
固沙樟子松种子萌发与幼苗生长对干旱胁迫的响应及抗旱性评价
2020-01-18 02:16王东丽张日升吴叶礼秦崧悦龙丹丹沈海鸥
浙江农林大学学报 2020年1期
王东丽, 张日升, 方 祥, 王 凯, 吴叶礼, 秦崧悦, 龙丹丹, 沈海鸥
(1. 辽宁工程技术大学 环境科学与工程学院, 辽宁 阜新123000; 2. 辽宁省沙地治理与利用研究所 章古台科尔沁沙地生态系统国家定位观测研究站, 辽宁 阜新123000; 3. 吉林农业大学 资源与环境学院, 吉林 长春130021)
樟子松Pinus sylvestrisvar.mongolica以其抗逆性强(耐旱性、 耐寒性)、 速生、 成材早的生物学特性, 已成为中国北方干旱半干旱地区山地、 草原和沙地植被恢复的主要造林树种之一, 发挥着重要的水土保持与防风固沙等生态效益[1-2]。 然而, 不同引种区的樟子松林在非衰老期开始出现生长衰退、 病虫害, 甚至死亡等现象[3-5]。 同时, 中国北方最早引种的樟子松已进入更新阶段, 天然更新是森林经营可持续发展的重要方式。 樟子松在天然分布区具有很强的天然更新能力, 但在大部分引种区却存在天然更新困难的问题[1,6-8]。 可是, 大面积的樟子松造林仍在进行[9], 其可持续发展问题亟需解决, 天然更新引起相关学者的广泛关注。 植物更新的关键敏感期在于种子萌发与幼苗生长[10]。 以往研究发现, 极端温度、 蒸发量和多年平均积雪日数等气候因子[11]与林分郁闭度、 枯枝落叶层、 植被盖度和立地条件等环境因子[7]是引种区樟子松天然更新的主要障碍, 朱教君等[12]研究指出: 干旱胁迫可能通过影响引种区沙地樟子松种子萌发而限制其天然更新。 然而, 不同生长发育阶段林木结实特性、 种子质量各异[13], 引种区不同生命周期阶段的樟子松固沙林种子萌发与幼苗生长特性是否存在差异, 对干旱胁迫的响应及抗旱性是否存在规律性, 是否在种子萌发期具备天然更新的优势, 这些问题亟需明确。 本研究以沙地引种区不同林龄樟子松种子为材料, 研究樟子松种子萌发与幼苗生长对干旱胁迫的响应, 采用隶属函数法综合评价其抗旱性, 明确不同生命周期樟子松固沙林在种子萌发期的天然更新优势, 为其经营管理与可持续发展提供指导, 进而保障其发挥生态防护功能。
1 材料与方法
1.1 供试材料
供试樟子松种子于2017 年9 月采集于辽宁省章古台地区, 选取4 个不同林龄的典型结实林分, 分别随机选择10 株结实株, 从树冠上部、 中部、 下部采集种子。 具体情况见表1。
1.2 干旱胁迫实验
采用聚乙二醇(PEG 6000)溶液模拟不同干旱胁迫处理, 设置质量分数0(ck)、 5%(T1)、 10%(T2)、15%(T3)、 20%(T4)等5 个胁迫梯度, 模拟水势梯度分别为0、 -0.054、 -0.177、 -0.393 和-0.735 MPa[14],每个处理3 次重复。 以铺有2 层滤纸的培养皿(直径9 cm)为培养床, 选取大小均一、 成熟饱满的种子置床, 采用100 ℃热水消毒杀菌, 每个重复放置50 粒种子, 置于人工气候培养箱。 依据研究区的多年平均气象观测资料, 培养条件设置为白天(光照时段)13 h, 温度25 ℃, 夜晚(黑暗时段)11 h, 温度为16 ℃, 光合光子照度为110 μmol·m-2·s-1, 相对湿度为60%。
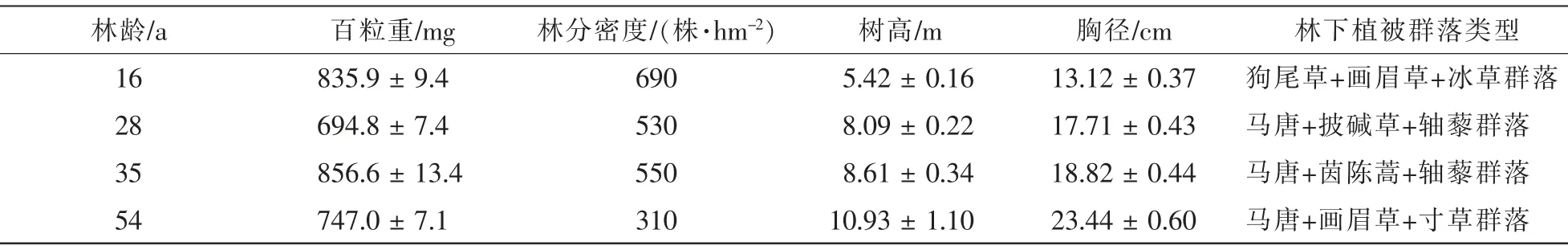
表1 种子及采集地的基本概况Table 1 Overview of the seed and its collection sites
1.3 种子萌发指标测定
从种子置床起, 隔24 h 记录1 次种子萌发数, 种子萌发以胚突破种皮且长为种子长度的一半时为标准(参考《国际种子发芽规程》), 记数, 直到连续5 d 不出现有萌发时为止。 萌发特性用以下指标来表征: ①萌发率=种子萌发总数/供试种子数×100%。 ②萌发时滞: 从种子置床到种子开始萌发持续的时间(d)。 ③萌发历时: 从第1 粒种子开始萌发至种子萌发结束所持续的时间(d)。 ④t50和t90: 从种子萌发开始, 种子萌发数分别达到萌发总数的50%和90%所持续的时间(d)。 其中, 萌发率表征种子萌发能力的大小, 萌发时滞和萌发历时表征种子萌发格局,t50和t90表征种子萌发速率。
1.4 幼苗生长指标测定
随机从各处理选取20 粒新萌发种子, 置于新的培养皿继续进行相应处理的培养。 待培养15 d 后,随机选择10 株幼苗, 在滤纸上吸干幼苗表面的水分后, 测定其幼苗鲜质量、 胚芽长、 胚根长以及胚轴直径。 为减少种子百粒重差异导致幼苗鲜质量的不准确性, 本研究采用幼苗生长相对值来表征, 即幼苗鲜质量与种子质量的比值。
1.5 抗旱性综合评价方法
采用模糊数学隶属函数法对不同林龄樟子松种子更新关键期抗旱性进行综合评价。 计算公式如下:

式(1)中: X(u)为隶属函数值;X为参试值某一抗旱指标的测定值;Xmax和Xmin分别为指标的最大值和最小值。 如果某一指标与抗逆性成负相关, 则用反隶属函数计算。 公式为:

先运用式(1)或式(2)求出不同林龄樟子松种子在不同胁迫下的隶属函数值, 将各林龄樟子松抗旱指标的隶属函数值加权平均。 通过比较加权平均值的大小, 确定不同林龄樟子松更新关键期抗旱性的强弱[14]。
1.6 数据处理与分析
数据统计与分析采用SPSS 18.0 软件, 采用最小显著差法(LSD)比较同一林龄下不同干旱胁迫对樟子松种子萌发率与幼苗生长特征影响的差异性, 采用双因素方差分析法分析不同林龄与不同干旱胁迫对樟子松种子萌发特征与幼苗生长特征的影响及二者之间的互作作用。 采用主成分分析法计算各指标的权重。 结果图绘制采用SigmaPlot 13.0 软件。
2 结果与分析
2.1 樟子松对干旱胁迫的种子萌发响应特征
2.1.1 种子萌发率 由表2 可见: 樟子松种子萌发率对干旱胁迫表现出极显著的响应(P<0.001), 总体表现出在干旱胁迫下呈下降趋势, 变化在0~78.0%(对照为88.3%~99.3%), 仅林龄为28 年生的樟子松种子在T1处理下种子萌发率(94.0%)高于ck(图1)。 樟子松种子萌发率对林龄、 林龄与干旱胁迫交互作用均表现出显著的响应。 在干旱胁迫下, 28 年生樟子松种子萌发率整体表现为较高, 在T4处理时仍可达24.7%, 且随着干旱胁迫的增加逐渐下降; 而其他林龄种子萌发率对干旱胁迫的响应具有差异性, 16年生樟子松种子萌发率在T4处理下显著降低(P<0.05), 而35 和54 年生樟子松种子萌发率分别在T2和T3处理下已显著下降(P<0.05)。 可见, 樟子松种子萌发率对干旱胁迫的响应存在阈值, 且不同林龄的响应阈值各异, 林龄相对较小的樟子松种子萌发率响应干旱胁迫的阈值较大。
2.1.2 种子萌发格局 樟子松种子萌发时滞对干旱胁迫表现出显著的响应, 而对林龄、 林龄与干旱胁迫交互作用的响应不显著; 萌发历时对干旱胁迫与林龄表现出显著的响应, 而对林龄与干旱胁迫交互作用的响应不显著(表2)。 不同林龄樟子松种子萌发时滞在ck 处理下均为5 d, 在各干旱胁迫处理下总体变长(图2), 除35 年生外, 樟子松种子萌发时滞在T4处理下仅为2.7 d; 在T3和T4处理下不同林龄间种子萌发时滞变化较长, 特别是16 年生的樟子松种子萌发时滞在T4处理下达9.0 d。 与ck 相比, 樟子松种子萌发历时在T4处理下显著缩短, 但对其他干旱胁迫处理的响应存在林龄间差异, 16 和28 年生樟子松种子萌发历时均有所延长, 分别为8.0~10.33 和7.7~11.0 d, 而35 和54 年生樟子松种子萌发历时均缩短, 分别为5.7~9.0 和4.3~7.0 d。 可见, 种子萌发对干旱胁迫响应为启动推迟, 林龄较小的种子萌发持续时间延长, 而林龄较大的则反之, 而且随着干旱胁迫的增强萌发格局具有不稳定性。
2.1.3 种子萌发速率 樟子松t50和t90对干旱胁迫均表现出显著的响应, 而对林龄的响应不显著,t50对林龄与干旱胁迫交互作用的响应不显著,t90则显著(表2)。 由图3 可知: 樟子松种子t50和t90总体表现为随着干旱胁迫的增强先增大再减小, 在T3时达到最大值, 但在T4处理时有所下降, 主要由于强干旱胁迫导致林龄大的少量种子短时间内完成萌发, 相对而言萌发速率变快。 可见, 樟子松种子对干旱胁迫响应为萌发变慢, 且干旱胁迫越强萌发越慢。
2.2 樟子松对干旱胁迫的幼苗生长响应特征

表2 林龄和干旱胁迫对樟子松种子萌发影响的双因素方差分析

图1 不同处理下种子萌发率Figure 1 Seed germination percentages of different treatments

图2 不同处理下种子萌发格局Figure 2 Seed germination patterns under different treatments
经过15 d 的培养, T4处理下的樟子松幼苗均未生长, 幼苗鲜质量对林龄与干旱胁迫响应显著, 但对两者的交互作用响应不显著(表3)。 如图4 所示: 与ck 比较(变化在5.1~5.5), 樟子松幼苗生长相对值在干旱胁迫条件下均有所下降(变化在2.4~4.8); 除35 年生的樟子松幼苗生长相对值随着干旱胁迫的增加而降低外, 其他林龄的均随干旱胁迫的增强呈现先增大后降低的趋势。
由表3 可知: 除胚根长与胚轴直径对林龄的响应不显著、 幼苗鲜质量对林龄与干旱胁迫的互作响应不显著外, 幼苗胚芽长、 胚根长和胚轴直径对不同处理均显著响应。 由表4 可知: 与ck 比较, 樟子松幼苗胚芽长在干旱胁迫处理下均有所下降; 35 年生樟子松幼苗胚芽长随着干旱胁迫的增强逐渐减小, 其他林龄的则在T2处理下大于在其他干旱处理; 54 年生樟子松幼苗胚芽生长显著小于其他林龄。 樟子松幼苗胚根生长在干旱胁迫处理下整体表现为有所增加, 且随着干旱胁迫强度的加强先增大后降低, 尤其在T2处理下达到最长; 且林龄越小的樟子松胚根长在干旱胁迫处理下变化幅度相对较大。 樟子松幼苗胚轴直径生长在干旱胁迫下均有所下降, 且整体表现为随着干旱胁迫的加强而减小的趋势。 可见, 樟子松幼苗对干旱胁迫响应为胚芽与胚轴直径生长受限制, 而胚根生长有所促进, 且林龄小的响应更敏感。

图3 不同处理间种子萌发速率Figure 3 Seed germination rate of different treatments
2.3 不同林龄樟子松更新关键期抗旱性的综合评价
采用隶属函数法对樟子松种子萌发率、 萌发时滞、 幼苗鲜质量、 胚芽长、 胚根长、 胚轴直径进行综合评价, 结合主成分分析法得到了不同林龄樟子松的隶属函数的加权平均值, 如表5 所示: 抗旱性从大到小依次为16、 28、 35、54 年生, 即随着林龄的增加逐渐降低。
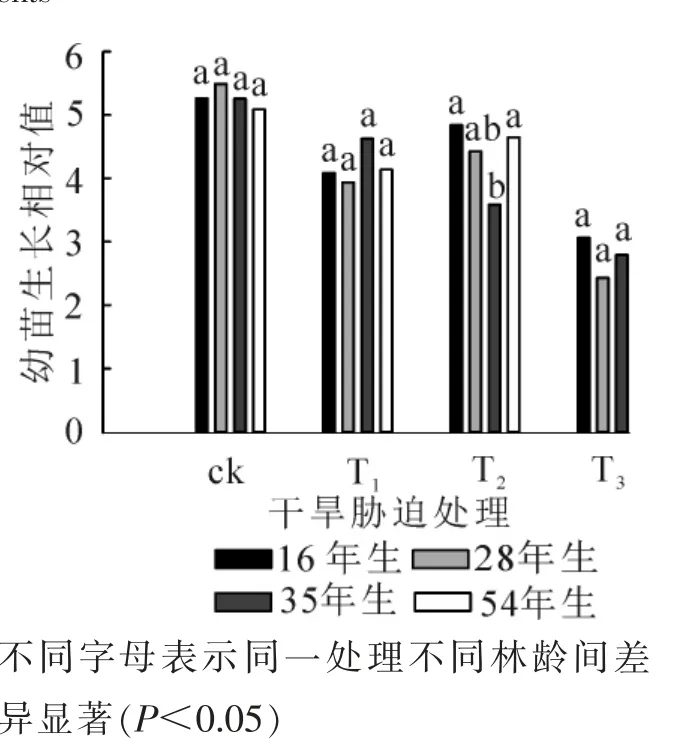
图4 不同处理下幼苗生长相对值Figure 4 Seedling growth relative value under different treatments

表3 林龄和干旱胁迫对樟子松幼苗生长影响的双因素方差分析Table 3 Two-way ANOVA analysis for the effects of forest age and drought stress on seedling growth for P. sylwestris var. mongolica
3 讨论
水分是植物种子萌发和幼苗生长、 存活的关键限制因子[15-16], 干旱胁迫是植物生长的逆境之一, 对植物的影响一直受到广泛关注[17]。 干旱胁迫下种子萌发、 幼苗生长及吸水特性对幼苗生存具有重要的意义[18]。 在脆弱生态系统中, 种子能够萌发成苗是植物在极端条件下生长发育的前提, 此阶段对干旱的适应性对于植物更新至关重要[10]。 大量研究利用PEG 6000 模拟干旱胁迫研究其对植物的影响及抗旱性,被证实是可靠的[15,19-20]。 本研究发现: 4 种林龄樟子松种子萌发率、 幼苗相对生长值、 胚芽长总体呈现随着干旱胁迫的加强而下降的趋势, 种子萌发时滞、 萌发历时、t50、t90、 胚根长与胚轴直径则呈相反的趋势, 因此, 引种樟子松种子萌发在干旱胁迫下总体为抑制性, 降低种子萌发率, 推迟种子启动萌发,延长种子萌发历时, 降低种子萌发速率, 这与以往对其他种子的研究结论一致[21]; 但本研究28 年生樟子松种子萌发率对干旱胁迫响应为低促高抑, 这与不少植物种子萌发对干旱胁迫的响应一致[15,19], 这种轻微的逆境胁迫能够激发植物种子自身保护机制, 减少种子萌发过程中膜系统的损伤, 进而提高其体内抗氧化物酶活性和可溶性蛋白质含量, 增强植物抗旱性, 提高植物种子的发芽率[20]。 此阶段的引种樟子松处于壮年期, 结实旺盛, 可为林分更新提供大量种子, 可利用轻度干旱胁迫处理在森林培育过程中进行引发。 植物在幼苗期对干旱胁迫反应最为敏感, 本研究不同林龄樟子松幼苗生长相对量、 胚芽与胚轴生长在干旱处理下均低于对照, 表现为引种樟子松幼苗生长受干旱胁迫限制。 种子发芽后能否有效定植成活主要取决于立地土壤水分状况和幼苗根系获取水分的能力[18]。 在干旱胁迫下, 胚根的生长对幼苗的生存、 生长和发育有着十分重要的意义[22]。 一定干旱胁迫下, 植物根系对土壤水分的减少最敏感, 胚根的伸长有利于从不同的土层深度获得更多水分, 这是植物适应干旱胁迫的一种生态适应表现[23]。 本研究樟子松幼苗胚根生长在干旱胁迫处理下整体表现为有所增加, 且随着干旱胁迫强度的加强先增大后降低, 可见引种樟子松幼苗具有抵抗一定干旱胁迫的能力, 这与何凌仙子等[24]对沙地赤松Pinus densiflora、樟子松和青海云杉Picea crassifolia的研究结果一致。
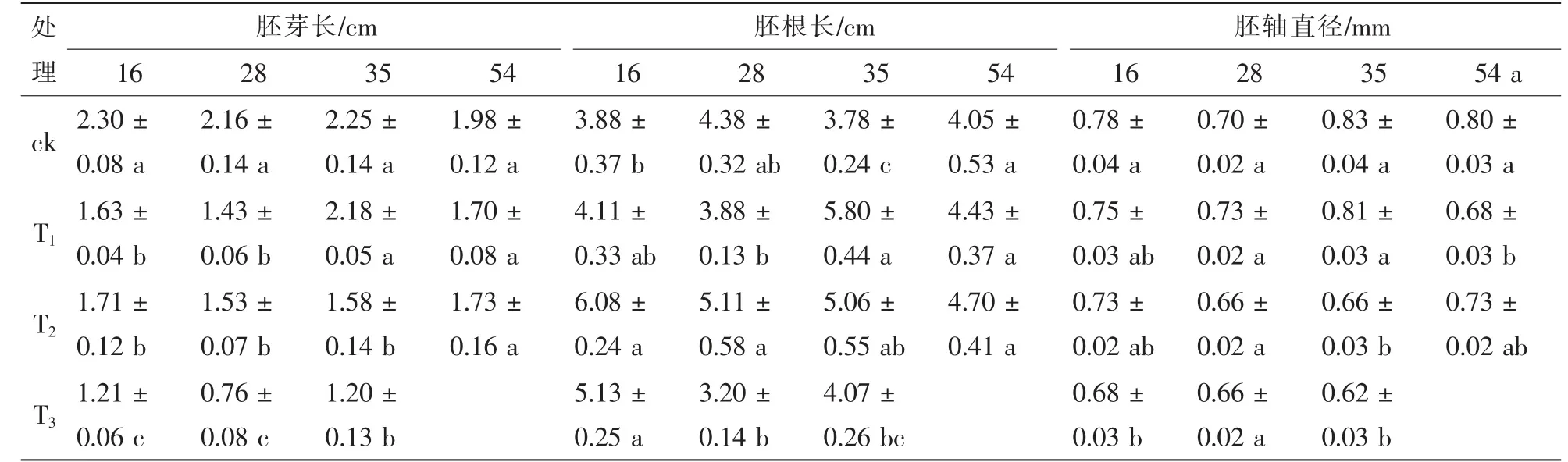
表4 不同处理下幼苗生长特征Table 4 Seedling growth characteristics of different treatments

表5 不同处理下樟子松种子萌发与幼苗生长指标隶属函数值及综合评价Table 5 Subordinate function values and comprehensive evaluation of seed germination and seedling growth for P. sylwestris var.mongolica under different treatments
植物的抗逆性是一个复杂的现象, 一般随个体的发育阶段而变化, 在逆境中的定植取决于萌发与幼苗阶段[25]。 种子的萌发和幼苗特性与其自身特性密切相关[26-27]。 林木不同生命周期阶段的结实特性各异, 如在青年期结实少而大, 壮年期结实多而小[28]。 本研究发现: 不同林龄樟子松种子萌发对干旱胁迫响应存在差异, 林龄相对较小的樟子松种子萌发率响应干旱胁迫的阈值较大, 不同林龄樟子松种子萌发率的抗旱性阈值有待深入研究, 进而为不同林龄樟子松人工促进天然更新的林分结构调控与立地管理提供科学依据。 潘昕等[27]研究表明: 种子的相对萌发率越大, 植物抗旱性越强。 由此可推断, 引种樟子松种子萌发在青壮年期具有较强的抗旱性。 本研究隶属函数法综合评价结果也证实这一结论。 林龄较小的种子萌发历时延长, 林龄大的种子仅有少数在强干旱胁迫下萌发持续时间短, 故种子萌发格局对干旱胁迫的响应存在林龄差异, 体现了不同生育期樟子松种子萌发响应干旱胁迫的适应策略不同。 樟子松种子对干旱胁迫响应总体为萌发变慢, 且随干旱胁迫增强萌发越慢。 干旱胁迫会使植株生长减慢甚至停止,干旱逆境下植物生长量的大小是衡量其抗旱能力的重要指标[29]。 本研究通过隶属函数法综合分析结果表明: 28 年生樟子松在种子萌发期具有较好的抗旱性, 但其幼苗各生长指标在干旱胁迫条件下较其他林龄小, 可能与其种子质量小有关。 54 年生樟子松在T4处理下幼苗完全不生长, 各幼苗生长指标在其他干旱胁迫处理下变化幅度也较小, 可见其对干旱胁迫的响应不敏感, 尤其是其胚根长随干旱胁迫的增强而增长较小, 而胚根长直接影响植物获取水分的能力, 再次表明林龄大的樟子松幼苗抗旱性弱; 相对而言, 16 年生樟子松的幼苗生长对干旱胁迫的响应更敏感。 本研究16 和35 年生樟子松的胚根长在各干旱胁迫处理下较其他林龄均变化较大, 表现出较强的抗旱性。
目前, 隶属函数法已被广泛应用植物抗旱性的综合评价, 特别是使植物种子萌发期抗旱性差异具有可比性、 可行性和可靠性[30]。 本研究采用隶属函数法综合分析, 不同林龄樟子松在种子萌发更新期抗旱性由强到弱依次为16、 28 、 35、 54 年生。 可见, 处于青壮年期的樟子松在更新敏感期的抗旱性更强。研究表明: 科尔沁沙地樟子松天然更新是可能的[31], 降水条件好的雨季, 在母树周围可见1 年生天然更新苗, 当年11 月仅有母树东北方向的幼苗能够存活, 但越冬后也会因寒干害死亡[7]。 可见, 樟子松种苗阶段的抗性对其更新成功的重要性。 由于引种区存在较多樟子松天然更新障碍的因子[7], 故在樟子松引种区宜进行人工促进天然更新。 因此, 对于林龄相对较小的更新林分, 创造有利于种子萌发出苗与幼苗存活、 建植的微生境, 保障其天然更新; 对林龄较大的衰退林分可优先选取抗逆性强的种子进行补播, 即选用青壮年樟子松种子, 进而提高人工促进天然更新的成功率。 然而, 植物的抗旱性是一种复合性状, 受形态、 解剖和生理生化等特性控制, 本研究仅限于种子萌发期抗旱性在萌发与形态方面的探究, 若要深入地掌握其抗旱性的机制仍需从解剖和生理生化方面开展进一步研究。
4 结论
干旱胁迫成为引种樟子松天然更新困难的重要障碍因子, 引种樟子松种子可通过降低萌发率, 推迟萌发, 延长萌发持续时间, 减缓萌发速率, 增加胚根生长适应干旱胁迫。 随着樟子松生命周期的演进,樟子松种子萌发期抗旱性逐渐减弱。 在人工促进引种樟子松天然更新时, 在林龄较小的更新林分, 创造有利于天然更新的微生境; 在林龄较大的衰退林分, 选用青壮年樟子松种子进行人工补播。
猜你喜欢
作物研究(2022年4期)2022-09-21
防护林科技(2022年5期)2022-08-22
安徽农业科学(2021年14期)2021-09-03
山西农业科学(2021年8期)2021-08-17
中国糖料(2021年3期)2021-07-13
辽宁林业科技(2021年3期)2021-07-08
防护林科技(2020年10期)2020-12-22
种子(2020年11期)2020-12-05
四川林业科技(2020年1期)2020-08-31
山西农业科学(2020年1期)2020-02-27
