
Comparative proteomic analysis of mature and immature oocytes in domestic cats
2020-02-20 16:10BongkochTurathumKulnasanSaikhunSittirukRoytrakulChinnaratChangsangfaSupitaTanasawetMorakotSroyrayaYindeeKitiyanant
Bongkoch Turathum, Kulnasan Saikhun, Sittiruk Roytrakul, Chinnarat Changsangfa, Supita Tanasawet,Morakot Sroyraya, Yindee Kitiyanant,
1Department of Anatomy, Faculty of Science, Mahidol University, Rama VI Road, Bangkok 10400, Thailand
2Department of Basic Medical Science, Faculty of Medicine Vajira hospital, Navamindradhiraj University, Bangkok 10300, Thailand
3Institute of Molecular Biosciences, Mahidol University, NakhonPathom 73170, Thailand
4National Center for Genetic Engineering and Biotechnology, National Science and Technology Development Agency, Pathumthani 12120, Thailand
5Department of Anatomy, Faculty of Science, Prince of Songkla University, Hat Yai, Songkhla 90112, Thailand
ABSTRACT Objective: To evaluate changes of feline (Felis catus) oocytes proteins during in vitro maturation by using the proteomic approach.Methods: Immature oocytes (germinal vesicle) isolated from female cats were cultured and collected at 0 h and 24 h. After collection, oocytes were investigated into immature (germinal vesicle) and mature (metaphaseⅡ) stages. The qualitative profiles of the proteins at the immature and mature stages were determined by one-dimensional electrophoresis and liquid chromatography-mass spectrometry.Results: Our data revealed that following 24 h in vitro maturation the maturation rate (metaphaseⅡstage) was 58.7%. Eighty-one of the 260 proteins analyzed were differentially expressed between the germinal vesicle stage and the metaphaseⅡ-arrest stage. Proteomic analysis of germinal vesicle and metaphaseⅡoocytes showed abundant expression of proteins involved in transportation (10%),indicating that this was a major characteristic of germinal vesicle oocytes. Similarly, analysis of the proteome of metaphaseⅡoocytes indicated that cell cycle proteins were overexpressed. Interestingly,proteins involved in DNA repair and apoptosis were only expressed in germinal vesicle oocytes and proteins involved in fertilization were only expressed in metaphaseⅡoocytes.Conclusions: The overexpression of certain proteins in germinal vesicle and metaphaseⅡis necessary for oocyte development and maturation. Our findings provide a valuable resource for further investigations into protein expression in oocytes at different developmental stages.
KEYWORDS: Domestic cats; Immature oocytes; Mature oocytes;Proteomics
1. Introduction
Most wild members of the felidae family are vulnerable,threatened, or in danger of extinction in nature. The domestic cat(Felis catus) serves as an animal model in the reproductive studies of endangered or nondomestic species. In vitro maturation (IVM) is a technique that allows saving the genetic materials from endangered species. The oocyte maturation process has been described by the changes in chromosomal morphology in the meiosis stage[1]. Oocytes are arrested at the germinal vesicle (GV) stage which is the first meiotic prophase. Upon the surge of gonadotropin secreted by the anterior pituitary gland, the immature oocytes have been stimulated to resume the first meiosis, ovulate and thereby arrest at metaphaseⅡ(MⅡ)[2]. Approximately 80% or less cultured cat oocytes achieve nuclear maturation and only 60% of the mature oocytes are fertilized by in vitro fertilization (IVF)[3].To understand the molecular mechanisms of oocyte biology,it is important to recognize the processes that regulate meiotic maturation of oocytes. Oocyte maturation is a complex process that involves the regulation of protein synthesis, degradation, and phosphorylation[4]. The processes of cellular differentiation and maturation are characterized by specific protein expression[5].Many proteins with well-defined functions have been identified during oocyte maturation. For example, high levels of glucose-6-phosphate dehydrogenase are essential for viable oocytes and for the generation of triphosphopyridine nucleotide, which is required in the process of fertilization[6]. Nucleoplasmin 2 and peptidylarginine deiminase 6 are proven maternal-effect proteins,which play crucial roles in early embryonic development[7,8].Peroxiredoxin 2, glutathione-S-transferase, and myomegalin 1 are involved in redox regulation and the cAMP-dependent signaling pathway. Both of them have been found to be correlated with oocyte maturation[9,10]. The activation of some protein kinases plays a key role in the meiotic maturation of oocytes. Several studies have investigated mammalian oocyte proteomics, including the exploration of bovine[11], pig[12], and mouse[13] oocyte proteins. The processes of IVM and IVF of oocytes collected from excised ovarian tissue have reached a level of consistency in certain species, to allow replacement of the costly and laborintensive processes of in vivo embryo production and recovery[14].The ability to grow and fertilize immature oocytes is beneficial to produce large numbers of embryos for developmental biology,cryopreservation, and genetic studies, as well as for live animal production. The domestic cat is an important model to study human genetic diseases and to develop the assisted reproduction in taxonomically-related endangered species. Proteomic analysis is a valuable technique that can be applied to differentiate protein expression between different stages of nuclear maturation of oocytes[5]. This information is helpful for further understanding the mechanism of oocyte maturation, which will improve the quality of oocytes after IVM. However, no previous reports have revealed proteomics-based investigations of fresh immature and mature oocytes in the felid family. Therefore, this study was to employ proteomic analyses to identify the proteins necessary for IVM of feline oocytes.
2. Materials and methods
2.1. Chemicals
Chemicals used in this study were purchased from Sigma (Sigma,St. Louis, MO, USA), unless indicated otherwise. Media were prepared once a week, filtered, and then kept in sterile bottles.
2.2. Oocyte collection
Prior to ovarian tissue collection, animal care and ethics approval was informed to Mahidol University Ethic Committee on Animal research. However, no ethical approval was required due to ovaries were collected after ovariohysterectomy for the purpose of permanent contraception from the veterinary clinic of the Veterinary Public Health Division, Bangkok Metropolitan Administration (total number of cats = 150). To collect cumulusoocyte complexes (COCs), ovaries from normal females of various breeds (> 6 months old) were repeatedly sliced in Petri dishes which contained TCM 199 (Invitrogen, Carlsbad, CA, USA)supplemented with 25 mM 2-hydroxyethyl, 0.1% polyvinylalcohol,0.1 mM glutamine, 2.5 mM sodium pyruvate, and 1% penicillinstreptomycin. COCs with more than three layers of cumulus cells with a darkly pigmented oocyte cytoplasm (grade 1 and grade 2)were selected for the experiments. Grade 1 COCs with more than 5 layers of compact cumulus cells and grade 2 COCs with 3 to 5 layers of compact cumulus cells[15].
2.3. IVM
The COCs were cultured in Dulbecco's modified eagle medium supplemented with follicle-stimulating hormone, luteinizing hormone, and estradiol. Ten COCs were cultured in Petri dishes containing 100μL culture medium under mineral oil in each drop for 24 h at 38.5 ℃ in 5% CO2.
2.4. Assessment of nuclear maturation
After 24 h of culture, COCs were denuded by exposure to 0.5%hyaluronidase for 5 min and gently pressed the pipette to remove cumulus cells. To analyze the stages of oocytes, the denuded oocytes were characterized as being in MⅡ-arrest stage according to the presence of the first polar body within the perivitelline space under a stereomicroscope (200×, Nikon SMZ1500, Japan).
2.5. Protein extraction and one-dimensional sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)
A total of 900 GV and MⅡ denuded oocytes were lysed by adding 0.5% SDS supplemented with protease cocktail inhibitor. Lysates were centrifuged at 17 530×g, at 4 ℃, for 20 min. The supernatant was collected and the protein concentration was determined by Lowry method. The protein lysates were mixed with 5× sample buffer and heated at 95 ℃ for 10 min before loading onto a 12.5%gel for SDS-PAGE. Electrophoresis was performed at 70 V in electrophoresis buffer and the gel was then silver-stained[15].
2.6. Gel slicing and tryptic in-gel digestion
To perform in-gel digestion of proteins, 20 μL of trypsin solution (10 ng/μL trypsin in 50% acetonitrile/10 mM ammonium bicarbonate) was added to the gels, followed by incubation at room temperature for 20 min. To keep the gels immersed throughout digestion, 30 μL of 30% acetonitrile was added and incubated overnight. To extract the peptide digestion products,30 μL of 50% acetonitrile in 0.1% formic acid was added to the gels and incubated with shaking for 10 min, and this was repeated three times. Extracted peptides were collected, dried by vacuum centrifugation, and stored at -80 ℃ for further mass spectrometric analysis. Prior to liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis, the peptides were dissolved in 20 μL 0.1% formic acid[15].
2.7. LC-MS/MS analysis
Peptide solutions were analyzed by using the HCTultra PTM Discovery System (BrukerDaltonics Ltd., UK) coupled to the UltiMate 3000 LC System (Dionex Ltd., UK). Peptides were separated on a nanocolumn (PepSwift monolithic column 100 μm i.d. 50 mm). Eluent A contained 0.1% formic acid and eluent B contained 80% acetonitrile/water with 0.1% formic acid. Peptide separation was carried out with a linear gradient from 10% to 70% eluent B at a flow rate of 300 nL/min for 13 min, including a regeneration step at 90% eluent B and an equilibration step at 10% eluent B, which took 20 min. Peptide fragment mass spectra were achieved in data-dependent AutoMS mode with a scan range of 300-1 500 m/z, three averages, and up to five precursor ions selected from the MS scan 50-3 000 m/z[15,16].
2.8. Protein quantitation and identification
DeCyder MS Differential Analysis software (DeCyderMS, GE Healthcare) was used to quantify the protein. The analyzed MS/MS data from DeCyderMS were submitted for database searches by using the Mascot software (Matrix Science, London, UK). The data were searched in the NCBI database for protein identification.Data normalization and quantification of the changes in protein abundance between the GV and MⅡ stages were performed and visualized by using MultiExperiment Viewer (Mev) software version 4.6.1. Data were normalized and quantified the changes of protein abundance between the GV and MⅡ, subsequently visualized by using MultiExperiment Viewer (Mev) software version 4.6.1. Gene ontology annotation including molecular function and biological process was assigned to the proteins identified according to the Uni-Prot database. The identified proteins were then submitted to the search tool STITCH (V4.0) to gain insight into their cellular functions and to annotate all of the functional interactions among proteins in the cell[15,16].
2.9. Statistical analysis
Statistical tests of the variance (ANOVA) tests of differences for these data sets were performed for statistically significant proteins.P<0.05 was considered statistically significant.
3. Results
3.1. IVM of feline oocytes
To verify the changes of protein during IVM, a total of 1 533 oocytes were cultured in vitro to induce oocyte maturation. The different stages of oocytes during IVM were referred to the GV,GV breakdown, MⅠ, and MⅡ stages, which were classified by morphological observations. Following 24 h of IVM, cat oocytes were divided into the GV and MⅡ stages by morphological analysis (Figure 1A and B). In Dulbecco's modified eagle medium supplemented with follicle-stimulating hormone, luteinizing hormone, and estradiol, the maturation rate was 58.7%. Based on morphological analysis, the GV and MⅡ stages of IVM were collected and samples were prepared for protein fractionation by SDS-PAGE and analysis by LC-MS/MS.
3.2. Quantitative proteome profile of feline GV and MII-arrested oocytes
A representative SDS-PAGE image from 900 GV and MⅡ-arrested oocytes were shown in Figure 2A. Gels were sliced into 15 pieces and assigned to in-gel tryptic digestion. Peptides were analyzed by LC-MS/MS. A total of 1 702 protein identifications were made from 20 μg of protein. Of these 1 702 proteins, 1 442 proteins were found in both the GV and MⅡ stages and 260 proteins were differentially expressed between GV and MⅡ stages (Figure 2B).
All 1 702 proteins were analyzed to determine the gene ontology annotations for biological processes and cellular compartments.The majority of proteins were found to involve in the metabolism(4.11%), transportation (4.00%), transcription (3.06%), and cell cycle (2.76%) (Figure 3A). Classification based on the subcellular localization (Figure 3B) indicated that 7.68% of proteins were from the plasma membrane and 7.31% were from the nucleus.
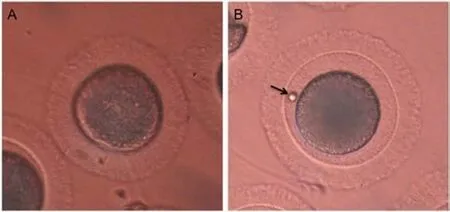
Figure 1. Morphological characterization of feline oocytes during in vitro maturation. (A) An intact germinal vesicle; (B) MetaphaseⅡ stage with extrusion of the first polar body (arrow).
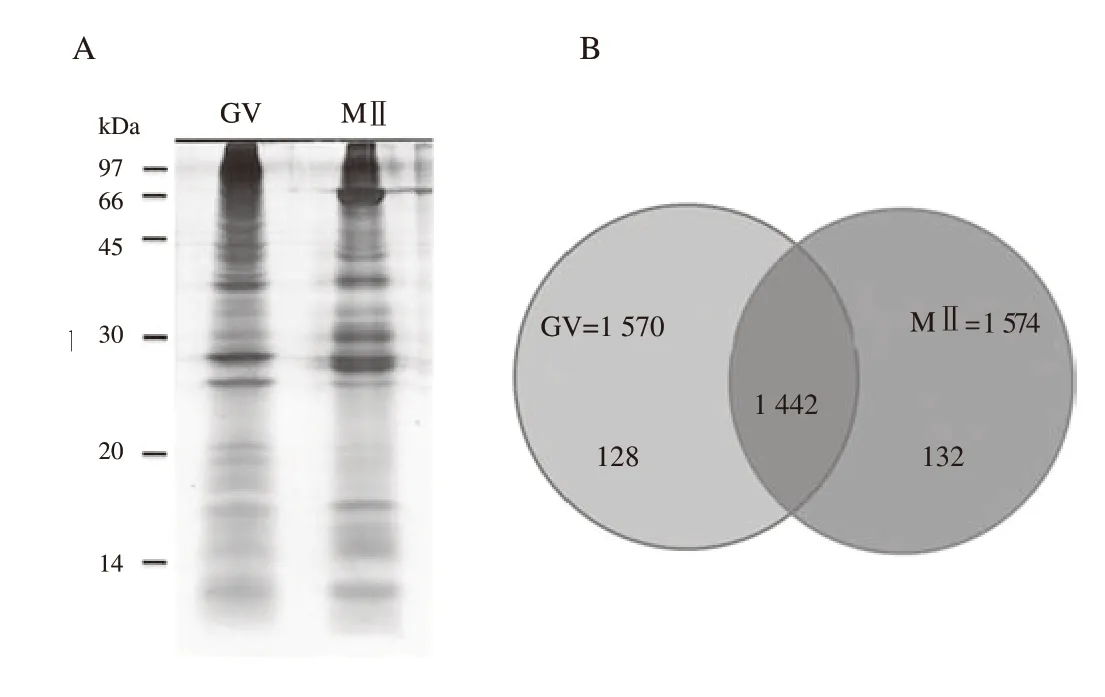
Figure 2. SDS-PAGE fractionation of denuded germinal vesicle (GV) and metaphaseⅡ (MⅡ) oocyte proteins. Pooled protein (20 μg of denuded GV or MⅡ oocyte protein) is run on a 12.5% acrylamide gel and then subjected to silver staining. A: Lane 1: total protein of GV oocytes; Lane 2: total protein of MⅡoocytes. Each gel lane is excised and then subjected to ingel digestion prior to LC/MS-MS analysis. B: 1 442 proteins are found in both the GV and MⅡ stages and 260 proteins were differentially expressed between GV and MⅡ stages.
A total of 260 proteins showed expression differences between the GV and MⅡ stages during IVM. Of these, 81 proteins showed at least two unique tryptic peptides with protein ID scores >10.Changes in protein expression over time in GV stage compared with MⅡ stage were evident using Multi-Experiment Viewer(MeV, version 4.9.0) software. In the MⅡ stage of feline oocytes during IVM, 41 proteins were overexpressed and 40 proteins were downregulated, the details and biological functions of which were presented in Table 1 and 2, respectively. Proteins participating in transportation, signal transduction, and cell cycle events were found to predominate among overexpressed proteins in GV stage. Compared with MⅡ oocytes, GV oocytes contained higher levels of proteins involved in transportation including inward rectifier potassium channel 9, stonin-2, lysosomal-associated transmembrane protein 5-like protein, and Shaw type potassium channel (Table 1 and 2). These results suggested that the feline IVM process may be dependent upon 81 specific proteins that were differentially expressed. These proteins might play an important role in the molecular events involved in feline oocyte development.
3.3. Classification of proteins identified in GV and MII stage oocytes
The 81 proteins that were differentially expressed during oocyte IVM were subjected to classify according to their associated biological processes and molecular functions using information based on the Gene Ontology and Uni-Prot databases.
The proteins that were overexpressed in MⅡ during IVM were clasified into the following biological functions: cell cycle 13%,metabolic process 10%, cellular organization 10%, development 8%, transcription 8%, signal transduction 8%, immune response 5%, transportation 5%, stress response 3%, translation 3%,fertilization 2%, differentiation 2%, and unknown function 23%.The upregulated proteins included the following: activin receptor type-1C, KIAA0445 (Rootletin), centromere protein T, claspin,DNA topoisomerase 2-alpha, rotatin, growth/differentiation factor 7, and titin all involved in the cell cycle and development; C-1-tetrahydrofolate synthase and cytoplasmic involved in biosynthesis;delta-like protein 1 precursor and zonapellucida sperm-binding protein 4 precursor involved in fertilization; complement factor H-like protein, tumor necrosis factor receptor superfamily member 10B-like protein, and interleukin-34 precursor involved in the immune response and apoptosis; ethanolamine-phosphate cytidylyltransferase-like protein, beta-1,4-N-acetyl-galactosaminyl transferase 3, galactocerebrosidase, UDP-GalNAc:beta-1,3-Nacetylgalactosaminyl-transferase 2-like protein, and adenosine triphosphate-dependent RNA helicase DDX47-like protein involved in metabolic processes; high mobility group B2-like protein, ninein isoform 2, and espin-like protein involved in cell organization; cryptic-like protein, 52 kDa repressor of the inhibitor of protein kinase, and torsin-1A-interacting 2-like protein involved in signal transduction; histone-lysine N-methyltransferase SETD1A isoform 2, snRNA-activating protein complex subunit 3, RNA-binding protein 14, and 40S ribosomal protein S3-like isoform 2 involved in transcription and translation; solute carrier family 25 member 39 and nuclear pore membrane glycoprotein 210 precursor involved in transportation.
The downregulated proteins in MⅡ during IVM were classified into the following biological functions: cell cycle 10%, signal transduction 10%, transportation 10%, transcription 8%, metabolic processes 8%, cellular organization 5%, apoptosis 5%, DNA repair 3%, tumor suppressor 3%, translation 3%, biosynthesis 2%,development 2%, and unknown function 31%. The downregulated proteins included the following: cyclin-dependent kinase 5 and ABL1 enzyme substrate 1 (CABLES1), synaptonemal complex protein 1, and sarcoma antigen NY-SAR-48, part involved in the cell cycle; alkyl dihydroxyacetone phosphate synthase and carnitine O-palmitoyl-transferase 1 involved in metabolic processes; hypothetical protein LOC100017349 and protein yippee-like 3 involved in apoptosis; Rho GTPase activating protein 17, signal-induced proliferation-associated 1-like protein 2, and serine/threonine-protein phosphatase 4 regulatory subunit 4 involved in signal transduction; zinc finger protein castor homolog 1-like protein (CASZ1), zinc finger protein 226-like protein,and sex-determining region Y protein involved in transcription;60S ribosomal L13a-like protein and diphthamide biosynthesis 1-like protein involved in translation; lysosomal-associated transmembrane 5-like protein, inward rectifier potassium channel 9, and Shaw type potassium channel involved in transportation;vascular endothelial growth factor receptor-2 involved in angiogenesis; and transmembrane 187-like protein and KIAA1110 protein involved in cellular organization.
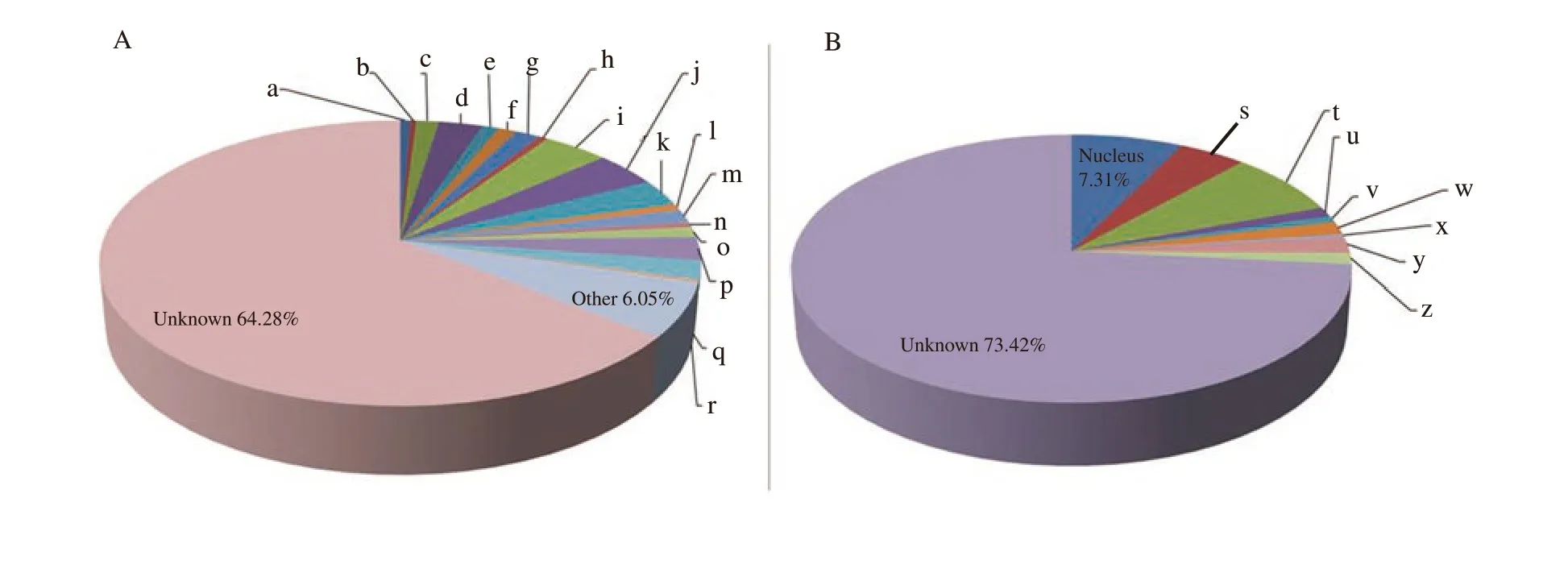
Figure 3. Gene ontology analyses of proteins identified in both germinal vesicle and metaphaseⅡ stages. Proteins are classified according to A) biological processes, and B) cellular compartment. Results are displayed as percentage of genes classified into a category over total number of class hits. a: apoptosis 0.65%; b: angiogenesis 0.35%; c: biosynthesis 1.41%; d: cell cycle 2.76%; e: cell adhesion 1.06%; f: DNA repair 1.12%; g: development 1.59%; h:differentiation 0.53%; i: metabolism 4.11%; j: transportation 4.00%; k: transcription 3.06%; l: translation 0.71%; m: immune response 1.59%; n: Oxidative stress response 0.53%; o: proteolysis 1.18%; p: signal transduction 2.64%; q: cellular organization 2.17%; r: fertilization 0.24%; s: cytoplasm 4.40%; t:plasma membrane 7.68%; u: endoplasmic reticulum 1.18%; v: Golgi apparatus 0.74%; w: mitochondria 1.61%; x: lysosome 0.31%; y: extracellular space 1.98%; z: cytoskeleton 1.36%.
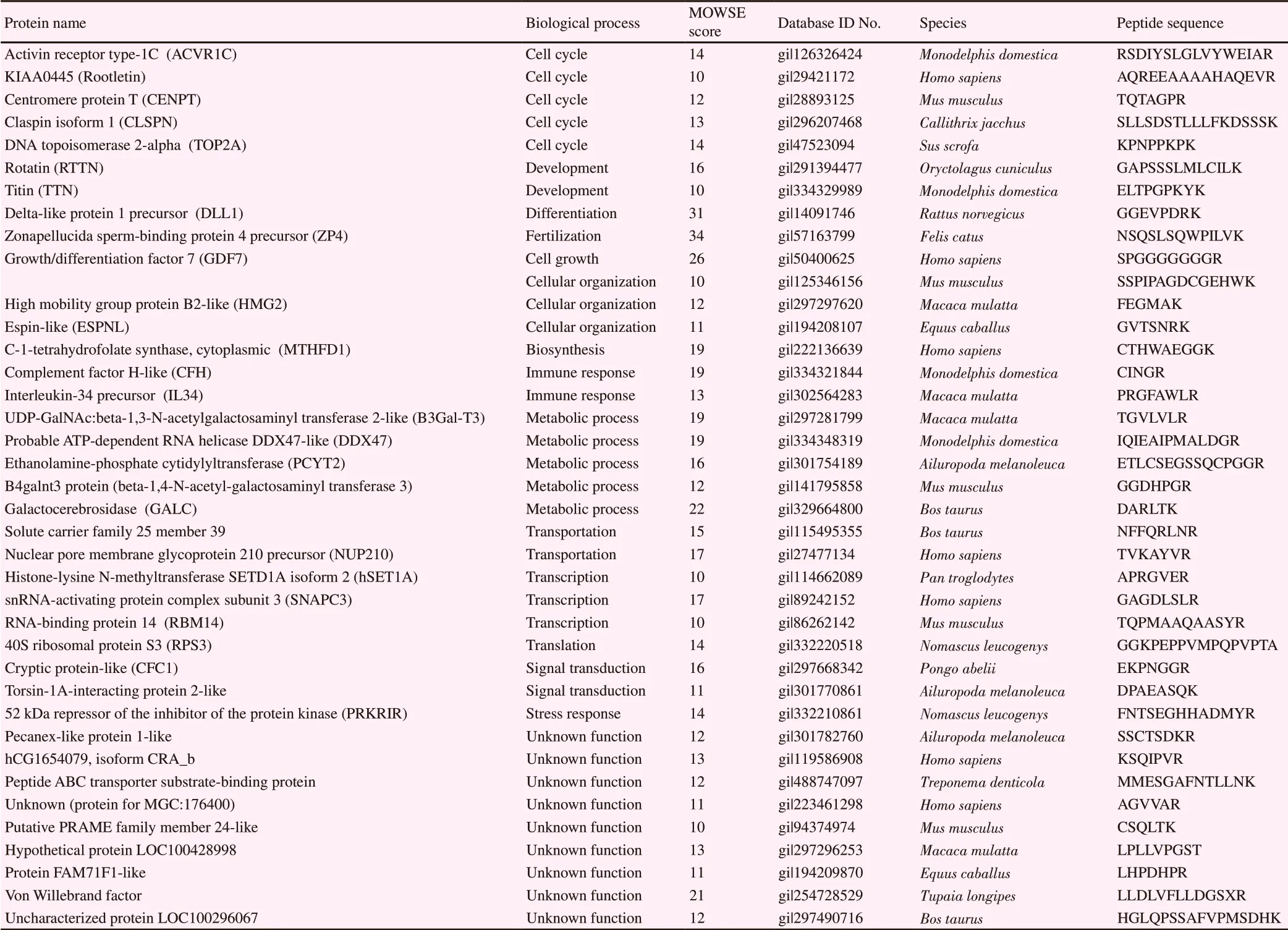
Table 1. Overexpressed proteins in metaphaseⅡ of in vitro maturation.
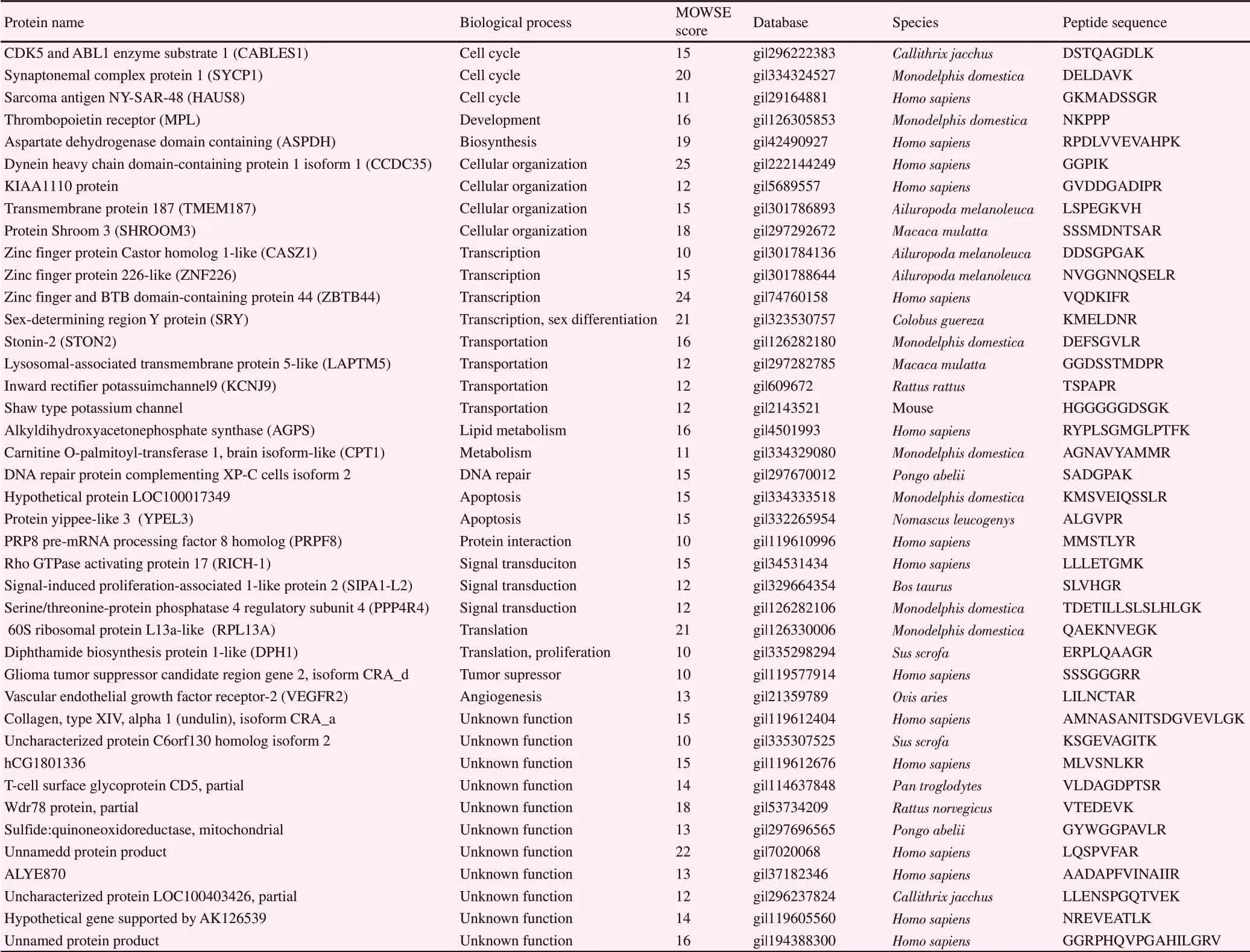
Table 2. Downregulated proteins in metaphaseⅡ-arrest stage of in vitro maturation.
4. Discussion
The complex process of oocyte nuclear maturation involves dynamic regulation of protein synthesis, degradation, and phosphorylation. Before IVM, oocytes remain at the diplotene stage of prophaseⅠand after 24 h of culture, the rate of maturation was found to be 58.7%. To improve the rate of maturation and subsequent fertilization, it is essential to understand the molecular mechanisms and proteins associated with meiotic maturation.
In this study, 81 differentially expressed proteins were identified by LC-MS/MS of GV- and MⅡ-stage oocytes. The results of this study was consistent with that of Wang et al[17] who reported that primary transporters and cation channel family members were more abundant in GV oocytes than in MⅡ oocytes.
In addition to proteins involved in the cell cycle, proteins involved in signal transduction and transcription were also identified in GV oocytes. CABLES1, Synaptonemal complex protein 1 (SCP1),and sarcoma antigen NY-SAR-48 were involved in the cell cycle. Cyclin-dependent kinase 5 and CABLES1 were cell cycle regulatory proteins that interact with both p53 and p73[18,19] and modulate the activity of female germline stem cells and oocytes. In mice, knockout of CABLES1 led to an increase in atretic immature oocytes within the ovaries and an increased occurrence of degenerating oocytes[20]. SCP1 is a key component of the protein complex that retains recombining chromosomes in prophaseⅠof the first meiotic stage in germ cells[21]. SCP1-deficient ovaries exhibited completion of meiotic prophaseⅠthat endowed oocytes with the capability to orchestrate follicle assembly in rat ovaries[22].
Rho GTPase activating protein 17, signal-induced proliferationassociated 1-like protein 2 and protein phosphatase 4 were involved in cellular signal transduction. Rho GTPase activating protein acts as a molecular switch regulating the actin cytoskeleton[23]. Rho GTPase was necessary for oocyte polar body emission and spindle rotation during meiosis in mouse oocytes[24]. Protein phosphatase 4 is a member of the serine-threonine phosphatases, which hydrolyze and remove phosphate groups from phosphoproteins and therefore antagonize protein phosphorylation and have been involved in regulating oocyte meiosis in mice[25]. Protein phosphatase 4 regulates the activity of histone deacetylase 3[26] and histone deacetylase 3 is implicated in cell cycle progression, proliferation,and differentiation during oocyte maturation[27].
CASZ1, zinc finger protein 226-like protein, and sex-determining region Y protein were involved in transcription. CASZ1 is a conserved transcription factor required for vascular patterning[28].The formation of the vascular system is important for embryonic development and homeostasis. In the absence of CASZ1, Xenopus embryos failed to develop a branched and lumenized vascular system, and CASZ1-depleted human endothelial cells displayed considerable changes in adhesion, morphology, and sprouting.CASZ1 was found throughout the developing myocardium and was downregulated in cells that re-enter the cell cycle[29].
Mature oocytes contain the complement of maternal proteins essential for fertilization and egg-embryo transition. In this study, the majority of proteins found to be overexpressed in MⅡstage participated in cell cycle events including: activin receptor type-1C, KIAA0445 (Rootletin), centromere protein T, claspin,and DNA topoisomerase 2-alpha activin receptor type-1C has been detected in several types of cells including granulose cells,cumulus cells, and oocytes[30]. Activin is secreted by granulosa cells, acting on the oocyte and granulosa cells through typeⅠand typeⅡ activin receptors[31]. Activin also promotes IVM and IVF in primate oocytes[32]. Centromere protein-T is a member of the centromere proteins essential for the attachment of microtubules to chromosomes, which occurs in the kinetochore[31]. Kinetochores are key structures in oocytes that control chromosome alignment leading to the completion of meiosis. Alterations of centromere proteins (in kinetochores) relate to the motion of chromosomes during pig oocyte maturation[33]. In human cells, ectopically localizing the N-termini of centromere protein-T and centromere protein-C to chromatin engages sufficient centromere components to drive the formation of pseudokinetochores that can bind microtubules and enhance chromosome segregation when the N-termini of centromere protein-T was mutated contributing to defective kinetochores[34]. Claspin is a checkpoint mediator protein that functions during cell cycle arrest in response to inhibit DNA replication. Its function is to phosphorylate and activate checkpoint kinase 1 for the regulation of DNA replication. In human cells,claspin and checkpoint kinase 1 are essential for the normal rates of replication fork progression[35]. In Xenopus egg extracts, Claspindepleted extracts were not able to arrest the cell cycle in response to DNA replication[36]. DNA topoisomeraseⅡ is an enzyme that activates DNA replication and chromosome segregation, and becomes localized and functions during oocyte maturation, egg activation, and embryo development by playing an important role in chromosome condensation and separation, and as a decatenation checkpoint during oocyte meiosis in mice[37].
Additionally, proteins involved in cellular organization, proteins involved in cellular development and transcription were also identified in MⅡ stage. High mobility group B2-like protein,ninein isoform 2, and espin-like protein were involved in cell organization. Ninein is associated with the centrosome throughout the cell cycle, where it binds and stabilizes the minus ends of microtubule anchoring them at the centrosome[38]. The espins,microfilament binding proteins, constitute an emerging family of actin-binding and actin-bundling proteins[39]. Rotatin, titin, and Delta-like protein 1 are proteins involved in cellular development.Rotatin is an essential protein for determination of the key leftright specification in vertebrate embryos. In mice, embryos deficient in rotatin showed randomized heart looping, delayed neural tube closure, and failure to undergo the critical step of axial rotation as a result of an embryonic defect[40]. Titin is a calciumresponsive protein and calcium currents are responsible for oocyte meiosis resumption. The expression of titin in human trophoblasts is recognized to participate in the processes of placentation and embryo development[41]. Delta-like protein 1 is a member of the epidermal growth factor-like family. It is widely expressed in embryonic tissues, as well as the ovaries of adult tissues[42]. The function of Delta-like protein 1 is still unknown.
In conclusion, the identification of differentially expressed proteins in GV and MⅡ stages aids our understanding of the processes of meiotic maturation and fertilization. Our current finding provides a valuable resource for further investigations into the functions of proteins specifically expressed in oocytes at different developmental stages.
Conflict of interest statement
We hereby confirm there are no conflicts of interest associated with this publication.
Acknowledgements
The authors would like to thank the Veterinary Clinic of the Veterinary Public Health Division, Bangkok Metropolitan Administration for kindly providing cat ovarian tissue and Proteomic Research Laboratory Unit, National Science and Technology Development Agency (BIOTEC), Pathumthani,Thailand, for assisting LC-MS/MS analysis. We also thank Kate Fox, DPhil, from Edanz Group (www.edanzediting.com/ac) for editing this manuscript.
Funding
The study was supported by a grant from the Royal Golden Jubilee, RGJ, Thailand (Grant number: PHD/0350/2551).
Authors' contributions
Bongkoch Turathum participated in all aspects of the experiment and writing the manuscript. Sittiruk Roytrakul contributed to the proteomic analysis. Chinnarat Changsangfa, Morakot Sroyraya and Supita Tanasawet were involved in oocyte collection, IVM, protein extraction and analysis. Kulnasan Saikhun and Yindee Kitiyanant designed the experiment and contributed to the analysis and discussion of data. All authors read and approved the final manuscript.
 Asian Pacific Journal of Reproduction2020年1期
Asian Pacific Journal of Reproduction2020年1期
- Asian Pacific Journal of Reproduction的其它文章
- Testosterone is a surrogate and proxy biomarker for severity of late-onset preeclampsia: A cross-sectional study
- Assessment of antioxidant status of women with polycystic ovarian syndrome
- Novel genetic variants of transferrin receptor 2 exon 4 and cytokines profile of anemic and nonanemic pregnant women in Central Java, Indonesia
- Estrogenic activity of hydroalcoholic extract of Clitoria ternatea Linn. leaves on rats
- Residual impact of 17α-methyltestosterone and histopathological changes in sexreversed Nile tilapia (Oreochromis niloticus)
- Identification of pathogenic microorganisms of repeat breeder dairy cows and a hyperimmune treatment approach