
噬菌体展示系统筛选糖尿病相关抗原蛋白及应用效果评价
2019-12-23 08:04刘尧林骏梁永羿山云王洪涛徐良
中国医药生物技术 2019年6期
刘尧,林骏,梁永羿,山云,王洪涛,徐良
论著
噬菌体展示系统筛选糖尿病相关抗原蛋白及应用效果评价
刘尧,林骏,梁永羿,山云,王洪涛,徐良
用噬菌体展示系统筛选与糖尿病相关的自身抗原新表位,表达新抗原肽段并评价其在糖尿病诊断中的应用效果。
以 I 型糖尿病血清作为靶分子,对 Ph.D.-12 噬菌体肽库进行淘选,测序淘选的噬菌体并用 ELISA 检测噬菌体与靶分子的结合效果,BLAST 分析特异性最好的表位与糖尿病的相关性,选择合适的目的基因合成重组质粒,用 Expi 293 细胞表达新抗原肽段并检测该抗原与临床血清的特异结合效果。
共完成 2418 个噬菌体单克隆的测序,淘选出 93 种表位序列,11 种具有代表性的表位中测序超过 200 次的有 4 种,对特异性最强的表位序列“-GTFLFSLCAAVY”进行分析后选择 mTOR 相关蛋白 mEAK-7 作为目的基因,抗原肽段与临床血清的特异性结合效果为:I型糖尿病阳性率为 17%,II型糖尿病阳性率为 4%。
mEAK-7 蛋白对糖尿病血清具有一定的抗原特异性,该基因与糖尿病可能有相关性,具有很好的研究价值。
噬菌体展示系统; 糖尿病; 自身抗体; 抗原表位
I 型糖尿病是由于自身免疫系统错误攻击和损伤胰岛β 细胞,使胰岛素合成和分泌减少或者完全缺失的自身免疫性疾病,临床上大多数 I 型糖尿病患者可被检测到胰岛自身抗体,目前来说,GADA、IAA、ICA、ZnT-8A、IA2A 这 5 种胰岛自身抗体是 I 型糖尿病检测应用最广泛、最受认可、相关性最强的自身免疫标志物[1-2]。本研究通过噬菌体展示系统获得与糖尿病相关的自身抗原新表位的氨基酸序列进行自身抗原肽段的表达,评估包含抗原表位在内人工构建的蛋白多肽对于糖尿病诊断的临床价值和意义。
1 材料与方法
1.1 实验材料、试剂
噬菌体展示试剂盒和ER 2738 购自 Biolab 公司;培养基组分、Xgal、IPTG 购自美国 Sigma 公司;抗-M13 抗体(HRP)购自英国 Abcam 公司;限制性内切酶和Expi 293 真核表达系统购自美国 Thermo Fisher 公司;去内毒素质粒抽提试剂盒和蛋白电泳试剂购自生工生物工程(上海)股份有限公司;ELISA 板购自美国康宁公司;TMB 单组分显色液购自北京索莱宝科技有限公司;糖尿病抗体检测试剂盒、全自动免疫印迹分析仪购自深圳市亚辉龙生物科技股份有限公司;全波长酶标仪购自美国 Thermo Fisher 公司;菌株DH5α 由本实验室保存;其他化学试剂为分析纯以上等级。
1.2 方法
1.2.1 噬菌体展示系统的淘选方法 分别收集来自健康人、I 型糖尿病、II 型糖尿病患者的血清,并将健康人,I 型糖尿病血清包被于 96 孔板中,150 μl/孔。TBST 溶液洗板 6 次,添加封阻液封阻,取 100 μl 原始噬菌体文库(2 × 1011PFU)添加到包被有健康人血清的孔里,室温作用 1 h,回收未结合的噬菌体液。将回收的噬菌体液添加到包被I型糖尿病血清的孔里,室温作用 1 h,TBST 洗板 10 次,用洗脱液洗脱与I型糖尿病血清特异性结合的噬菌体。将特异性噬菌体感染至大肠杆菌 ER 2738 中扩增 4.5 h,离心,取上清,用 PEG/NaCl 4 ℃沉淀过夜。次日将沉淀物离心,弃上清,将沉淀物溶于 TBST 溶液中,此即扩增的噬菌体。将淘选后的噬菌体进行滴度测定、扩增,进入下一个循环淘选。每一轮进行 4 个循环淘选,一共进行4 轮淘选。
1.2.2 测序淘选的噬菌体 取每个循环淘选后未扩增的噬菌体液 5 μl 感染至大肠杆菌 ER 2738 并涂于含有四环素的 LB 培养皿上,37 ℃孵育过夜,随机挑取 160 个单克隆噬菌斑,扩增后将单克隆噬菌体送至上海生工进行测序。剩余噬菌体液保存于 4 ℃用于后续 ELISA 检测实验,或者用灭菌甘油 1:1 稀释后,–20 ℃贮存。
1.2.3 ELISA 检测噬菌体对靶分子的特异性 对所有序列的测序数据进行统计和分析,将测序频次高于 200 的噬菌体进行扩增并滴定。准备两个96 孔微孔板,每个待鉴定克隆分别在每个板上对应一列孔(12 个孔)。板 1 用于包被 I 型糖尿病血清后进行 ELISA 检测所选序列与靶分子的结合力;板 2 仅用封阻液封阻后作为空白对照用于检测所选序列对 BSA 包被塑料板的结合力。将 1 × 1012PFU 的噬菌体进行 5 倍系列梯度稀释至第 12 孔,浓度分别为:1 × 1012、2 × 1011、4 × 1010、8 × 109、1.6 × 109、3.2 × 108、6.4×107、1.28 × 107、2.56 × 106、5 × 105、1 × 105和 2 × 104PFU,用多通道移液器将稀释好的噬菌体吸取 100 μl 加入包被有靶分子的板 1 的对应孔中,再吸取 100 μl加入板 2 的对应孔中,每个浓度梯度做 3 个重复。室温振荡作用 1 h,洗板 6 次,添加稀释后的抗-M13 抗体(HRP 标记),200 μl/孔,室温作用1 h,洗板 6 次后每孔加入 200 μl 底物溶液,室温作用 20 min,酶标仪检测 405 nm 处的吸光度。
1.2.4 分析表位序列与糖尿病的相关性 将上述验证特异性最好的表位序列导入 NCBI 数据库中进行 BLAST 分析,查找与该表位序列具有同源关系的人源的氨基酸或蛋白片段,分析其与糖尿病、胰岛 β 细胞或者相关疾病的关系,定位与糖尿病自身抗体相关的抗原表位。
1.2.5 重组蛋白表达 选择与糖尿病相关性最大的基因,设计与之对应的核苷酸编码序列,以定位的抗原表位为中心向该基因上下游分别延伸约200 个碱基作为目的基因,选取pcDNA 3.1 作为真核表达载体,合成重组质粒。将合成的重组质粒酶切并电泳鉴定后扩大培养,抽提无内毒素质粒并转染进 Expi 293 细胞中进行重组蛋白的表达,收集表达的细胞培养液,电泳检测表达效果。
1.2.6 ELISA 检测重组蛋白的抗原特异性 将纯化的重组蛋白稀释至 1 μg/ml,添加到 96 孔板中,100 μl/孔,在 4 ℃包被过夜;PBST 溶液洗板 3 次,加入封阻液封阻,用样品稀释液分别将健康人、I 型糖尿病、II 型糖尿病患者的血清稀释 100 倍,添加到 96 孔板中,每个样本一个孔,室温孵育 30 min,洗板 3 次,添加稀释后的 HRP 标记的兔抗人 IgG,100 μl/孔,室温作用 30 min,洗板 3 次后每孔加入 100 μl 的 TMB 显色液,室温孵育 15 min,每孔加入100 μl 终止液,酶标仪检测各孔 450 nm 吸光度。
1.2.7 与其他糖尿病自身抗体对比 用糖尿病抗体检测试剂盒和全自动免疫印迹分析仪分别对健康人、I 型糖尿病、II 型糖尿病患者的血清进行检测,统计 GADA、IAA、ICA、ZnT-8A、IA2A 的阳性率。
1.3 统计学处理
实验数据采用 GraphPad Prism 8.0 软件进行分析及作图,多组差异比较采用单因素方差分析,以< 0.05 为差异有统计学意义。
2 结果
2.1 淘选的表位序列测序结果
本实验每轮淘选出 640 个单克隆噬菌体(表 1),经过 4 轮淘选的送测序样本共计 2560 个,最终获得有效测序样本共计 2418 个,占总样本的 94.4%,共淘选出 128 种表位序列,其中 15 种表位序列出现的频次较高,具有一定代表性。排除各轮之间 35 种重复序列后得到特异表位序列 93 种,排除各轮之间 4 种重复代表性表位序列后得到代表性特异表位序列 11 种,代表性特异表位序列样本数为 1620 个(表 2),占有效测序样本的 67%。

表 1 淘选的表位序列结果
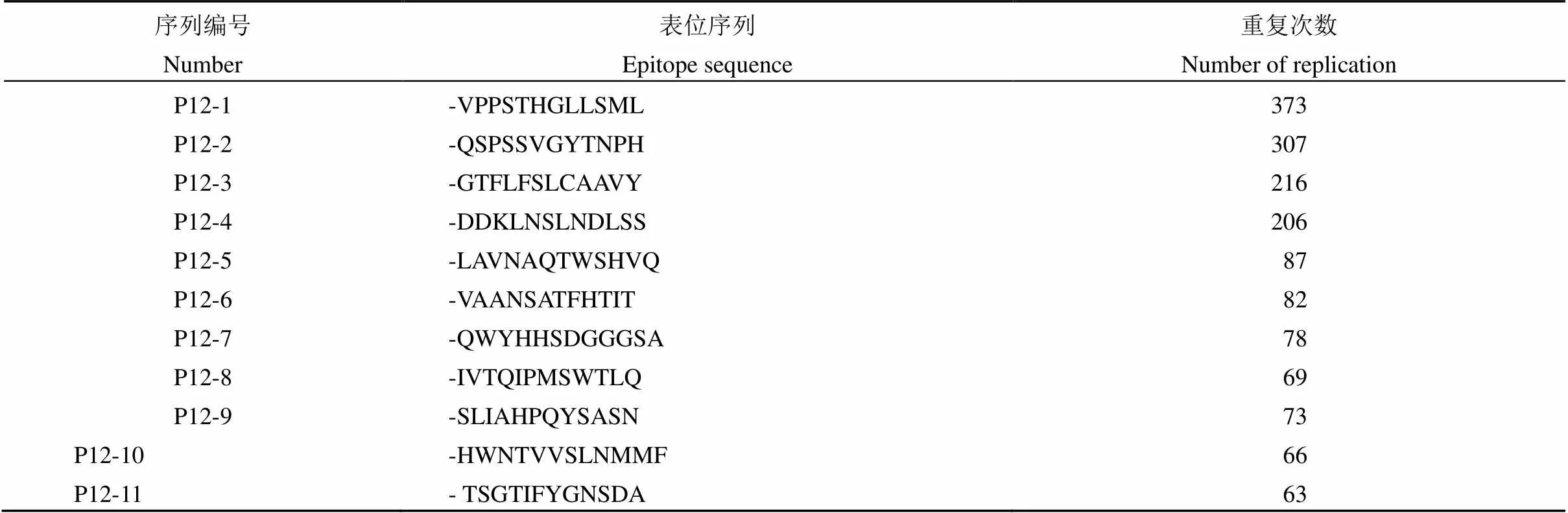
表 2 具有代表性的特异表位序列统计结果
2.2 ELISA 检测噬菌体对靶分子的特异性
ELISA 检测结果显示同一种噬菌体在浓度为 1 × 1012、2 × 1011和 4 × 1010PFU 时检测值差异较小,但 P12-3 在各个浓度梯度的检测值都与其他噬菌体差异较大(< 0.0001),比其他几个噬菌体的检测值高至少 3 倍。虽然 P12-1 和 P12-2 出现的频次高于 P12-3,但是 P12-3 的特异性明显强于其他几种噬菌体,特异性由强到弱分别是:P12-3 > P12-2 > P12-1 > P12-4(图 1A)。P12-1、P12-2 和 P12-4 只有在浓度大于 8 × 109PFU 时才有明显的检测效果,而 P12-3 在浓度介于 2.56 × 106~4 × 1010PFU 之间的检测结果有良好的线性关系(y = 3E + 09x2– 4E + 09x + 6E + 08,2= 0.9984)(图 1B)。
2.3 BLAST 分析多肽序列
将特异性最高 P12-3 的表位序列“-GTFLFSLCAAVY”导入 NCBI 数据库进行 BLAST分析,与该序列具有同源性的人源基因共有 60 个序列,统计后获得 5 个与该序列具有较高同源性的人源基因(表 3)。
2.4 重组质粒及真核表达的目的蛋白的鉴定
通过分析,选取与“-GTFLFSLCAAVY”同源性最高的 mTOR 相关蛋白 mEAK-7 作为目的基因并合成重组质粒,合成 DNA 分子量约为 480 bp(图 2A),经过真核细胞 Expi293 表达,重组的 mEAK-7 分子量约为 18 kD,符合预期重组蛋白的分子量大小(图 2B)。
2.5 ELISA 检测表达的重组蛋白的抗原特异性
将纯化后的重组蛋白作为抗原包被在 96 孔板上,用 ELISA 方法检测健康人、I 型糖尿病、II 型糖尿病血清(各 200 例)与重组蛋白的特异性反应,结果显示,健康人样本均呈现阴性结果,I 型糖尿病样本有 34 个呈现阳性结果,阳性率为 17%,II 型糖尿病样本有 8 个出现阳性结果,阳性检出率为 4%,说明 mEAK-7 蛋白对于糖尿病患者血清具有一定的特异性(图3)。

图 1 噬菌体与靶分子的特异性检测(A:ELISA检测不同表位的噬菌体在不同浓度条件下与靶分子的特异性识别;n = 3,*P < 0.0001;B:ELISA分析P12-3的检测线性范围)
Figure 1 Specific identification of phage and target molecules (A: Specific identification of target molecules and different epitopes at different concentrations by ELISA; n = 3,*< 0.0001; B: Linear range of ELISA analysis for P12-3)
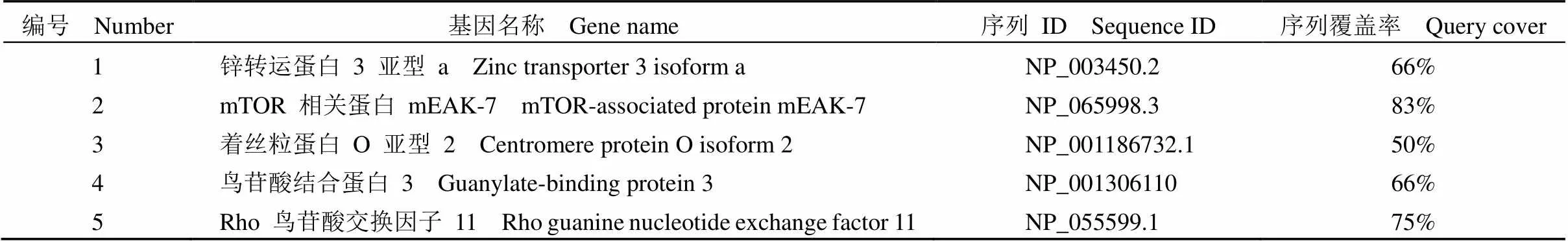
表 3 表位序列同源性分析结果

1:单酶切;2:双酶切;3:对照;4:mEAK-7;M:Marker
Figure 2 Identification of recombinant plasmids and recombinant proteins (A: Agarose gel electrophoresis analysis of recombinant plasmids; B: SDS-PAGE analysis of recombinant protein)

图 3 ELISA 检测重组蛋白与临床血清的结合效果
Figure 3 Identification of the binding effect of recombinant protein and clinical serum by ELISA
2.6 免疫印迹法检测其他糖尿病自身抗体阳性率
用免疫印迹法对健康人、I 型糖尿病、II 型糖尿病患者血清(各 200 例)进行检测,结果显示I 型糖尿病患者血清中 GADA 和 ZnT-8A 阳性率较高(表 4),分别为 61.5% 和 52%,ICA、IAA、IA-2A 的阳性率介于 17% ~ 20%,与 mEAK-7 的阳性检出结果近似;II 型糖尿病患者血清的 GADA阳性率也是最高的,达到 17.5%,ZnT-8A、ICA、IAA、IA-2A 的阳性率均小于 10%,而 ICA 和 IA-2A 的阳性率分别为 4.5% 和 4%,与 mEAK-7 的阳性检出结果类似。

表 4 GADA、ICA、IAA、ZnT-8A、IA-2A 的阳性率检测
3 讨论
mTOR 信号通路在生物体内众多细胞信号通路中占据相当重要的地位,参与调控如增殖、生长、分化、凋亡等多种生命现象,同时在炎症、肿瘤、代谢和心血管疾病的发病机制中起重要作用。已有文献报道 mTOR 信号通路与胰岛素和糖代谢相关,在长期高浓度的葡萄糖刺激下,信号通路 Insulin/PI3K/PKB 下游的 mTOR/S6K1 处于激活状态,过度表达而抑制胰岛素的信号传导,从而引起胰岛素抵抗[3-4]。目前关于 mEAK-7 相关功能的研究报道很少,也没有文献提出其与糖尿病相关,但有报道提出 mEAK-7 能够将 mTOR 招募到溶酶体中与其相互作用,通过 S6K2 和 4E-BP1 激活另一种 mTOR 信号通路,调控细胞增殖和迁移[5]。因此,mEAK-7 有可能通过调节 mTOR 信号通路而调节胰岛素分泌。
噬菌体展示技术成为探测蛋白空间结构、探索受体与配体之间相互作用结合位点、寻找高亲和力和生物活性的配体分子的有利工具,噬菌体展示技术目前已广泛应用于细胞生物学、食品、环境、农业、医药等领域[6-9]。本研究应用噬菌体展示技术最终筛选出的特异性最高的抗原表位“-GTFLFSLCAAVY”,且 ELISA 实验证明真核表达的 mEAK-7 蛋白能够与糖尿病血清中的靶分子有一定的特异结合效果。与目前广泛认可的糖尿病自身抗体GADA、IAA、ICA、ZnT-8A、IA-2A 阳性率对比,mEAK-7 在 I 型糖尿病中的阳性率接近于ICA、IAA、IA-2A 的阳性率,在 II 型糖尿病中的阳性率接近于 ICA、IA-2A 的阳性率,因此mEAK-7 对糖尿病自身抗体的检测具有一定的辅助作用,为深入研究糖尿病自身抗原抗体提供一定的实验依据。
[1] Kang HH,Liu JH,Wu LN, et al. Application and research progress of islet autoantibodies in diabetes. Lab Med Clin, 2018, 15(13):2015- 2020. (in Chinese)
康欢欢, 刘建华, 吴丽娜, 等. 胰岛自身抗体在糖尿病中的应用及研究进展. 检验医学与临床, 2018, 15(13):2015-2020.
[2] Wang TR, Bao Z. Study on combined detection of diabetes autoantibodies in diabetes typing. J North Pharm, 2015, 12(12):129- 130. (in Chinese)
王天荣, 鲍哲. 糖尿病分型中糖尿病自身抗体联合检测的探讨. 北方药学, 2015, 12(12):129-130.
[3] Harrington LS, Findlay GM, Gray A, et al. The TSC1-2 tumor suppressor controls insulin-PI3K signaling via regulation of IRS proteins. J Cell Biol, 2004, 166(2):213-223.
[4] Um SH, D'Alessio D, Thomas G. Nutrient overload, insulin resistance, and ribosomal protein S6 kinase 1, S6K1. Cell Metab, 2006, 3(6):393- 402.
[5] Nguyen JT, Ray C, Fox AL, et al. Mammalian EAK-7 activates alternative mTOR signaling to regulate cell proliferation and migration.
Sci Adv, 2018, 4(5):eaao5838.
[6] Shukra AM, Sridevi NV, Chandran D, et al. Production of recombinant antibodies using bacterio phages. Eur J Microbiol Immunol (Bp), 2014, 4(2):91-98.
[7] Shi LF, Wu Y, Li CY. Identification of high-affinity VEGFR3-binding peptides through a phage-displayed random peptide library. J Gynecol Oncol, 2015, 26(4):327-335.
[8] Zhao YY, Zhao SM, Liang Y, et al. Application of phage display technology in food safety. Sci Technol Food Ind, 2017, 38(14):342- 346. (in Chinese)
赵岩岩, 赵圣明, 梁颖, 等. 噬菌体展示技术在食品安全分析中的应用. 食品工业科技, 2017, 38(14):342-346.
[9] Hua XD, Shi HY, Wang MH. Phage display peptide library technology and its research progress in immunoassay of pesticide residue. J Food Saf Qual, 2014, 5(12):3955-3961. (in Chinese)
华修德, 施海燕, 王鸣华. 噬菌体展示肽库技术及其在农药残留免疫分析中的研究进展. 食品安全质量检测学报, 2014, 5(12):3955- 3961.
Screening of diabetes-associated antigen protein by phage display system and evaluation of its application effect
LIU Yao, LIN Jun, LIANG Yong-Yi, SHAN Yun, WANG Hong-tao, XU Liang
We aim to screen a novel epitope of antigen associated with diabetes by phage display system, andexpress new antigenic peptides and evaluate its potential application in the diagnosis of diabetes.
Type 1 diabetes patients serum were used for the panning of the Ph.D.-12 phage display library, and the selected phases were sequenced and the binding effect of the phage on the target molecule was detected by ELISA. The correlation between the best specific epitope and diabetes was analyzed by BLAST, and appropriate target gene was selected to construct recombinant plasmid. The new antigen peptide was expressed in Expi 293 cells and detected for specific binding effect on clinical serum.
A total of 2418 phage monoclonal sequences were sequenced, and 93 epitope sequences were selected, from which 4 of the 11 representative epitopes were sequenced more than 200 times. The best specific epitope sequence was identified as “-GTFLFSLCAAVY”. After analysis, mTOR-associated protein mEAK-7 was selected as a target gene, and the positive rate for the specific binding effect of the antigen peptide on clinical serum was 17% and 4% in type I and II diabetic serum, respectively.
mEAK-7 protein may have specific binding effect on diabetic serum. This gene may be related to diabetes and has good research value.
Phage display system; Diabetes; Autoantibody; Antigen epitope
XU Liang, Email: xuliang@szda.gov.cn
Author Affiliations:In-vitro Diagnostic Reagents Testing Department, Shenzhen Institute for Drug Control/Shenzhen Testing Center of Medical Devices, Gungdong 518057, China
10.3969/j.issn.1673-713X.2019.06.007
广东省食品药品监督管理局科技创新项目(2018ZDB14);深圳市科技计划项目(JCYJ20170307090049352)
518057 深圳市药品检验研究院/深圳市医疗器械检测中心体外诊断试剂部
徐良,Email:xuliang@szda.gov.cn
2019-09-09
猜你喜欢
传染病信息(2022年4期)2022-09-17
中国高原医学与生物学杂志(2022年3期)2022-06-22
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
智慧健康(2021年2期)2021-03-15
昆明医科大学学报(2021年1期)2021-02-07
科学24小时(2020年4期)2020-05-14
温州医科大学学报(2019年4期)2019-04-28
数码世界(2018年1期)2018-12-23
生物学教学(2018年10期)2018-11-29
