
引种美国红橡的纤维形态、 微纤丝角及结晶度
2020-01-18 02:38:32李聪聪黄利斌
浙江农林大学学报 2020年1期
李聪聪, 潘 彪, 王 慧, 黄利斌
(1. 南京林业大学 材料科学与工程学院, 江苏 南京210037; 2. 江苏省林业科学研究院, 江苏 南京
211153)
木材作为环境友好型的绿色原材料, 是国民经济发展的基础材料。 为坚持生态发展战略, 中国林业相关部门启动了一批重点生态工程, 在天然林保护的同时, 大力发展人工林种植。 随着国内民众生态意识的增强和国家森林保护力度的加大, 木材刚性需求增加, 木材的对外依赖度达到了近50%。 据测算,到2020 年, 中国木材年需求量将达到8 亿m3, 对大径材和珍贵树种用材需求也会大幅增加[1]。 引进国外优良树种是提高林业生产力、 填补国内名特优新经济林、 满足人们对林产品需求的有效途径, 对于在短期内实现有效供给、 缓解木材及林产品的供需矛盾, 实现可持续发展, 具有十分重要的意义。 中国土壤气候条件与北美相似, 引种美国红橡树种具有得天独厚的优势。 引种树种能在短时间内增加当地的植物种类, 树种的合理布局和应用, 对弥补优质用材资源不足有重要意义。 橡树又称栎树, 是壳斗科Fagaceae 栎属Quercus树种的统称, 用途广泛, 材质优异, 是优质商品材和燃料的重要来源[2]。 中国自1998 年开展北美栎树种质资源引种与栽培研究, 对引种试验、 引种树苗抗逆性[3]研究较多, 为栎类树种引进开发利用提供了理论基础。 纤维形态特征与木材生长状况以及强度、 密度以及物理、 力学性质有密切关系, 可用来进行木材的材性预测。 微纤丝角在一定程度上可以反映木材的物理和力学性质, 如木材的尺寸稳定性、 干缩湿胀及弹性模量、 强度和蠕变特性等, 是评价木材品质性能和良种选育的重要依据[4]。 木材的结晶度与木材密度、 强度、 尺寸稳定性存在着正相关关系[5], 研究结晶度有助于了解木材解剖结构与其性质之间联系。 本研究以纳塔栎Quercus nuttallii、 水栎Quercus nigra和舒玛栎Quercus shumardii为对象, 比较了3 种美国红橡纤维形态、 微纤丝角和结晶度的异同, 以期为引种栎木的科学利用提供理论依据, 为更大范围引种栽培提供理论基础。
1 材料与方法
1.1 实验材料
根据GB/T 1927-2009《木材物理力学试材采集方法》采集试样[6]。 选取14 年生纳塔栎、 水栎、 舒玛栎各4 株; 其中纳塔栎、 水栎采自江苏省林业科学研究院句容苗圃(年平均气温为15.2 ℃, 年降水量为1 058.8 mm)。 纳塔栎平均树高为8.48 m, 平均胸径为19.28 cm; 水栎平均树高为10.68 m, 平均胸径为15.38 cm; 舒玛栎采自于南京彩树种植有限公司江宁区种植基地(年平均气温为15.4 ℃, 年降水量为1 200.0 mm), 平均树高为11.70 m, 平均胸径为15.20 cm。
1.2 实验方法
1.2.1 木材纤维形态测定 将样木自胸高处圆盘锯解, 立即沿南北向自髓心向外逐年切取样块并依次编号, 由于第1, 2 年轮宽度较窄, 合并为1 个样品。 将各标号木块切成火柴棍大小, 每个年轮制4 组平行样。 用富兰克林离析法[7]离析细胞, 制成临时切片, 置于普通光学显微镜下; 随机测量50 个细胞长度、 宽度和腔径, 计算双壁厚、 壁腔比。 结果取平均数。
1.2.2 木材微纤丝角测定 将3 种样木按1.2.1 制成厚度约为2 mm 的试样, 采用Ultima IV 组合型多功能水平X-射线衍射仪(XRD)按Cave 0.6T 法[8]测量试材微纤丝角。 结果取平均数。
1.2.3 木材结晶度测定 将3 种样木按1.2.1 制成火柴棍大小试样, 磨成80~100 目木粉, 室温下压成薄片后测定木材结晶度, 同时分析木材2θ 衍射强度曲线。 计算结果取平均数。 木材纤维的衍射强度在2θ=22°是极大值, 其积分强度用I002表示; 在2θ=18°附近出现波谷, 是木材纤维中无定形区衍射的散射强度, 其积分强度用Iam表示。 相对结晶度(ICr)公式[9]如下:ICr=100×(I002-Iam)/I002。
1.2.4 数据处理 试验数据采用Excel 2013 进行方差分析及回归分析, Origin 软件作图。
2 结果与分析
2.1 纤维形态
由表1 可知: 纳塔栎纤维长度径向变化范围为1 032.46~1 255.46 μm, 平均值为(1 172.14±86.88)μm; 水栎纤维长度径向变化范围为1 018.04~1 281.27 μm, 平均值为(1 178.68±95.54) μm; 舒玛栎纤维长度径向变化范围为1 010.34~1 292.56 μm, 平均值为(1 162.45±112.67) μm。 其中, 舒玛栎的平均纤维长度最短, 纳塔栎和水栎之间差异较小, 三者栎木纤维长度种间不具有显著差异(P>0.05)。 按照国际木材解剖学家协会理事会发布的纤维长度分级标准(<0.9 mm 的属于短纤维, 0.9~1.6 mm 的属于中等纤维, >1.6 mm 的属于长纤维), 3 种栎木纤维均属于中等纤维, 大于62 年生蒙古栎Q. mongolica纤维平均长度(1 001.20 μm)[10], 与40 年生欧洲栓皮栎Q. suber(960.00~1 220.00 μm)[11]、 33 年生冬青栎Q. ilex(1 103.00 μm)及24 年生深红栎Q. coccifera纤维平均长度(1 116.00 μm)相近[12]。 由图1A可见: 3 种栎木纤维长度从髓心至树皮方向总体呈上升趋势, 但径向变化尚未趋于稳定, 说明树木仍处于幼龄期。
3 种栎木纤维宽度为14.14~18.51 μm, 舒玛栎纤维宽度最大(16.91 μm±1.20 μm), 水栎(16.56 μm±0.70 μm)和纳塔栎(15.86 μm±1.11 μm)略小, 变异系数均小于10%, 变异较小。 纤维宽度种间差异不显著(P>0.05)。 3 种栎木纤维宽度均小于40 年生欧洲栓皮栎(18.40~21.49 μm)。 由图1B 可知: 3 种栎木纤维宽度的径向变化均随生长轮增加而增加, 但存在着一定的波动。
纤维双壁厚是木材质量、 基本密度和强度的物质基础[13], 受生态环境条件和气候因子的影响较大[14]。研究发现: 水栎双壁厚最大(11.19 μm±0.54 μm), 纳塔栎(10.23 μm±0.91 μm)和舒玛栎(10.96 μm±1.03 μm)略小, 3 种栎木双壁厚种间具有显著差异(P<0.05)。 由图1C 可知: 随生长轮增加, 3 种栎木纤维双壁厚径向均呈增长趋势。 较其他2 种栎, 水栎更易受生态环境条件和气候因子影响, 因而纤维双壁厚变化波动也较大。
纤维壁腔比是纤维双壁厚与纤维胞腔直径的比值, 是衡量木材纤维强度的重要指标。 纤维壁腔比越小, 则木材密度越大, 材质越坚硬[15]。 木材纤维工业原料中通常要求纤维为中级长度(900.00~1 600.00 μm), 长宽比不低于35, 壁腔比小于1[16]。 本研究发现: 3 种栎木壁腔比为1.55~2.44, 水栎壁腔比平均值最大(2.10±0.23), 纳塔栎(1.85±0.26)和舒玛栎(1.91±0.16)略低, 纤维壁腔比种间具有显著差异(P<0.05)。 3 种栎木壁腔比均大于1, 未达到纤维工业原料要求。 从髓心向外, 3 种栎木壁腔比径向均略微上升, 在一定范围内存在波动(图1D)。

表1 纤维形态的比较分析Table 1 Comparative analysis of fiber morphology
2.2 微纤丝角
微纤丝角是影响木材强度的主要因素, 通常作为材质评定及早期良种选育的重要指标。 本研究发现: 纳塔栎微纤丝角径向变化范围为30.02°~38.73°, 平均值为33.79°。 水栎为26.12°~33.52°, 平均值为30.48°。 舒玛栎为37.65°~30.93°, 平均值为34.10°。 3 种栎木微纤丝角大小依次为舒玛栎(34.10°)、 纳塔栎(33.79°)和水栎(30.48°), 微纤丝角种间具有极显著差异(P<0.01)。 3 种栎木微纤丝角平均值为32.79°, 略大于采用细胞壁高分辨率显微照片测定的夏栎Q. robur(22°~31.6°)[17]的微纤丝角, 可能与栎木未达到成熟状态有关, 也可能是因为测量方法不同, XRD 法测量时采用的是细胞壁各壁层微纤丝角平均值。
从图2 可以看出: 3 种栎木微纤丝角随生长轮增加总体呈下降趋势。 原因可能是试材越靠近髓心,树木年龄越小, 形成层原始细胞产生的子细胞胞壁就越薄, 各层的平均微纤丝角就越大[18]。
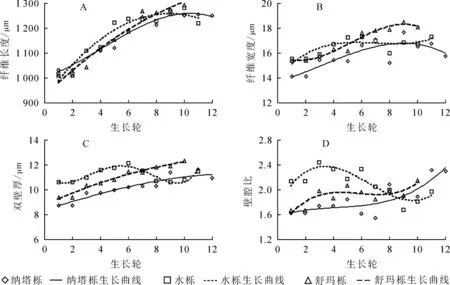
图1 纤维形态随生长轮的径向变化规律Figure 1 Radial variation of fiber morphology and growth ring from pith to bark
2.3 结晶度

图2 微纤丝角随生长轮的径向变化规律Figure 2 Radial variation of microfibril angle and growth ring from pith to bark

图3 结晶度随生长轮的径向变化规律Figure 3 Radial variation of the crystallinity and growth ring from pith to bark
纳塔栎结晶度径向变化范围为50.07%~52.52%, 平均值为51.35%。 水栎结晶度径向变化范围为51.45%~55.47%, 平均值为53.30%。 舒玛栎结晶度径向变化范围为51.23%~53.83%, 平均值为52.97%。结晶度大小依次为水栎(53.30%)、 舒玛栎(52.97%)和纳塔栎(51.35%), 均大于加拿大栎Q. canariensis(38.93%)[19], 3 种栎木种间具有极显著差异(P<0.01)。 纳塔栎、 水栎和舒玛栎结晶度的变异系数分别为1.63%、 2.80%和1.43%, 变异较小。 如图3 所示: 结晶度随生长轮增加总体呈上升趋势, 还未处于平稳状态, 可能与其处于生长时期有关。 结晶度径向变化趋势与其他学者研究一致: 在树木生长初期形成层细胞分裂速度较快, 结晶度较小, 随着树龄的增加, 结晶度也逐渐增大最后趋于稳定[20]。
2.4 预测模型
纤维形态、 微纤丝角与生长轮间关系密切。 对生长轮与纤维形态、 微纤丝角进行拟合分析, 建立适当的模型, 对人工林材质早期预测及林木的定向培育、 优化利用具有十分重要理论意义和应用价值。 以生长轮为自变量(x), 纤维长度、 纤维宽度、 纤维双壁厚、 微纤丝角分别为因变量(y), 进行回归拟合。
如表2 所示: 3 种引种栎木纤维长度与生长轮的关系与2 次曲线拟合度较好, 相关系数(R2)均在0.900 以上, 回归效果理想, 与大多数研究结果一致[21], 符合生物学生长规律。 3 种引种栎木纤维宽度与生长轮的3 次曲线拟合度高, 其中舒玛栎拟合效果最佳, 相关系数达到0.960。 纳塔栎和舒玛栎纤维双壁厚与生长轮的2 次曲线有较好的拟合关系, 水栎纤维双壁厚与生长轮的3 次曲线拟合关系较好, 其中舒玛栎拟合效果最佳, 相关系数达到0.972。 纳塔栎微纤丝角与生长轮的关系与对数函数有较好的拟合, 水栎及舒玛栎与2 次曲线有较好的拟合, 其中水栎拟合效果最佳, 相关系数为0.748。

表2 纤维形态、 微纤丝角与生长轮的回归分析Table 2 Regression analysis of fiber morphology, microfibril angle, and growth ring
3 结论
对纳塔栎、 水栎和舒玛栎纤维形态测量得知: 3 种引种栎木纤维长度为1 010.34~1 292.56 μm, 纤维宽度为为14.14~18.51 μm, 纤维长度、 宽度种间不具有显著差异(P>0.05), 其径向变化均为从髓心至树皮方向呈上升趋势, 但径向变化尚未趋于稳定; 3 种栎木纤维双壁厚为8.77~12.34 μm, 壁腔比为1.55~2.44, 纤维双壁厚、 壁腔比种间具有显著差异(P<0.05), 其径向变化均为从髓心至树皮方向略微上升, 在一定范围内存在波动。
3 种栎木中舒玛栎微纤丝角最大(34.10°), 纳塔栎(33.79°)和水栎(30.48°)略小, 微纤丝角种间具有极显著差异(P<0.01)。 径向变化为微纤丝角随着生长轮增加而降低; 结晶度大小依次为水栎(53.30%)、 舒玛栎(52.97%)和纳塔栎(51.35%), 结晶度种间具有极显著差异(P<0.01), 径向变化为结晶度随生长轮增加总体呈平缓上升趋势。
对生长轮与各指标进行拟合分析发现: 3 种栎木纤维长度与生长轮关系用2 次曲线拟合效果理想,相关系数均在0.900 以上。 纤维宽度与生长轮关系: 舒玛栎用3 次曲线拟合效果理想, 相关系数在0.900 以上。 双壁厚与生长轮的关系: 舒玛栎用2 次曲线拟合效果理想, 相关系数在0.900 以上。 微纤丝角与生长轮的关系: 水栎用2 次曲线拟合效果较好, 相关系数在0.700 以上。
由此认为: 14 年生引种美国红橡仍处于幼龄期, 纤维形态、 微纤丝角与结晶度径向变化趋势尚未趋于稳定。
猜你喜欢
建筑与预算(2024年2期)2024-03-22 06:51:36
大自然探索(2024年1期)2024-02-29 09:10:32
东坡赤壁诗词(2023年1期)2023-05-30 12:52:04
军事文摘(2021年16期)2021-11-05 08:49:06
西北林学院学报(2021年2期)2021-04-08 00:28:52
浙江农林大学学报(2019年2期)2019-03-25 12:37:40
中国科技博览(2017年39期)2017-09-07 09:14:31
核技术(2016年4期)2016-08-22 09:05:24
塑料制造(2016年5期)2016-06-15 20:27:39
中国质量与标准导报(2014年5期)2014-02-28 22:23:22
