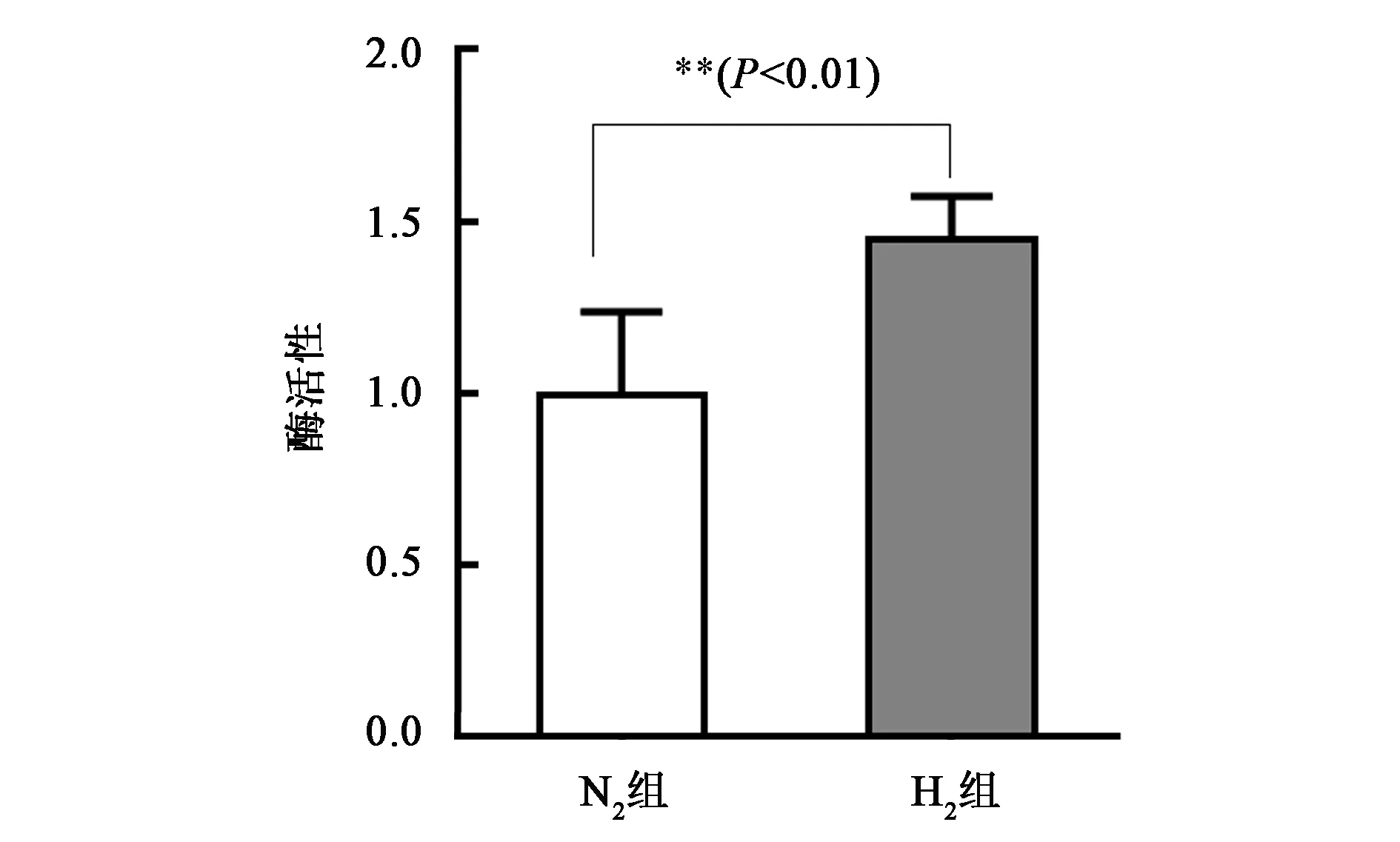
图3 氢气影响线粒体复合物Ⅰ的酶活性
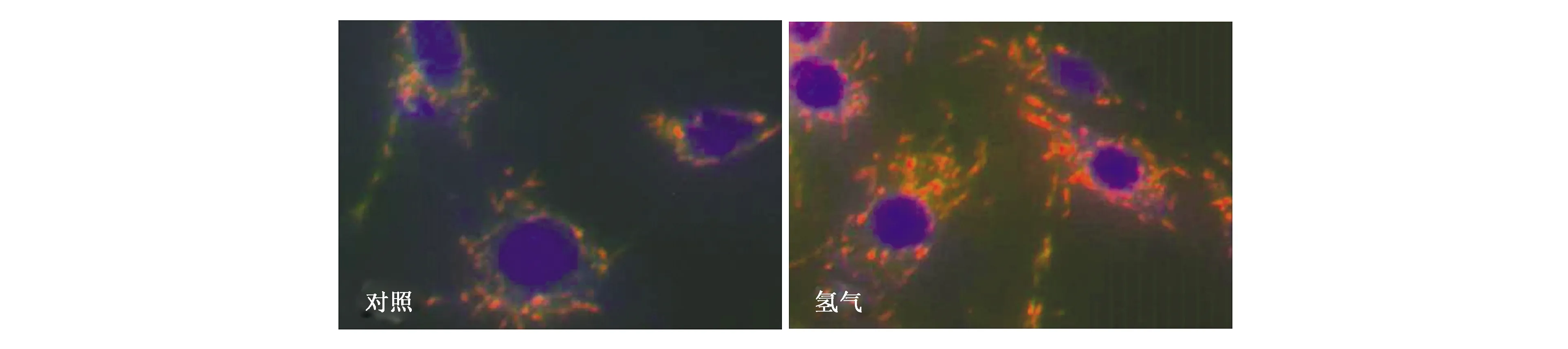
图4 氢提高C6细胞的线粒体膜电位
3.1.4氢气与线粒体活性氧的产生 活性氧(reaction oxygen species, ROS)在胞内信号转导、细胞应答、损伤修复、衰老等过程中发挥着重要的作用。在线粒体的电子传递过程中,电子沿着一系列的电子传递体传递给氧生成水。但是,位于呼吸链底物端的一些物质,如黄素蛋白、非血红素铁蛋白(non-heme iron proteins)、醌,尤其是半醌等发生氧化还原的能障是很低的,常常直接将电子传给O2产生超氧阴离子(O2-),再被SOD等转化为H2O2进而形成·OH等活性氧(图2A)。线粒体中FMN和NADH脱氢酶的铁硫中心是O2-产生的主要部位,特别是复合物Ⅰ对CoQ 的过度还原会形成大量的活性氧[25]。
氢气被认为有清除·OH的作用,我们认为氢气在生理条件下并不能与·OH直接反应,但大量的研究又证明氢气有抗氧化作用,因而我们推测氢气有可能在源头减少自由基的产生,也就是在电子传递给O2产生 O2-这一步发挥作用,即具有在自由基产生之初关上“阀门”的开关作用。
为了验证这一假设,本团队以苄基紫精(benzyl viologen,BV)作为电子受体代替CoQ检测氢气的作用(图2A)。由于CoQ的吸收峰在270~280 nm处,与蛋白质的吸收峰重合,因此很难通过检测CoQ的还原来研究电子在复合物Ⅰ的传递过程。BV接受电子的位点主要位于复合物Ⅰ的铁硫簇,其吸收峰在570~600 nm,本团队在研究中用BV代替CoQ作为电子受体检测电子从NADH通过复合物Ⅰ的传递过程。反应中加入抗霉素A和叠氮化钠分别抑制复合物Ⅲ和复合物Ⅳ活性,加入Na2S2O4启动反应,在570 nm处实时检测还原型BV的信号。结果显示,加入氢气后可以明显抑制电子传递到BV(图2B),表明氢气可能在电子经过复合物Ⅰ的传递过程中抑制活性氧的产生(未发表数据,2018年氢分子生物医学分会学术年会大会报告)。
BV是PD、AD等神经退行性疾病的诱发剂,可以从线粒体复合物Ⅰ和Ⅲ接受电子传递给O2产生O2-等活性氧自由基,对细胞造成损害,氢气对BV接受电子的抑制也表明氢气在对抗活性氧产生方面的功能,特别在活性氧爆发时,这种源头控制显得尤为重要。本团队用电镜研究氢气对线粒体的保护作用,有机磷农药毒死蜱造成脑神经元线粒体减少、溶胀、基质空间增大、嵴的破坏和非晶基质密度的出现,氢气可以保护线粒体,明显减少损伤,增加线粒体的数量[6],这些结果为氢气对抗毒性物质损害提供了依据。
3.1.5氢气对细胞的代谢重编程 线粒体在生命活动中发挥着非常重要的作用,作为生物体重要的能量工厂,线粒体是氧化磷酸化的场所,同时也负责连接糖酵解、氨基酸代谢、核苷酸代谢等重要的生命过程。氢气对线粒体的作用必然影响细胞的代谢和基因表达调控,如氢气与线粒体复合物Ⅰ的作用会改变三羧酸循环(TCA)α-酮戊二酸、琥珀酸等的代谢,这些物质是HIf-1、α-酮戊二酸依赖的非血红素铁氧化酶的重要调节物,也可以调节TET (ten eleven translocation) 家族的表达,通过甲基化等表观遗传修饰发挥作用[26-28]。
肿瘤在很多情况下是一种代谢障碍性疾病,肿瘤细胞线粒体功能受损,主要利用糖酵解途径生长。本团队研究脑胶质瘤时发现,氢气对肿瘤细胞的杀死作用有限,但可以诱导肿瘤干细胞分化[29],进一步的研究表明,这一过程中肿瘤细胞由于氢气的作用发生了代谢重编程(未发表数据)。这可以进一步解释为什么氢气对众多的肿瘤都能发挥作用,同时也可以解释对于不同代谢类型的肿瘤,氢气的作用会有差别。
3.2 细胞膜的氢代谢
细胞膜(cell membrane)又称细胞质膜(plasma membrane,PM),是防止细胞外物质自由进入细胞的屏障,它保证了细胞内环境的相对稳定。质膜上存在的氧化还原系统,如NADH-氧化还原酶类、NADPH-氧化酶、单脱氢抗坏血酸还原酶、苹果酸脱氢酶等,与许多细胞功能如质子分泌、离子吸收、生长控制、信号转导、病原微生物的防御反应等生理功能密切相关[30]。细胞跨膜质子流动在细胞生长、离子跨膜运输、细胞pH稳态调节等重要生理活动中均发挥着重要作用。
膜结合氢化酶(membrane-bound hydrogenase, MBH)存在于许多微生物中,如大肠杆菌(Escherichiacoli)、巴氏甲烷八叠球菌(Methanosarcinabarkeri)等[14,31],但高等生物的细胞膜还没有代谢氢气的报道。本团队研究发现纯化的植物质膜囊泡能够产生氢气,并且氢气明显影响质膜上NADH-铁氰化钾氧化还原酶的活性,进一步利用非损伤微测技术(non-invasive micro-test technique,NMT)对植物细胞进行实时检测,发现氢气能够显著影响细胞表面的质子泵出,以及钠、钾等离子的流动[32]。
细胞膜内外电荷差异在各种细胞上具有极其重要的生理功能,神经细胞是研究细胞膜电位最佳的选择。本团队进一步采用膜片钳技术测定氢气对活体脑片动作电位的影响,循环缓冲液等气 体流速通入二氧化碳、氧气、氢气混合气30 min确保饱和,测试过程中持续通入;使用膜片钳在全细胞模式下记录大脑皮层动作电位(AP)。结果显示氢气组与对照组相比大鼠皮层神经细胞的静息膜电位和动作电位阈值发生改变,氢气处理组静息膜电位差升高(图5A),神经细胞爆发动作电位阈值升高(图5B),表明氢气可能对神经细胞膜离子通道的开放和关闭有影响(未发表数据)。有研究表明氢水能减少小鼠吗啡成瘾戒断症状,减少戒毒痛苦[33];Mizuno等[34]的研究显示氢水能够降低焦虑、抑郁和神经自主功能,对神经兴奋具有一定的抑制作用,此外也有氢气改善睡眠的报道。结合本团队上述脑片膜片钳结果所显示的氢气可以调节神经细胞膜电位,推测氢可能影响神经细胞的兴奋传导功能,这可能是氢气发挥神经调节作用的基础。
以上这些结果进一步证明细胞膜的氢代谢是氢气可能的靶点之一,这为拓展氢气的研究靶点提供了参考。
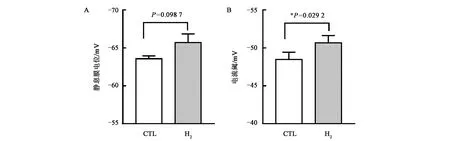
A:静息条件下神经细胞膜电位; B:外加电压刺激下神经细胞动作电位阈值
4 展望
氢分子具有重要的生理功能。氢与羟自由基的直接反应可能不是其发挥生物学功能的主要方面;我们不否认氢具有抗氧化的作用,但氢气的抗氧化作用属于在自由基产生的源头进行控制;酶的活性中心具有特殊的微环境和高反应活性,是氢分子作用的关键点,包括线粒体膜的氧化还原酶类特别是复合物Ⅰ、细胞膜的氧化还原酶类及离子通道等都受到氢气的调节,这表明氢气的作用可能是多靶点且主要基于酶学反应的过程。氢气作为能源其反应产物是水,是属于未来的清洁能源;氢气在生物体内最终的代谢归宿也是水,这区别于所有的药物,它同样是属于未来的可能改变人类健康模式的气体。
猜你喜欢
石油石化绿色低碳(2022年4期)2023-01-06
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
中成药(2018年7期)2018-08-04
中成药(2018年3期)2018-05-07
中成药(2017年5期)2017-06-13
医学研究杂志(2015年12期)2015-06-10
中国药业(2014年21期)2014-05-26
科学启蒙(2013年2期)2013-07-11
