
凡纳滨对虾对饲料中钙、磷的营养需求
2020-01-17 04:58安文强黎文伟谭北平杨奇慧迟淑艳董晓慧刘泓宇杨原志张海涛
水产科学 2020年1期
安文强,黎文伟,谭北平,杨奇慧,迟淑艳,董晓慧,刘泓宇,章 双,杨原志,张海涛
( 1.广东海洋大学 水产学院,水产动物营养与饲料实验室,广东 湛江 524088; 2.农业部华南水产与畜禽饲料重点实验室,广东 湛江 524000 )
凡纳滨对虾(Litopenaeusvannamei)又称南美白对虾,是世界三大主要养殖虾类之一,生长迅速,肉质鲜美,对环境变化耐受力强,在世界各地广泛养殖[1]。钙是甲壳动物的重要组成成分,除了构成体壳,还参与肌肉收缩、血液凝固、神经传递、调节渗透压、激活酚氧化酶原系统以及维持细胞膜的完整性和通透性[2-3]。磷是动物细胞核酸及细胞膜的重要成分,同时又直接影响所有细胞的能量反应,对动物的生长、骨骼的矿化有极大的作用,缺乏磷会抑制对虾的生长[4]。而饲料中钙、磷的水平过高会导致对虾向周围水环境的矿物排泄量增加,污染水体。有研究建议,将饲料中钙、磷比值当成一个独立的矿物质水平的指标[5],在满足对虾的生长需求时应注意严格控制钙、磷的添加量。
目前有关对虾钙、磷需求量已有研究。Deshimaru等[6]发现,在饲料中无需添加钙,仅需添加2%的磷即可满足日本囊对虾(Marsupenaeusjaponicus)的生长需求;而Kanazawa等[7]却发现,满足日本囊对虾生长的饲料磷添加量应为1%。中国明对虾(Fenneropenaeuschinensis)摄食含磷0.91%、钙2.2%的饲料时生长最佳[8]。海水养殖的凡纳滨对虾的基础饲料中不添加钙只添加0.35%磷即可维持生长和存活;若添加1%和2%的钙,则需分别添加0.5%~1%和1%~2%的磷才能维持对虾的正常生长[9]。低盐水体养殖的凡纳滨对虾摄食添加2%钙的饲料时,生长受抑制[10]。斑节对虾(Penaeusmonodon)饲料中钙含量为1.25%时,添加1%~1.5%的磷生长最佳[11]。饲料中钙含量为1.25%、磷含量为1.32%时印度明对虾(F.indicus)生长最佳[12]。但同时探查凡纳滨对虾商品饲料中钙、磷适宜添加量的研究尚未见报道。笔者研究在商品饲料中添加不同水平钙、磷对凡纳滨对虾生长性能、体成分、组织钙及磷的含量、血清生化指标及免疫酶活性等指标的影响,以确定凡纳滨对虾饲料中适宜的钙、磷含量,为生产凡纳滨对虾高效配合饲料提供参考。
1 材料与方法
1.1 材料
养殖试验在广东省湛江市东海岛广东海洋大学海洋生物研究基地室内海水养殖系统中进行。凡纳滨对虾购于湛江市东海岛中联虾苗厂。
试验饲料以红鱼粉、虾壳粉、豆粕、花生粕和玉米蛋白粉为蛋白源,鱼油、豆油和大豆卵磷脂为脂肪源,面粉为糖源,基础饲料配方见表1。
分别以乳酸钙、磷酸二氢钠(国药集团化学试剂有限公司)为钙源和磷源,在钙添加水平为0、0.5%、1.0%条件下,分别添加0、0.4%、0.8%、1.2%、1.6%的磷,配制15种试验饲料。饲料原料粉碎过80目筛,按配方准确称取,逐级扩大法混合,第一次混合15 min后,加油和卵磷脂,然后混匀,过80目筛,加入蒸馏水(30%,V/m),再混合均匀,压制成直径分别为1.0 mm和1.5 mm的颗粒饲料,60 ℃熟化30 min,风干后编号分装,-20 ℃冰箱中保存备用。

表1 基础饲料组成及营养水平(风干基础) %Tab.1 Ingredients and nutrient levels of the basal diet (air-dry basis)
注:1.每千克维生素预混料含有Vitamin premix contains the following per kg:维生素A醋酸酯retinyl acetate 10.00 g,维生素D3VD350.00 g,维生素E VE 99.00 g,维生素K VK 5.00 g,维生素B1VB125.50 g,维生素B2VB225.00 g,维生素B6VB650.00 g,维生素B12VB120.10 g,泛酸钙 calcium pantothenate 61.00 g,烟酸nicotinic acid 101.00 g,生物素biotin 25.00 g,肌醇inositol 153.06 g,叶酸folic acid 6.25 g,纤维素cellulose 389.09 g; 2.每千克矿物质预混料含有Mineral premix contains the following per kg:碘酸钾 KIO340.03 g,氯化钴 CoCl24.07 g,硫酸铜 CuSO419.84 g,柠檬酸铁 FeC6H5O713.71 g,硫酸锌 ZnSO428.28 g,硫酸镁 MgSO40.12 g,硫酸锰 MnSO412.43 g,氯化钾 KCl 15.33 g,亚硒酸钠 Na2SeO32.00 g,沸石粉 zeolite power 864.19 g; 3.钙磷添加组用乳酸钙和磷酸二氢钠替代基础饲料组中的微晶纤维素. In the calcium and phosphorus addition group, microcrystalline cellulose in basic diet group is replaced with calcium lactate and sodium dihydrogen phosphate.
1.2 方法
试验开始前,给试验虾投喂基础饲料一周,使对虾适应饲料。试验共设15个处理,每个处理3个平行,每平行为1个0.30 m3的玻璃钢桶。对虾禁食24 h后,挑选规格均匀、健壮、活力强、初体质量(4.2±0.15) g的对虾,随机分配于玻璃钢桶中,每桶40尾,养殖8周。日投喂4次(7:00、11:00、17:00、21:00),饱食投喂,投喂1 h后查料,根据对虾摄食及天气情况调整投喂量。养殖期间24 h连续充气。对虾养殖前两周每2 d换水1次,后期日换水1次。试验期间水温28~31 ℃,盐度21~24,pH 7.8~8.2,溶解氧≥5 mg/L,氨氮<0.2 mg/L,亚硝酸盐<0.05 mg/L。
试验结束时禁食24 h后称量质量,计数,计算生长指标。然后每桶随机取15尾虾,逐尾用1 mL注射器自第5步足基部血窦取血,合并置于Eppendor管中,4 ℃冰箱保存过夜,以3000 r/min离心10 min后收集血清,-80 ℃冰箱保存备测血清指标。另每桶取10尾虾,沥干水后装于封口袋中,剩余的对虾剥离虾壳和肌肉,-20 ℃冰箱保存,用于测定常规成分和钙、磷含量。
饲料、全虾及肌肉样品的水分测定采用105 ℃烘干恒定质量法,粗蛋白采用凯氏定氮法(KjeltecTM8400,瑞典);粗脂肪采用索式抽提法(抽提剂为石油醚);粗灰分采用马福炉550 ℃灼烧法测定。
饲料、全虾、肌肉和虾壳的钙、磷含量的测定方法:样品烘干粉碎后,置于具塞试管中,加入5 mL默克硝酸,70 ℃水浴消化2 h,然后在消化炉上赶酸5 h,加入1 mL双氧水,赶酸至余下1 mL稀释。最后用电感耦合等离子体发射光谱仪(ICP)测定钙、磷含量。
血清指标测定方法:碱性磷酸酶活性采用南京建成试剂盒方法测定;酚氧化酶活性参照文献[13]的方法测定;总胆固醇、甘油三酯、钙离子和无机磷含量用全自动生化分析仪(日立7020HITACHI,日本)检测。
各项生长指标按下式计算:
2.1.3 供试品溶液的制备 取本品粉末(过4号筛)约0.3 g,精密称定,置具塞锥形瓶中,精密加甲醇50 mL,密塞,摇匀,称定质量,放置过夜,超声处理(功率250 W,频率50 kHz)30 min,取出,放冷,再称定质量,用甲醇补足减失的质量,摇匀,滤过,取续滤液,过微孔滤膜,得到供试品溶液。
质量增加率/%=(mt-m0)/m0×100%
特定生长率/%·d-1= (lnmt-lnm0)/t×100%
饲料系数=mf/(mt-m0)
蛋白质效率=(mt-m0)/(mf×P)
存活率/%=nt/n0×100%
式中,mt、m0分别为终末均质量、初始均质量,mf为饲料摄入量,P为饲料蛋白质含量,nt、n0分别为终末尾数、初始尾数。
1.3 数据处理
试验结果用平均值±标准差表示,用SPSS 17.0进行双因素方差分析,若存在显著性差异,再进行Duncan′s多重比较,以检验组间的差异显著性;P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 饲料钙、磷添加水平对凡纳滨对虾生长性能的影响
本试验中,饲料钙水平、磷水平及钙和磷水平的交互作用均极显著影响对虾的质量增加率、特定生长率、蛋白质效率、饲料效率和存活率(P<0.01)(表2)。0.5%钙水平和0.8%磷水平与不添加钙水平、磷水平组相比,极显著提高了对虾的质量增加率、特定生长率、蛋白质效率,并降低了饲料效率(P<0.01),钙水平、磷水平极显著提高了对虾的质量增加率、特定生长率和蛋白质效率,降低了饲料效率(P<0.01),0.5%钙水平与不添加钙水平组相比显著提高了存活率(P<0.05)。质量增加率和特定生长率随着磷水平的提高均呈先升后降的趋势,0.8%磷水平和1.2%磷水平的质量增加率和特定生长率差异不显著(P>0.05),但显著高于其他水平组(P<0.05)。0.8%~1.6%组的饲料效率差异不显著(P>0.05),但显著低于其他各组(P<0.05)。1.6%磷水平与不添加磷水平组相比显著降低了存活率(P<0.05)。
以特定生长率为判断依据,通过二次回归曲线分析得出:饲料中不添加钙时,饲料中可添加1.12%的磷;添加0.5%的钙时,饲料中可添加0.88%的磷;添加1%的钙时,饲料中可添加0.78%的磷。对虾在添加0.5%的钙和0.88%的磷时有最大特定生长率(图1)。
2.2 饲料中钙、磷添加水平对凡纳滨对虾全虾及肌肉常规成分含量的影响
饲料钙水平极显著影响对虾的肌肉粗蛋白、粗脂肪、粗灰分含量,全虾粗蛋白、粗脂肪的含量(P<0.01)(表3)。饲料磷水平极显著影响对虾的肌肉粗蛋白、粗脂肪和全虾粗蛋白的含量(P<0.01)。饲料钙、磷水平的交互作用极显著影响对虾的全虾粗蛋白和肌肉粗蛋白、粗脂肪的含量(P<0.01),显著影响对虾的全虾粗脂肪的含量,对全虾和肌肉粗灰分差异不显著(P<0.05)。钙水平极显著提高了全虾和肌肉粗蛋白的含量,降低了肌肉粗脂肪、粗灰分及全虾粗脂肪的含量,0.5%~1.0%钙水平极显著降低了肌肉粗脂肪、粗灰分的含量(P<0.01)。随磷水平的升高,全虾粗蛋白的含量呈先升后降的趋势,1.2%组全虾粗蛋白含量极显著高于其他处理(P<0.01)。1.6%磷水平提高了全虾粗灰分的含量,但与1.2%磷水平之间无显著性差异(P>0.05)。对虾肌肉粗灰分含量随磷水平的升高呈上升趋势,但各组之间差异不显著(P>0.05)。
2.3 饲料中钙、磷水平对凡纳滨对虾组织钙、磷含量的影响
饲料钙水平极显著地影响全虾和虾壳的钙、磷含量和肌肉的磷含量(P<0.01),显著影响肌肉的钙含量(P<0.05)(表4)。磷水平及钙磷水平的交互作用均极显著地影响全虾、肌肉和虾壳的钙、磷含量(P<0.01)。钙水平极显著提高了全虾的钙、磷含量、虾壳的磷含量(P<0.01)。随着钙水平的提高肌肉磷含量呈先升后降的趋势,0.5%钙水平组肌肉磷含量显著高于其他水平(P<0.05)。全虾、肌肉和虾壳的钙、磷含量随着饲料磷水平的提高均呈先增后减的趋势,1.2%磷水平全虾的钙、磷含量及虾壳的钙含量显著高于其他水平(P<0.05)。

表2 饲料钙、磷添加水平对凡纳滨对虾生长性能的影响Tab.2 Effects of dietary Ca and P supplemental levels on growth performance of Pacific white shrimp L.vannamei
注: 同一列中上标不同字母的平均值间差异显著(P<0.05),各组上标中有相同字母表示组间差异不显著(P>0.05),下同.
Note: the means with different superscripts in the same column are significantly different (P<0.05); the means with same superscripts indicate no significant difference among groups (P>0.05); et sequentia.

图1 3个不同钙添加水平下凡纳滨对虾饲料中磷的最适添加量Fig.1 The optimum supplemental level of P in diet for Pacific white shrimp L. vannamei under three Ca supplemental levels
表3饲料钙磷添加水平对凡纳滨对虾全虾及肌肉常规成分的影响(干物质基础) %

Tab.3 Effects of dietary Ca and P supplemental levels on approximate composition of whole body andmuscle of Pacific white shrimp L.vannamei(DM basis)

表4 饲料钙磷添加水平对凡纳滨对虾机体钙磷含量的影响Tab.4 Effects of dietary Ca and P supplemental levels on Ca and P levels in tissues of Pacific white shrimp L.vannamei
(续表4)

添加量 Supplemental amount全虾 Whole body肌肉 Muscle虾壳 Exoskeleton钙(总钙)Ca (TCa)磷(总磷)P (TP)钙含量Ca content磷含量P content钙含量Ca content磷含量P content钙含量Ca content磷含量P content0.5(2.17)1.2(2.74)40.90±0.86h20.20±0.43fg6.13±0.18abc14.73±0.68cd156.85±1.96i16.30±0.07gh0.5(2.18)1.6(3.11)37.69±1.72efg19.38±0.62ef5.81±0.10a14.44±0.11cd150.82±0.59gh14.82±0.27f1.0(2.69)0(1.38)33.08±0.82c15.03±0.72bc5.86±0.21a11.61±0.12b138.08±1.15a8.95±0.09b1.0(2.68)0.4(1.83)35.92±0.40de16.68±0.32d6.78±0.15de14.21±0.45c144.89±0.47cd13.70±0.37de1.0(2.68)0.8(2.31)38.96±1.05gh18.76±0.29e6.53±0.04cde15.71±0.22ef148.88±0.49fg16.50±0.40h1.0(2.63)1.2(2.70)44.10±0.78i21.21±0.83g6.47±0.01bcde14.46±0.24cd157.06±1.17i16.69±0.19h1.0(2.70)1.6(3.16)39.15±0.81gh20.20±0.46fg6.07±0.07ab14.05±0.10c152.82±0.55h15.66±0.33g钙水平/% Ca level033.89±4.10a15.51±2.25a6.49±0.53b13.79±2.60a144.74±5.45a13.29±2.97a0.536.54±3.71b17.59±2.37b6.29±0.42a14.37±1.39b147.47±6.74b13.83±3.13b1.038.24±3.92c18.37±2.42c6.34±0.36ab14.01±1.42a148.34±6.92b14.30±3.04c磷水平/% P level030.68±2.24a14.12±1.03a6.10±0.28a10.92±1.46a137.79±1.19a8.42±0.47a0.434.04±2.45b15.39±1.51b6.97±0.23c14.32±0.41b143.16±1.69b13.40±0.45b0.837.35±2.08c17.42±1.70c6.55±0.20b15.94±0.25c147.80±1.28c16.11±0.50d1.241.36±2.35d20.06±1.17e6.37±0.21b14.78±0.51b155.20±2.98e16.19±0.59d1.637.67±1.63c18.80±1.62d5.88±0.17a14.33±0.33b150.31±2.59d14.92±0.65c双因素方差分析P值 P-value of two-factor ANOVA钙水平 Ca level0.0000.0000.0410.0090.0000.000磷水平 P level0.0000.0000.0000.0000.0000.000钙水平×磷水平 Ca level×P level0.0000.0000.0010.0000.0000.000
2.4 饲料中钙、磷的添加水平对凡纳滨对虾血清指标的影响
饲料中钙、磷的水平均极显著影响血清碱性磷酸酶、酚氧化酶的活性和总胆固醇、甘油三酯的含量(P<0.01)(表5)。钙、磷水平的交互作用极显著影响血清碱性磷酸酶的活性和钙离子、无机磷的含量(P<0.01)。饲料钙、磷水平及钙、磷水平的交互作用对虾血清钙离子的含量影响不显著(P>0.05)。钙水平显著提高了血清碱性磷酸酶、酚氧化酶的活性及总胆固醇的含量(P<0.05)。1%钙水平组对虾血清酚氧化酶的活性极显著高于其他水平组(P<0.01)。随着钙水平的提高对虾血清甘油三酯的含量呈先升后降的趋势,0.5%钙水平组显著高于其他水平组(P<0.05)。磷水平显著降低了血清碱性磷酸酶的活性和甘油三酯的含量,1.2%~1.6%磷水平也显著降低了总胆固醇的含量(P<0.05)。酚氧化酶的活性和无机磷的含量均随着磷水平的提高呈先升后降的趋势,0.8%磷水平组的酚氧化酶活性显著高于其他水平组(P<0.05)。

表5 饲料中钙、磷添加水平对凡纳滨对虾血清指标的影响Tab.5 Effects of dietary Ca and P supplemental levels on serum indices of Pacific white shrimp L.vannamei
(续表5)
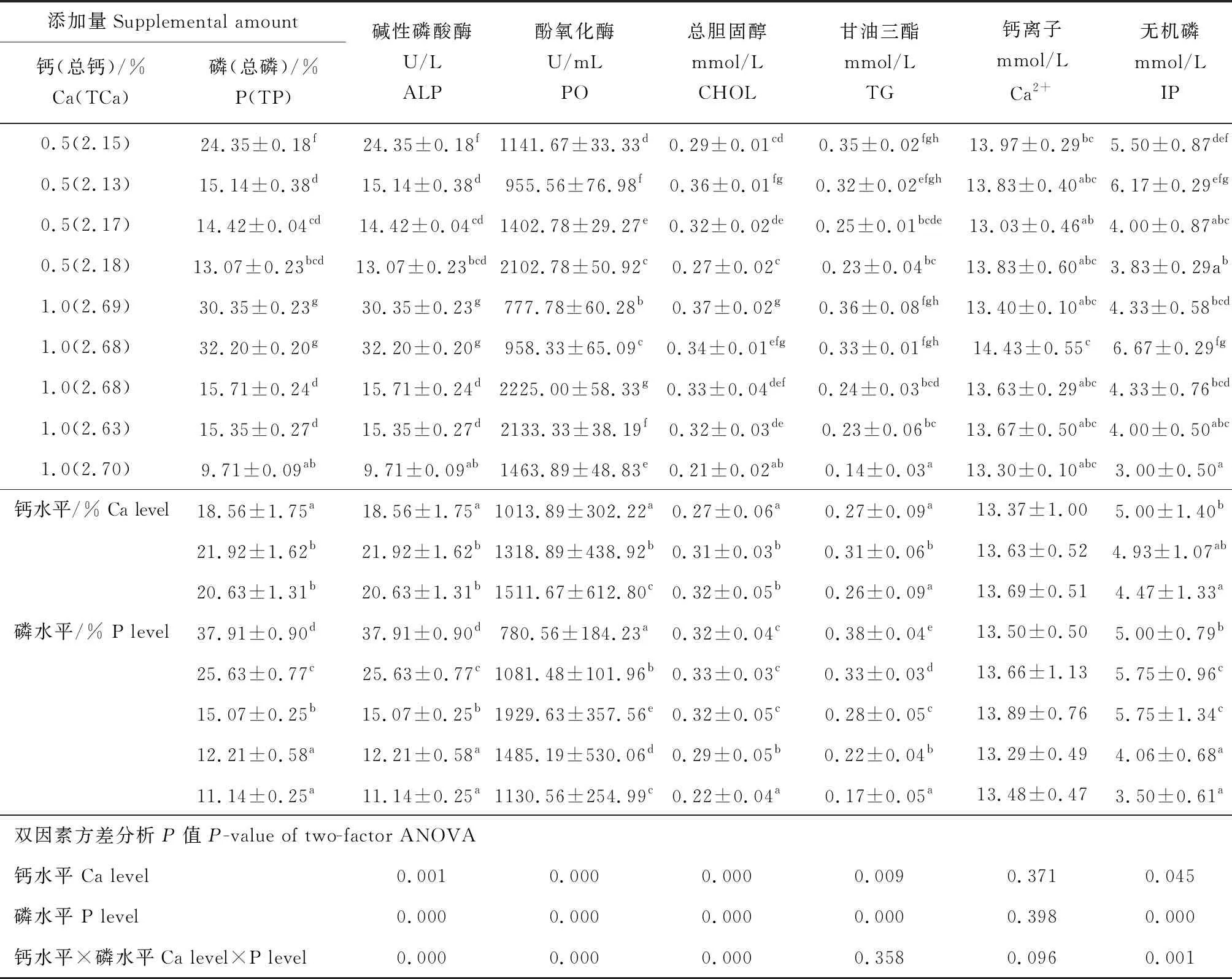
添加量Supplemental amount钙(总钙)/%Ca(TCa)磷(总磷)/%P(TP)碱性磷酸酶U/LALP酚氧化酶U/mLPO总胆固醇mmol/LCHOL甘油三酯mmol/LTG钙离子mmol/LCa2+无机磷mmol/LIP0.5(2.15)24.35±0.18f24.35±0.18f1141.67±33.33d0.29±0.01cd0.35±0.02fgh13.97±0.29bc5.50±0.87def0.5(2.13)15.14±0.38d15.14±0.38d955.56±76.98f0.36±0.01fg0.32±0.02efgh13.83±0.40abc6.17±0.29efg0.5(2.17)14.42±0.04cd14.42±0.04cd1402.78±29.27e0.32±0.02de0.25±0.01bcde13.03±0.46ab4.00±0.87abc0.5(2.18)13.07±0.23bcd13.07±0.23bcd2102.78±50.92c0.27±0.02c0.23±0.04bc13.83±0.60abc3.83±0.29ab1.0(2.69)30.35±0.23g30.35±0.23g777.78±60.28b0.37±0.02g0.36±0.08fgh13.40±0.10abc4.33±0.58bcd1.0(2.68)32.20±0.20g32.20±0.20g958.33±65.09c0.34±0.01efg0.33±0.01fgh14.43±0.55c6.67±0.29fg1.0(2.68)15.71±0.24d15.71±0.24d2225.00±58.33g0.33±0.04def0.24±0.03bcd13.63±0.29abc4.33±0.76bcd1.0(2.63)15.35±0.27d15.35±0.27d2133.33±38.19f0.32±0.03de0.23±0.06bc13.67±0.50abc4.00±0.50abc1.0(2.70)9.71±0.09ab9.71±0.09ab1463.89±48.83e0.21±0.02ab0.14±0.03a13.30±0.10abc3.00±0.50a钙水平/% Ca level18.56±1.75a18.56±1.75a1013.89±302.22a0.27±0.06a0.27±0.09a13.37±1.005.00±1.40b21.92±1.62b21.92±1.62b1318.89±438.92b0.31±0.03b0.31±0.06b13.63±0.524.93±1.07ab20.63±1.31b20.63±1.31b1511.67±612.80c0.32±0.05b0.26±0.09a13.69±0.514.47±1.33a磷水平/% P level37.91±0.90d37.91±0.90d780.56±184.23a0.32±0.04c0.38±0.04e13.50±0.505.00±0.79b25.63±0.77c25.63±0.77c1081.48±101.96b0.33±0.03c0.33±0.03d13.66±1.135.75±0.96c15.07±0.25b15.07±0.25b1929.63±357.56e0.32±0.05c0.28±0.05c13.89±0.765.75±1.34c12.21±0.58a12.21±0.58a1485.19±530.06d0.29±0.05b0.22±0.04b13.29±0.494.06±0.68a11.14±0.25a11.14±0.25a1130.56±254.99c0.22±0.04a0.17±0.05a13.48±0.473.50±0.61a双因素方差分析P值 P-value of two-factor ANOVA钙水平 Ca level0.0010.0000.0000.0090.3710.045磷水平 P level0.0000.0000.0000.0000.3980.000钙水平×磷水平Ca level×P level0.0000.0000.0000.3580.0960.001
3 讨 论
3.1 饲料钙、磷水平对凡纳滨对虾生长性能的影响
Ambasankar等[11]研究发现,0~2.5%磷水平显著提高了斑节对虾的质量增加率。0.71%~1.59%磷水平显著提高了黑鲷(Acanthopagrusschiegelii)幼鱼的质量增加率、特定生长率和蛋白质效率[14]。本试验结果与上述研究结果一致,也与凡纳滨对虾[15]、中华绒螯蟹(Eriocheirsinensis)[16]的研究结果一致。斑节对虾的质量增加率在1.5%磷水平达到最大值,而2%磷水平组的质量增加率显著降低[11]。本试验结果也表明,随着磷水平的升高,凡纳滨对虾的质量增加率、特定生长率和蛋白质效率呈先升后降的趋势,在1.6%磷水平时质量增加率、特定生长率显著下降。印度明对虾的饲料效率随饲料磷水平的升高呈先降后升的趋势(饲料钙水平为1.25%)[12]。黑线鳕(Melanogrammusaeglefinus)[17]、黑鲷幼鱼[18]、军曹鱼(Rachycentroncanadum)[19]、草鱼(Ctenopharyngodonidella)[20]的研究中也有类似结论。这说明饲料中高水平的磷会抑制对虾对其他矿物元素的吸收,从而抑制其生长,磷的添加量必须控制在适宜的范围内。饲料中磷含量过高会导致草鱼生长缓慢,甚至死亡[20-21]。Pan等[15]发现,0.91%~1.63%的磷水平不影响体质量0.16 g的凡纳滨对虾的生长,这可能与幼虾阶段生长速度快,蜕壳次数多,需要较多的磷有关。Niu等[22]也发现,早期对虾对磷的需要量更高。
Davis等[9]研究发现,若未添加钙,基础饲料中含0.35%的磷就可以维持对虾的生长和存活;若添加1%钙,则添加2%的磷时对虾质量增加率和存活率最大。Cheng等[10]报道,盐度为2的水体中养殖的对虾在投喂无钙饲料时,添加0.77%的有效磷(0.93%总磷)就能满足其生长;当添加1%的钙时,饲料中需添加1.22%的有效磷(2%的总磷),对虾才具有较好的生长性能。黄凯等[23]以磷酸二氢钙作为凡纳滨对虾饲料的钙源和磷源,在盐度为2的水体中,发现磷酸二氢钙添加量为1%时,对虾的生长速度最快;当添加量为3%时,对虾的生长性能最佳。而Pan等[15]同样也是用磷酸二氢钙作为饲料的钙源和磷源,在盐度17~20的海水中养殖,发现当磷酸二氢钙添加量为1.7%(总钙为1.83%,总磷为1.33%)时,凡纳滨对虾生长和饲料转化率最佳。本试验中二次回归曲线拟合得出,随饲料钙水平的升高,获得最佳生长性能的磷需要量却是逐渐下降的,不同于已有的研究结果;主因素分析表明,适量的钙水平(0.5%)可显著提高对虾的生长性能指标和成活率,显著降低饲料系数。本试验中对虾的初始质量为(4.2±0.15) g,远远高于上述研究中的幼虾体质量,同时养殖水体盐度为21~24。由此可见,即使同种虾钙、磷需求量也不尽相同,这可能与养殖对象的生长阶段、养殖水体盐度、饲料配方、磷源与钙源等因素有关。钙、磷间有显著的交互作用,但二者的相互作用还有待进一步研究。
3.2 饲料中钙、磷水平对凡纳滨对虾全虾及肌肉常规成分的影响
Niu等[22]研究发现,凡纳滨对虾全虾粗蛋白含量随饲料磷水平的上升而显著升高。随饲料磷水平(0.72%~1.74%)的升高,红螯螯虾(Cheraxquadricarinatus)[3]肌肉粗蛋白含量上升。本试验结果表明,磷水平提高了凡纳滨对虾的全虾和肌肉中粗蛋白的含量。类似的还有黑线鳕[17]、日本花鲈(Lateolabraxjaponicus)[24]、斑点叉尾(Ietaluruspunetaus)[25]和黑鲷幼鱼[14]。本试验中,随着磷水平的升高,凡纳滨对虾全虾粗脂肪的含量呈下降趋势,肌肉粗脂肪的含量在1.6%磷水平显著下降。这与红螯螯虾[3]、凡纳滨对虾[22]、草鱼[21]和虹鳟(Oncorhynchusmykiss)[26]等研究结果一致。Roy等[17]认为,这可能是因为磷缺乏抑制了动物体内脂肪酸的β-氧化,脂肪的利用率降低,转而利用蛋白质供能,导致机体脂肪累积和蛋白含量减少;而随着饲料中磷含量的逐渐升高,增强了动物体内脂肪酸的β-氧化,从而增强机体利用脂肪供能的能力,增加糖原的生成,促使蛋白质沉积,所以机体的蛋白含量会增加而脂肪含量减少。灰分和磷含量是水产动物磷营养研究中的常用指标[26],本试验中,随着饲料磷水平的升高,对虾的全虾和肌肉粗灰分的含量呈上升趋势。中华绒螯蟹[27]、红螯螯虾[3]、克原氏螯虾(Procambarusclarkii)[28]和中国明对虾[29]的研究也得到了类似的结果。罗文佳等[5]认为,这可能是磷含量的增加和离子之间的协同作用增加了矿物质在机体中的沉积。
3.3 饲料中钙、磷水平对凡纳滨对虾组织钙、磷含量的影响
鱼虾机体的钙、磷含量,常常作为研究鱼虾饲料钙、磷适宜添加量的参考指标[30-31]。Pan等[15]研究发现,随着饲料磷水平(0.91%~1.63%)的升高,凡纳滨对虾全虾和虾壳的钙、磷含量呈上升趋势。同一钙水平下,对虾的肌肉钙、磷含量随饲料磷水平的升高而增加[10]。本试验中,对虾的全虾、肌肉、虾壳的钙和磷含量随饲料磷水平的升高而升高,与上述结果,也与红螯螯虾[3]、克原氏螯虾[28]、中华绒螯蟹[27]等报道相一致。任泽林等[29]发现,中国明对虾饲料钙水平与虾壳钙含量之间,饲料磷水平与虾壳磷含量及肌肉磷含量之间呈线性关系。凡纳滨对虾全虾及虾壳的钙、磷含量与饲料中钙、磷的水平呈正相关联系[5]。大麻哈鱼(Oncorhynchusketa)[32-33]、虹鳟[34-36]的研究也证实了鱼体钙、磷含量与饲料磷水平之间存在正相关的关系。但有研究表明,印度明对虾体内磷含量与饲料磷添加量无关[37];Davis等[38]的研究也表明,饲料中钙、磷含量与对虾组织矿物元素含量没有直接相关性;其他凡纳滨对虾[39]研究及美洲龙虾(Homarusamericanus)[40]等研究中也有类似的研究结论。这可能与虾在不同生长阶段的蜕皮循环有关,因为甲壳动物蜕皮前后甲壳和肌肉中钙、磷含量会变化,在蜕壳过程中矿物元素会反复损失[23,40]。
3.4 饲料中钙、磷水平对凡纳滨对虾血清指标的影响
碱性磷酸酶是一种磷酸单酯酶,可催化磷酸的水解反应及基团的转移反应,为二磷酸腺苷磷酸化形成三磷酸腺苷提供无机磷酸[41]。碱性磷酸酶是一种水解磷酸酯及焦磷酸酯的水解同工酶,其活性受到饲料磷水平的影响,可作为动物体内磷酸状态的评价指标之一[42]。本试验中,饲料磷水平显著降低了对虾血清碱性磷酸酶的活性。饲料磷水平0.5%~2.0%使凡纳滨对虾血清碱性磷酸酶活性显著降低[10]。这与本试验研究结果一致,也与真鲷(Pagrosomusmajor)[31]、日本沼虾(M.nipponensis)[43]等结果相一致。但随饲料磷水平的升高,红螯螯虾血清碱性磷酸酶活性也显著升高[3],在日本花鲈[24]、虹鳟[26]中也有相似结果。但斑点叉尾血清碱性磷酸酶活性与饲料磷水平间无显著相关性[25]。这可能是碱性磷酸酶的活性受到水化学[44]、采食量[45]和生长阶段[46]等的影响。
酚氧化酶原激活系统是一种酶级联系统,甲壳动物的酚氧化酶原系统存在于血淋巴中的颗粒细胞中[47]。系统中的酚氧化酶原可以被细菌脂多糖、钙离子及胰蛋白酶等激活后转变成具有活性的酚氧化酶[48]。Sung等[49]研究发现,镁离子、钙离子能增强斑节对虾和罗氏沼虾(M.rosenbergii)酚氧化酶的活性。本试验饲料钙水平提高了凡纳滨对虾的血清酚氧化酶的活性,与上述研究一致;而随着磷水平的提高,凡纳滨对虾血清酚氧化酶活性均呈先升后降的趋势。这可能是饲料中的磷主要以磷酸根的形式存在,过多的磷酸根与铁、镁等金属离子反应生成不溶性磷酸盐,从而影响铁、镁等离子的消化吸收和血清酚氧化酶的活性[50]。
本试验中,饲料磷水平对血清钙离子无显著性差异。在黄颡鱼(Pelteobagrusfulvidraco)[51]中也有类似报道。唐琴等[51]认为,饲养试验结束后,饥饿24 h后取样,血清的钙、磷含量已不能反映鱼体的摄食情况。本试验中,饲料磷水平显著降低了对虾的血清总胆固醇和甘油三酯含量。0.61%~1.51%磷水平降低了军曹鱼血清总胆固醇和甘油三酯含量[19]。黑鲷血清总胆固醇和甘油三酯含量随饲料磷水平的升高而降低[18]。本试验结果与上述研究结果一致,也与大黄鱼(Pseudosciaenacrocea)[52]、日本花鲈[24]、双棘黄姑鱼(Protonibeadiacanthus)[53]报道相同。黄晓聪等[53]认为,这与脂肪代谢有关,磷缺乏导致脂肪生成增多,当磷足够时脂肪生成减少,总胆固醇和甘油三酯的含量下降。
4 结 论
初体质量(4.2±0.15) g的凡纳滨对虾,在水温28~31 ℃、盐度21~24条件下,饲料中添加0%、0.5%、1%的钙时,饲料中磷的适宜添加水平为1.12%、0.88%、0.78%。在添加0.5%钙和0.88%的磷时凡纳滨对虾特定生长率最大。
猜你喜欢
今日农业(2022年15期)2022-09-20
当代水产(2022年8期)2022-09-20
当代水产(2022年7期)2022-09-20
南方农业学报(2022年5期)2022-08-09
水产科学(2022年3期)2022-06-08
当代水产(2022年5期)2022-06-05
当代水产(2022年3期)2022-04-26
大连海洋大学学报(2021年6期)2022-01-10
当代水产(2021年8期)2021-11-04
当代水产(2021年4期)2021-07-20
