
水汽压差与钾素互作对高温下番茄营养吸收与光合特性的影响
2020-02-03 03:16:00张嘉宇焦晓聪宋小明丁崌平李建明
干旱地区农业研究 2020年6期
张嘉宇,焦晓聪,宋小明,丁崌平,柏 萍,李建明
(西北农林科技大学园艺学院, 陕西 杨凌 712100)
饱和水汽压差(VPD,vapor pressure difference)可以用来表征空气的干燥程度,是影响植物生长发育的重要环境因素之一。番茄 (SolanumlycopersicumL.)是一种喜温性的蔬菜,在夏季高温季节栽培时经常会受到高温胁迫。伴随夏季高温经常出现湿度低于30%的干燥环境,空气干燥不仅对植物的营养状况产生抑制,也会影响植株进行光合作用[1];前人研究表明,生长在高VPD下的植物,其生长和产量会受到极大的抑制,而通过降低VPD可以有效缓解水分胁迫,维持气孔功能,进而提高番茄植株的光合作用和品质产量[2]。因此, 通过对VPD的优化调控可以改善植物在高温逆境下的表现。
VPD已被广泛认为是水分运输的蒸发驱动力,在大气-植物-土壤连续体中,VPD的改变会影响植物的水分运输,进而影响植物体内营养元素的吸收和分配。许多学者对VPD如何影响植株的营养吸收进行了一系列研究。Kupper等[3]和 Sellin等[4]的研究表明,白桦和杂交白杨长期暴露于高湿环境下会降低叶片的氮、磷含量; Lihavainen等人[5]指出降低VPD可以改善植物氮代谢的平衡,进而影响植物对环境变化的抵抗力;另外也有研究表明,白天大气湿度的升高会增加夜间水通量的潜力,促进了白桦树叶片对氮、磷的吸收与积累[6]。然而,目前的研究主要集中在氮素上,有关钾素的研究较少。钾(K)是植物必需的营养元素之一,参与许多影响植物生长发育的生理生化过程。在高温胁迫下施用钾肥可以调节植物细胞的渗透势和膨压[7],调控物质代谢和光合作用相关酶的活性,进而提高植物的光合和抗热性[8]。在设施蔬菜生产中普遍存在重氮、磷肥轻钾肥的现象, 严重影响了植株对氮、磷元素的吸收,因此研究钾素与其他营养元素之间的关系,确定适宜的钾素水平就显得十分重要。此外,关于VPD与钾素交互作用对设施番茄植株生长的影响目前未见文献报道。
因此,本文深入探讨了高温环境下VPD与钾素互作对番茄营养吸收和光合响应的影响,对番茄幼苗光合响应、CO2响应、气体交换参数和氮磷钾元素含量及积累量的变化等进行了研究,以期改善高温胁迫下番茄的营养与光合状态,探寻适宜的钾素供应水平,为提高钾素利用率以及合理施肥提供理论依据。
1 材料与方法
1.1 试验地点与材料
试验于 2018年9—10月在西北农林科技大学南校区进行。供试番茄品种为‘金棚14-6’,种子播种于人工气候室中(昼/夜温度为25℃/18℃,相对湿度为65%/80%,光周期为12 h,光合有效辐射为400 μmol·m-2·s-1)。4周后,幼苗3叶1心时,将其移栽至规格为13 cm(高)×17 cm(直径)的小花盆中,内装基质(珍珠岩和蛭石以体积比2∶1 混合组成)470 g,盆上覆有锡箔纸,防止土壤表面水分蒸发。之后开始进行温度、VPD处理,缓苗3 d后浇灌不同浓度的营养液,此时记为第0天。每隔3 d浇一次,每次浇营养液100 ml;根据植株蒸腾量,每天补充相当于蒸腾量(重量测定)的水,确保将基质水含量保持在田间持水量的85%~90%。间隔10 d用纯水清洗一次基质,防止栽培基质中盐分增加过多,对根系造成影响;处理30 d后取样并测量各生长指标。
1.2 试验设计
试验在2个人工气候室(浙江求是AGC-P系列通用型培养架气候室,中国杭州) 内进行,白天低水汽压差(LVPD)和高水汽压差(HVPD)处理的相对湿度分别维持在(70±5)%和(30±5)%(LVPD/HVPD:1.69/3.66 kPa),夜间湿度均设定为(75±5)%(VPD:0.56 kPa)。昼/夜温度设定均为35℃/18℃。其中白天最高气温出现在14∶00,夜间最低温度出现在5∶00左右;且生长室内的温度变化为梯度升高或降低;所有处理的光照时间为 14 h·d-1(6∶00—20∶00),光合有效辐射(PAR)设定为1 000 μmol·m-2·s-1,其他时段为 0(夜间)。
每个人工气候室内设3个钾素水平, 以日本山崎(番茄)配方营养液为基础,K+浓度上下浮动,3个水平营养液中K+浓度分别为2、4、8 mmol·L-1(分别表示为K2、K4、K8),每个处理25棵幼苗,采用完全随机设计。营养液中其它成分含量参照日本山崎番茄配方,用KCl调整营养液中K元素浓度,使用尿素CO(NH2)2作为低钾水平的补充氮源。营养液配方具体情况见表1。
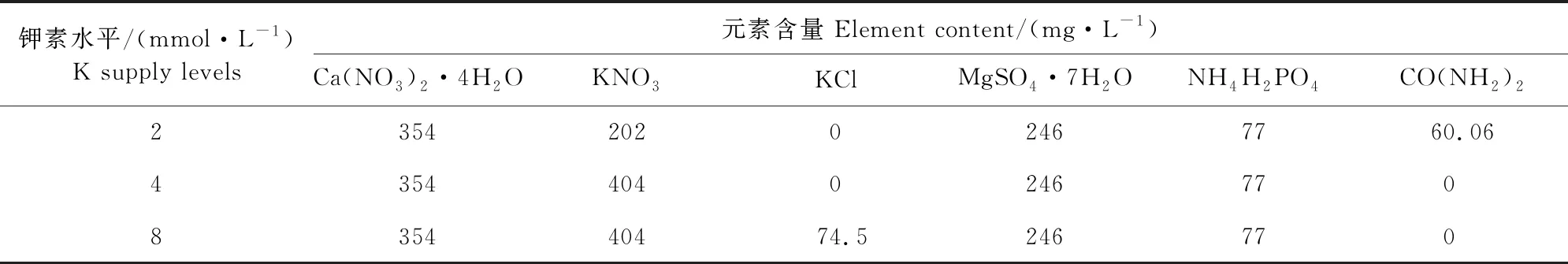
表1 不同供钾水平下的营养液配方(大量元素)
1.3 测定项目及方法
1.3.1 植物干重和氮、磷、钾元素的测定 幼苗处理30 d后随机整株取样,每处理取5株。将番茄样品植株按根、茎、叶分开,在105℃下杀青30 min,然后于75℃下烘干至恒重,用天平称量干重。将各处理烘干称重的干样先用微型植物粉碎机混合、粉碎,再用浓H2SO4-H2O2消煮,消煮液用于氮、磷、钾元素的测定。全氮和全磷含量采用高分辨自动化学分析仪(AA3,Seal,Germany)测定,全钾含量采用火焰光度计(M410, Sherwood, Britain)测定,并根据下式进行计算:
元素积累量=元素含量×干物质质量
(1)
1.3.2 气体交换参数的测定 利用Li-6800型便携式光合仪(Li-COR,Inc,USA,配置红蓝光源和 CO2注入系统)测定番茄的光合指标。在处理的第30天选取长势一致的番茄功能叶片,每个处理选5株,每株测定3片叶,共15次重复。设定光强1 000 μmol·m-2·s-1、CO2浓度400 μmol·m-2·s-1、叶室温度35℃;测定光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr) 等参数。
1.3.3 光合响应参数的测定 从各处理分别挑选5株长势基本相同的植株来测定光响应参数,测定部位为完全展开的新叶。采用Li-6800光合测定仪配置的AQ自动测量程序进行测定,CO2浓度用缓冲钢瓶控制在(400±5) μmol·mol-1,叶室温度为35℃,利用红蓝光源,设置12个光合有效辐射(PAR)梯度(1 800、1 600、1 200、1 000、800、600、400、200、150、100、50、20、0 μmol·m-2·s-1),以PAR为横轴绘制光响应曲线,根据Ye等[9]的直角双曲线修正模型获得表观光量子效率(AQY)、最大净光合速率(Pmax)、暗呼吸速率(Rd)、光饱和点(LSP)、光补偿点(LCP)等相关生理参数。
1.3.4 CO2响应参数的测定 从各处理分别挑选5株长势基本相同的植株来测定CO2响应参数,测定部位为完全展开的新叶。采用Li-6800光合测定仪配置的ACI自动测量程序进行测定,设置光强为1 000 μmol·m-2·s-1,叶室温度为35℃,同时利用CO2注入系统设置400、300、200、100、50、400、400、600、800、1 000、1 200、1 500的CO2浓度值,以CO2浓度为横轴绘制CO2响应曲线,根据叶子飘等[10]的直角双曲线修正模型获得初始羧化速率(CE)、最大净光合速率(Pmax)、CO2补偿点(CCP)、CO2饱和点(CSP)和光呼吸速率(Rp)等相关生理参数。
1.3.5 叶片水势的测定 每个处理随机挑选5株幼苗,每株测定3片叶,切下叶片,用压力室(PMS-1000,Corvallis, OR, USA)测量其叶水势。
1.4 数据处理
试验数据用SPPS 19.0软件进行统计分析,Duncan新复极差法进行差异显著性检验,采用Excel 2010和Origin 2017作图。
2 结果分析
2.1 VPD与钾素互作对番茄干物质和叶片水势的影响
由图1可知,VPD与钾素对于植株干物质和水势均存在显著的交互作用。与高VPD相比,低VPD在3种钾素水平下显著增加了番茄幼苗的干物质,随着钾素水平的提高,分别增加了47.6%、66.8%和22.3%,在中钾时达到最大值,干物质为12.73 g;而在高VPD下,植株干物质会随着钾素水平的增加而增加,在高钾时达到最大值,干物质为8.76 g。另外,叶片水势也呈现出相似的趋势:低VPD下番茄幼苗的叶片水势显著高于高VPD,且中钾时存在最大值,为-0.74 MPa, 而在高VPD下,叶片水势在各钾素水平下不存在显著差异。
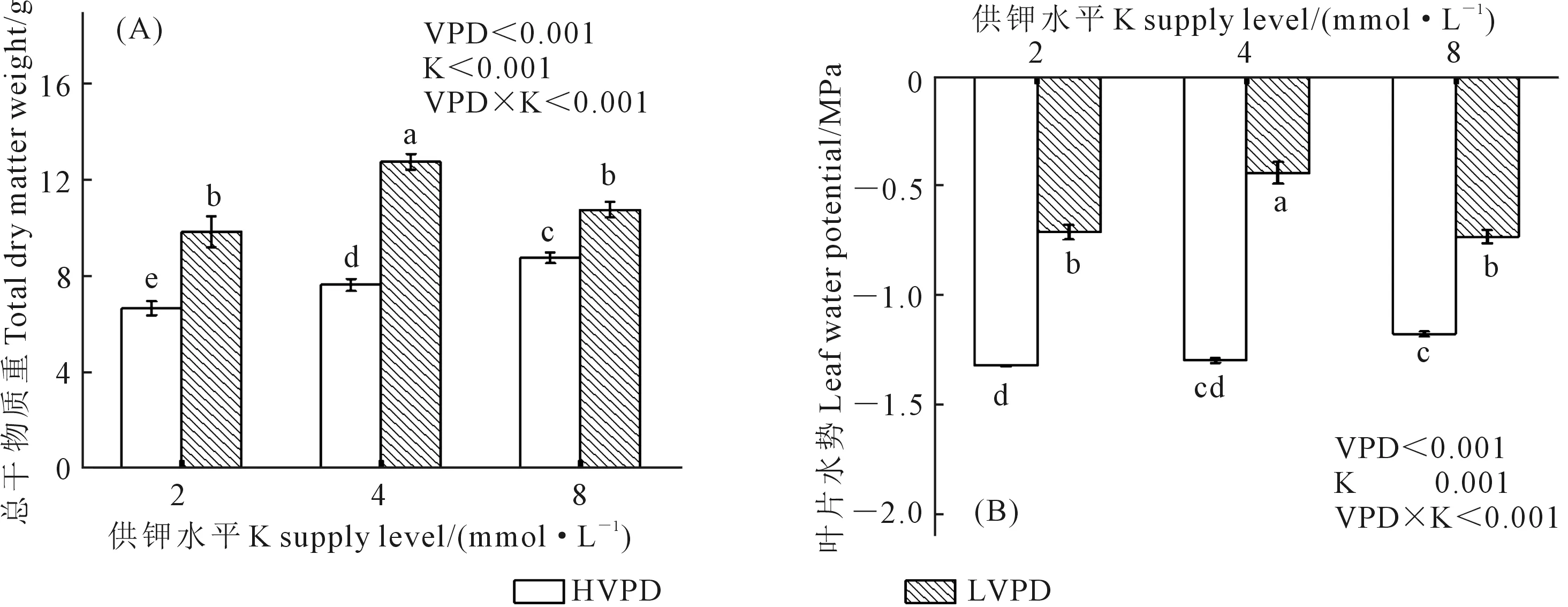
注:图中不同小写字母表示处理间在0.05水平差异显著。下同。Note:Different lowercase letters in the figure indicate significantly different among different treatments at the 0.05 level. The same below.
2.2 VPD与钾素互作对番茄各器官氮、磷、钾含量的影响
由图2可知,与高VPD相比,低VPD显著降低了中钾与高钾水平下番茄根的钾含量,但对低钾水平下的钾含量无显著影响;低VPD在各钾素水平下均显著降低了番茄叶片的钾含量,且随着钾素水平的增加,叶片钾含量分别减少了17.2%、15.1%和6.2%;对于茎的钾含量来讲,其值受钾素水平调控,而VPD的变化对其影响不显著。在各钾素水平下,低VPD显著降低了番茄各器官的氮含量,同时,也降低了番茄根与叶的磷含量,但对于茎中磷含量来讲,低VPD显著增加了低钾时的磷含量,而显著降低了中钾与高钾时的磷含量。
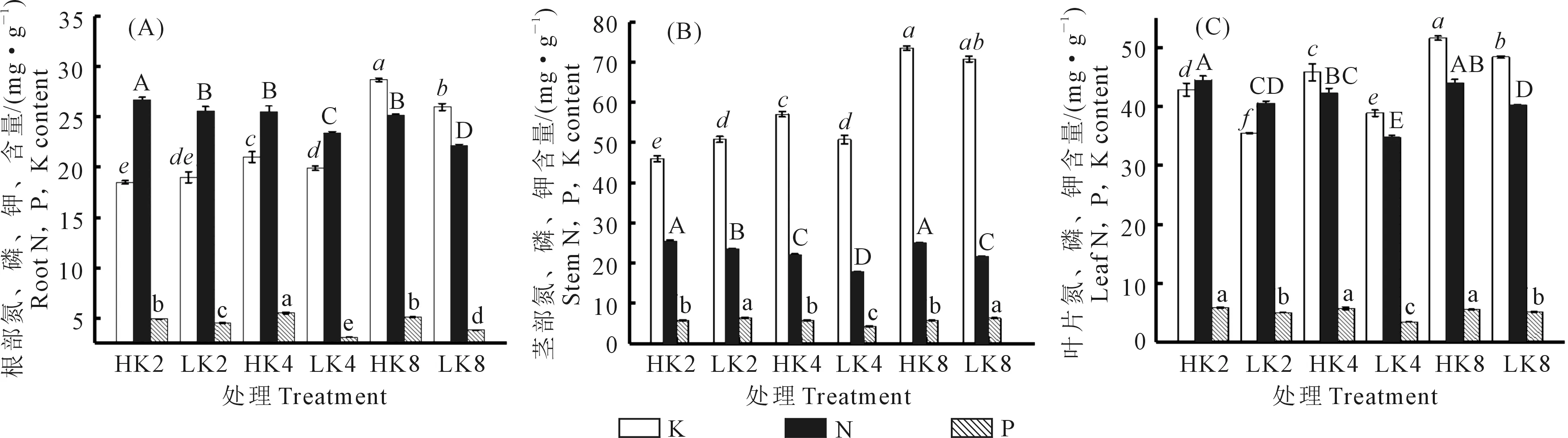
注:图中不同字母(大写字母代表氮,斜体字母代表钾,小写字母代表磷)表示处理间在0.05水平差异显著,下同。Note:Values followed by different letters in the figure (capital letters represent nitrogen, italic letters represent potassium, and lowercase letters represent phosphorus) indicate significantly different among different treatments at the 0.05 level. The same below.HK2: HVPD+K2, LK2: LVPD+K2, HK4: HVPD+K4, LK4: LVPD+K4, HK8: HVPD+K8, LK8: LVPD+K8.
2.3 VPD与钾素互作对番茄各器官氮、磷、钾积累量的影响
由图3可知,在高VPD环境下,植株各器官的钾、氮积累量会随着钾素水平的增加而增加;而在低VPD下变化规律不明显,其中叶片的钾、氮积累量随着钾素水平的增加呈现出先上升后下降的趋势,在中钾水平时出现最大值,钾、氮积累量分别为186.0、214.5 mg;而茎和根中的钾积累量呈现逐渐上升的趋势,氮积累量的变化趋势不明显;对于各器官磷积累量来讲,钾素水平的变化对其无显著影响。在不同钾素水平下,降低VPD会显著增加番茄各器官的氮、磷、钾积累量,其中在高钾水平时,植株总钾量与总磷量出现最大值,分别为382.4 mg和43.5 mg;在中钾水平时,植株总氮量出现最大值,为301.4 mg。
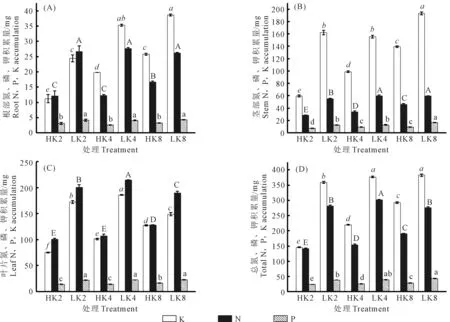
图3 VPD与钾素互作对番茄各器官氮、磷、钾积累量的影响
植株中钾素的分配以茎中最多,其次为叶,呈茎>叶>根的规律,这可能与钾参与茎部纤维素的合成和茎粗的增加有关,而氮素与磷素的分配则呈现出叶>茎>根的趋势,叶片中氮、磷积累的增多有利于光合作用的进行。
2.4 VPD与钾素互作对番茄气体交换参数的影响
由图4可以看出, VPD与钾素对于植株光合速率具有显著的交互作用。与高VPD相比,降低VPD使番茄叶片的净光合速率(Pn)、胞间CO2浓度(Ci)及气孔导度(Gs)显著升高,其中Pn与Gs在中钾时达最大值,Ci在高钾时达最大值。同时,高VPD显著增加了番茄幼苗的蒸腾速率(Tr),高钾时达最大值,低VPD下不同钾素水平对Tr无显著影响。与高VPD相比,低VPD下不同钾素水平(由低到高)的Pn分别提高了18.9%、32.6%和10.3%;Ci分别提高了24.1%、16.1%和17.2%;Gs分别提高了106.2%、92.5%和63.5%;Tr则分别降低了30.5%、30.6%和38.3%。在同一VPD下,钾素水平对Ci无显著影响。
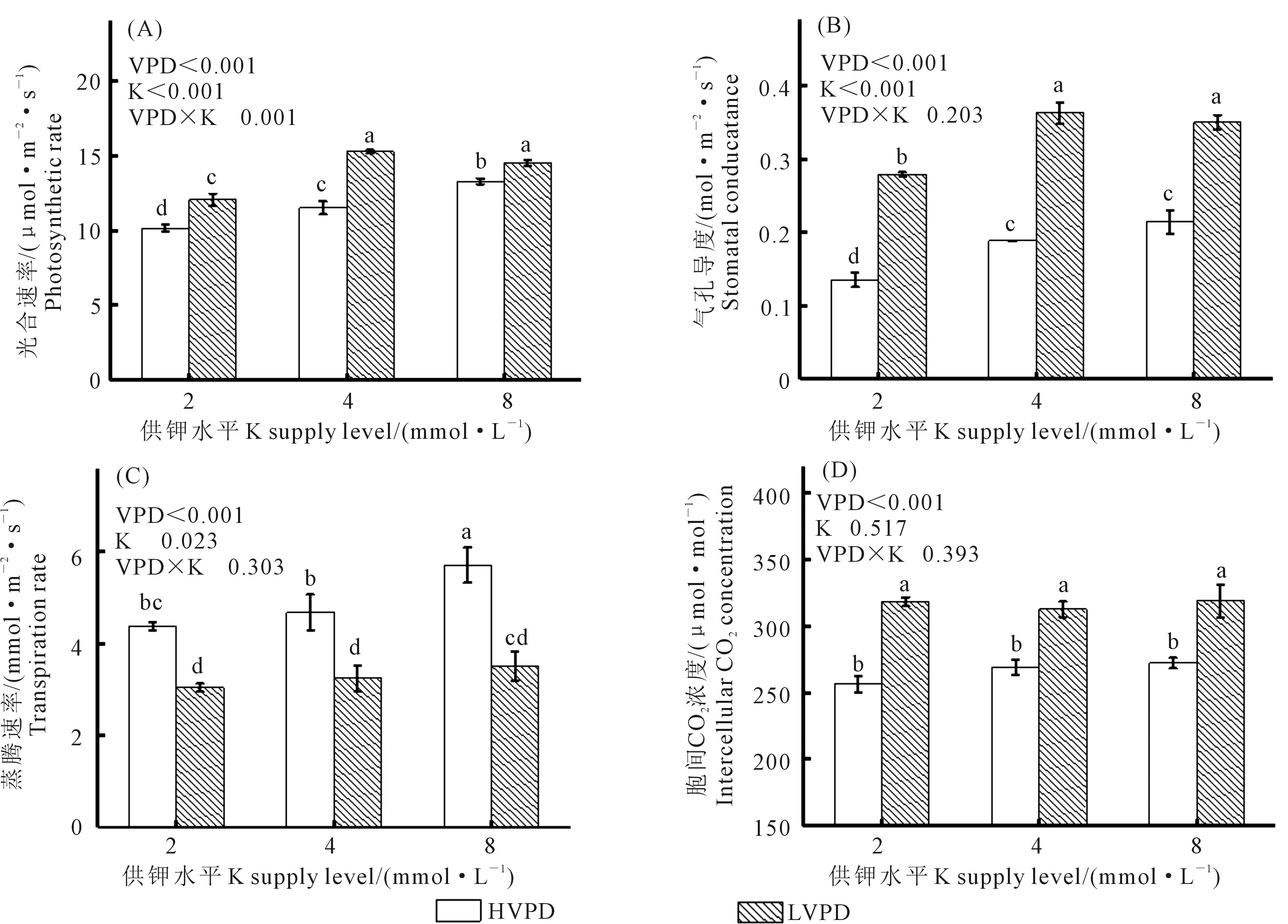
图4 VPD与钾素互作对番茄气体交换参数的影响
2.5 VPD与钾素互作对番茄CO2响应的影响
研究光合响应和CO2响应曲线可以更好地了解植物的光合生理特性,同时也反映了植物对环境的响应情况。在有效CO2浓度范围内,低VPD处理下各组的最大净光合速率(Pmax)、初始羧化效率(CE)均显著大于高VPD处理(表2),且均在中钾时有最大值;与高VPD相比,随着钾素水平的增加,低VPD下Pmax分别提高了18.3%、19.3%和9.4%,CE分别提高了37.2%、26.9%和12.3%。同时,降低VPD显著降低了番茄幼苗的光呼吸速率(Rp),且不同钾素水平对光呼吸速率没有产生显著影响。
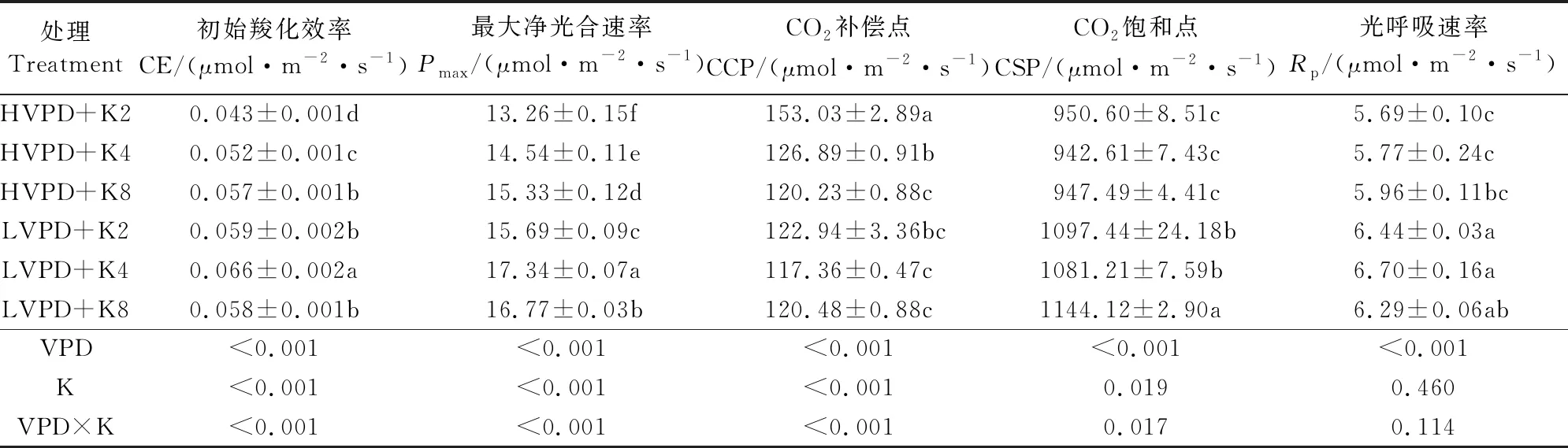
表2 VPD与钾素互作对番茄CO2响应曲线拟合参数的影响
植物进行净光合作用时可利用的CO2范围大小一般用CO2饱和点(CSP)与补偿点(CCP)之间的差值来表示。低VPD下低钾、中钾、高钾水平(K2、K4、K8)对应的进行净光合作用的CO2范围分别为974.51、1026.76 μmol·m-2·s-1和960.74 μmol·m-2·s-1;高VPD下对应的响应区间依次为797.57、815.72 μmol·m-2·s-1和827.27 μmol·m-2·s-1;在不同钾素水平下,降低VPD,各响应区间均增加。高VPD下,响应区间随着钾素水平的增加而增加,而低VPD下规律不明显。其中,低VPD下的中钾处理区间值最大,说明其对不同CO2浓度环境适应性最强。
2.6 VPD与钾素互作对番茄光合响应的影响
由表3可知,在有效光强范围内,低VPD处理下各组的最大净光合速率(Pmax)、表观量子效率(AQY)和暗呼吸速率(Rd)均显著大于高VPD处理,其中中钾水平时出现Pmax值,高钾时出现最大AQY值,低钾时出现最大Rd值;与高VPD处理相比,随着钾素水平的增加,低VPD下Pmax分别提高了36.6%、22.4%和8.8%,AQY分别提高了40.8%、19.7%和13.8%,Rd分别提高了23.9%、49.8%和46.4%;钾素水平越低,番茄幼苗的Rd越大,说明低钾胁迫提高了植株对光合产物的消耗。
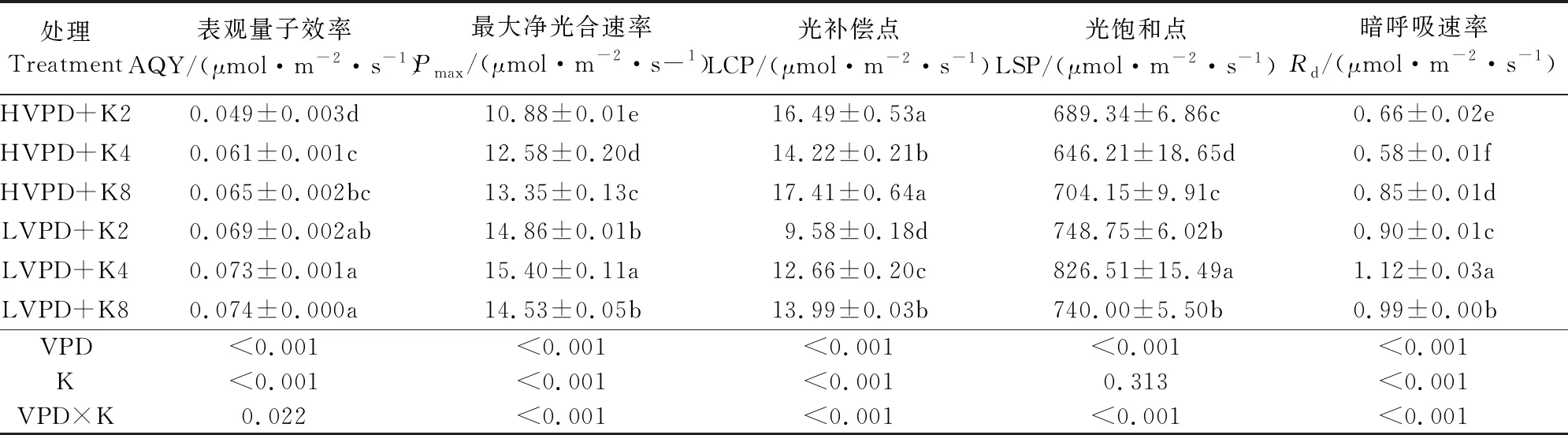
表3 VPD与钾素互作对番茄光合响应曲线拟合参数的影响
不同环境下植物对光照的需求是不一样的,这些差异主要是由植物叶片的光饱和点和光补偿点来决定的。低VPD下各钾素水平的光饱和点(LSP)均显著高于高VPD下相应处理,其中中钾处理的LSP显著高于其余处理。光补偿点(LCP)在不同处理间的差异性变化与光饱和点(LSP)的差异性变化没有内在一致性[11]。低VPD下低钾、中钾、高钾水平(K2、K4、K8)对应的光环境适应区间分别为732.84、774.54 μmol·m-2·s-1和726.01 μmol·m-2·s-1;高VPD下的响应区间依次为 676.71、636.46 μmol·m-2·s-1和688.37 μmol·m-2·s-1。结果表明,降低VPD增大了番茄幼苗对光环境的适应区间,其中,低VPD下的中钾处理区间值最大,说明其对不同光环境的适应性最强。
3 讨 论
3.1 VPD与钾素互作对番茄氮、磷、钾吸收的影响
植物生长受许多种环境因素的调节。引起植物生长公认的最重要环境因素之一是大气湿度。水分蒸发是由叶片组织与大气之间的水汽压差驱动的;在高湿条件下,植物会以相对较小的成本吸收水分中的CO2,但同时蒸腾作用的降低可能会削弱植物对矿物质养分的吸收[12]。本试验发现, 在各钾素水平下,低VPD对番茄叶片的氮、磷、钾含量,对根的钾、磷含量以及茎的氮含量均有显著的稀释效应;在中钾水平时, 低VPD对茎的钾、磷含量、根的氮含量也有明显的稀释效应(图2)。在低VPD下,植株体内氮、磷、钾元素含量的下降有不同的解释,一方面是与植株碳同化增加所引起的养分稀释效应有关[13];另一方面是在低VPD条件下,叶片与空气之间的湿度梯度降低,从而使茎液通量密度降低,这阻碍了可溶性矿物质在土壤中的大量流动和根系对养分的吸收[14]。
在本试验各供钾水平下,低VPD均显著增加了植株的总氮、磷、钾吸收量(图3),这可能归因于高湿环境下的植株普遍具有较高的生物量(图1A),且相对湿度的升高会增加不同植物吸收根系的表面积和根尖数量[15],促进了根系对养分的接触和获取,一定程度上弥补了由低VPD下蒸腾速率降低(图4C)导致的蒸腾驱动的质量流减少对养分吸收带来的负面影响。同时,大量研究表明氮、钾交互作用呈正相关关系,表现在钾促进氮的吸收和积累[16]。随着钾素水平的提高,植株氮素吸收水平也随之增加,适宜浓度的钾素水平更有利于氮从根部向地上部运输[11]。本试验也得出了相似的结果,即在中钾水平时,低VPD下的叶片氮积累量显著高于其他处理。
3.2 VPD与钾素互作对番茄光合生理的影响
光合作用是植株生物量积累的物质基础,影响植物的生长发育进程[17]。空气湿度是影响植物光合生理的关键环境条件之一,翁晓燕等[18]研究认为,空气湿度不仅能从气孔开度的闭合上影响叶片光合,还能通过改变细胞内Rubisco酶的活性直接影响光合速率。另外,钾素与Rubisco酶活性密切相关。前人研究表明,钾素胁迫下叶片Rubisco酶活性显著降低,这成为限制叶片光合速率的主要原因[19]。在CO2响应曲线中初始斜率(即羧化效率CE)与Rubisco含量及活性呈正相关,是反映CO2利用效率的重要指标[20]。本试验结果表明,高温下增加空气湿度显著增加了番茄幼苗的CE(表2),在中钾水平时达到最大值,且VPD×K对CE存在显著的交互作用,间接说明了VPD与钾素共同提高了Rubisco酶活性,进而促进了植株光合作用。
通过光合与CO2响应曲线研究植物对环境的响应一直是科学研究的热点问题。Pmax反映了植物叶片的最大光合能力,AQY是反映叶片对光能,尤其是弱光利用情况的重要指标,Rd是植物消耗光合产物的速率。本研究中,低VPD处理下的番茄叶片Pmax、AQY和Rd与高VPD处理相比均显著增加(表3),说明了低VPD处理下番茄幼苗吸收与转换光能的色素蛋白复合体较多,可以更有效地利用弱光[21]。同时,低VPD处理下番茄幼苗的LSP呈上升趋势,且LCP较低,这表明空气湿度的升高能够增强植株对外界环境的适应性,提高了番茄对光的适应范围[22]。CCP是植物利用CO2进行光合作用的临界浓度。低VPD处理下的植株CCP都比较低,CSP和表观羧化效率CE较高(表2),且具有较高的CO2浓度适用区间和高CO2利用率等光合特性,侧面说明了降低VPD可以缓解高温对植株光合的抑制。光呼吸可以作为保护光合作用的反应中心以免被强光所破坏的一个途径;光呼吸的改变是植物面对逆境时所做出的适应性反应。前人研究表明,水分胁迫下植株叶片气孔减小,叶肉细胞内CO2/O2比例发生变化,会导致光合作用下降的同时光呼吸增强[23]。本试验也得出了类似的结果,即高VPD下的番茄叶片均具有较高的光呼吸速率(表2)。
矿质营养元素与植物的光合作用密切相关,在不同水平的钾素供应下,植物的光合能力会发生显著改变,钾主要影响气孔开闭和相关酶活性,在农业生产中可以通过钾肥的合理施用来提高植物的光合能力和生产潜力[24]。本试验中,低VPD环境下Pn和Gs随着供钾水平增大呈先升后降的趋势,番茄叶片Pmax、AQY和CE的变化也响应了这一变化趋势,说明低VPD下适宜的钾素水平能够缓解高温对光合造成的不良影响,一定程度上提高了番茄的光合生产潜力,这一方面是由于正常的钾浓度可以促进氮素的吸收利用(图3),而叶片光合作用速率会随叶片氮素的增加而增加[25],氮素可以提高叶绿素的合成与相关酶活性[26];另一方面也与叶片水势变化带来的气孔导度提高有关 (图4B),钾素作为植物体内主要的渗透调节物质,在调节植物细胞水势的方面起着重要作用[8]。低VPD下中钾水平的叶片钾素积累量最多(图3C),随着钾素的增多,叶片水势也随之提高(图1B);此外,降低VPD可以通过抑制蒸腾驱动力和降低蒸腾速率(图4C)来缓解叶片脱水,有效地防止了叶片水势的下降[27]。叶片维持较高的水势以及保卫细胞膨压可以减少CO2进入气孔的阻力,维持植物气孔开放(即较高的Gs),进而改善了光合作用。而在高VPD环境下相关光合参数却随钾素水平的增加而增加,高钾水平下综合光合能力最强。不同VPD下最适宜施钾量有差异可能是由于植物组织中存在一个钾素的临界值[28],过量的钾素会抑制植株生长[29]。同时,本研究还发现,不论处于何种VPD环境,不同钾素水平均未对番茄幼苗的光呼吸速率(Rp)产生显著影响(表2),这表明番茄幼苗对钾胁迫具有一定的适应性和自我保护能力[11]。
4 结 论
1)与高VPD相比,在不同钾素水平下降低VPD均能显著增加番茄幼苗的干重、水势和对氮、磷、钾元素的积累量,其中在中钾水平下干重、水势和氮积累量达到最大值,分别为12.73 g、-0.74 MPa和301.4 mg。
2)低VPD环境下,番茄叶片的净光合速率(Pn)、胞间CO2浓度(Ci)及气孔导度(Gs)显著升高,其中Pn与Gs在中钾水平时达到最大值15.3 μmol·m-2·s-1和0.36 mol·m-2·s-1;降低VPD增加了植株对不同CO2浓度和光环境的适应区间,在中钾水平时区间值最大,分别为1 026.76 μmol·m-2·s-1和774.54 μmol·m-2·s-1。
3)高VPD高钾处理与低VPD中钾处理在各自的湿度环境内最有利于植株生长,对环境的综合适应能力最强;从资源节约的角度来看,高温环境下降低VPD一定程度上可以实现生产上钾肥的减量使用。
猜你喜欢
小猕猴学习画刊(2022年12期)2022-02-06 03:00:52
今日农业(2021年21期)2022-01-12 06:31:52
今日农业(2021年10期)2021-11-27 09:45:24
今日农业(2020年23期)2020-12-15 03:48:26
中国火炬(2015年12期)2015-07-31 17:38:35
植物营养与肥料学报(2014年1期)2014-03-11 20:24:36
中国火炬(2013年3期)2013-07-24 14:15:06
植物营养与肥料学报(2012年1期)2012-10-26 02:49:22
植物营养与肥料学报(2011年2期)2011-10-26 03:52:06
中国火炬(2011年8期)2011-07-25 10:44:25
