
氮素添加对贝加尔针茅草原土壤团聚体碳、氮和磷生态化学计量学特征的影响
2019-12-20 02:41:28李明秦洁红雨杨殿林周广帆王宇王丽娟
草业学报 2019年12期
李明,秦洁,红雨,杨殿林*,周广帆,王宇,王丽娟
(1.内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特 010022;2.农业农村部环境保护科研监测所,天津 300191;3.内蒙古呼伦贝尔市草原监督管理局,内蒙古 海拉尔 021008;4.内蒙古呼伦贝尔市草原研究所,内蒙古 海拉尔 021008)
氮沉降增加是全球气候变化的重要现象之一,已经对全球氮素循环产生了严重的影响,甚至可能对未来气候变化起到决定性的作用[1]。进几十年来,由于工农业的飞速发展,使得矿物燃料被大量开采燃烧以及大量农业化肥被生产使用,这些人类活动导致向大气中排放的含氮化合物与日俱增,大气氮沉降也呈迅猛增加的趋势[2]。在20世纪,大气氮沉降使得陆地生态系统氮素增加了3~5倍,且据有关研究显示预计到21世纪末全球氮沉降还将增加到现在的2.5倍[3-4]。中国是仅次于欧洲和美国的全球第三大氮沉降区[5],大量的氮素沉降对生态系统中的氮循环产生了极大的影响[6]。
草原生态系统是构成我国陆地生态系统的重要部分,约占我国国土面积的41%,远大于森林生态系统和农田生态系统[7-8]。草原生态系统具有提高土壤稳定性、维持生物多样性和促进物质循环的功能,同时还对维持全球二氧化碳平衡以及改善小气候等有着重要的贡献,但是草原生态系统也是最容易受人类活动和气候变化影响的陆地生态系统之一[9]。内蒙古呼伦贝尔草原是全球三大草原之一,是中国最美丽的、保存最完好的草原,作为我国北方重要的生态屏障,也是国家重要的畜牧业生产基地和畜产品主要产区[10]。草原生态系统能否健康发展直接影响着畜牧业的发展,草原既是畜牧业发展重要的生产资料,又承载着重要的生态功能[11]。贝加尔针茅(Stipabaicalensis)草甸草原是呼伦贝尔草原主要的地带性植物群落类型之一,在该地区植被群落中占据绝对优势[12]。土壤作为草原生态系统的核心,为人类的生存生活提供了必要的物质资源,是人类生活生产的物质基础,因此研究氮沉降增加对贝加尔针茅草原土壤生态系统的影响具有重要的意义[13]。
土壤团聚体是土壤颗粒通过有机物的胶结作用及动植物活动的团聚作用而形成的土壤结构的基本单元[14-15],土壤团聚体的组成与土壤的理化性质有着密切的联系,影响着植物的生长[16]。土壤有机碳作为土壤理化的重要指标,其组分特征影响着土壤质量和土壤肥力,并且对促进土壤团聚体的形成有着重要的作用,同时土壤团聚体的形成还可能是土壤固碳的过程[17-18]。
生态化学计量学结合了生态学和化学计量学等基本原理,是研究生态系统能量平衡和多重化学元素平衡的科学[19],其中碳(carbon)、氮(nitrogen)和磷(phosphorus)3种主要组成元素(以下简写为C、N和P)是生态化学计量学的主要研究内容[20]。研究土壤C、N和P的生态化学计量学特征对揭示土壤养分循环和平衡机制有着重要的价值[21]。我国学者对生态化学计量学的研究成果主要集中于植物方面的生态计量学特征,对土壤及土壤团聚体的生态化学计量学研究相对较少,而对于模拟氮沉降对土壤团聚体生态化学计量学的研究更是少之又少[22]。
基于此,本研究以内蒙古贝加尔针茅草原为研究对象,开展了为期8年的模拟氮沉降野外控制试验,研究氮沉降增加背景下贝加尔针茅草原土壤团聚体氮磷生态化学计量学特征,以期为全面分析和评估氮沉降对草原生态系统的影响提供基础资料。
1 材料与方法
1.1 试验区域概况及试验设计
试验样地位于内蒙古呼伦贝尔市伊敏苏木境内的贝加尔针茅草原(119°35′-119°41′ E、48°27′-48°35′ N),属半干旱大陆性季风气候。样地区域地势平坦,海拔760~770 m,年均降水量约330 mm,年平均气温-2.4~2.2 ℃,年平均无霜期100~120 d。研究区域植被覆盖密度高,贝加尔针茅在群落中占据绝对优势,此外羊草(Leymuschinensis)也是优势种。土壤类型为暗栗钙土。试验样地于2010年6月开始开展模拟氮沉降试验,采用裂区设计,共4个重复试验,每个重复试验设6个施氮水平,依次为: 0、15、30、50、100和150 kg N·hm-2·yr-1(不包括大气沉降的氮量),分别用N0、N15、N30、N50、N100和N150表示。重复间距设5 m的隔离带,小区面积8 m×8 m,各小区间隔2 m。氮肥选NH4NO3,用水溶解后均匀喷洒于各小区内[23-24]。
1.2 样品采集
于2018年8月上旬采集土壤样品,每个小区依“S”形采集10个点的原状土样轻微混合,采样深度为0~15 cm[24]。去除植物根系及其他土壤入侵物后,较大的土块沿着自然断裂面掰成直径约为1 cm的小块。并用硬塑料盒带回实验室,置于阴凉通风处风干,测定混合土壤样品的基本理化性质(表1)。
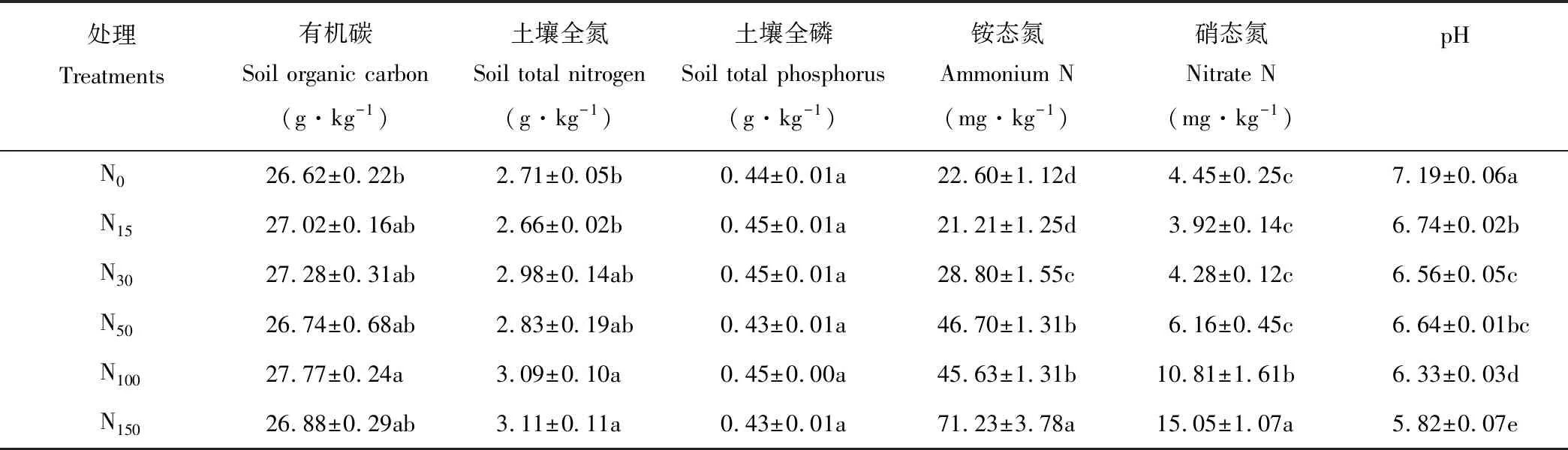
表1 氮素添加条件下土壤基本理化性质
注:同列不同小写字母表示处理之间差异显著(P<0.05)。下同。
Note: Different lowercase letters in the same column mean significant difference at 0.05 level among treatments. The same below.
1.3 测定方法
采用干筛法分离土壤团聚体[25],待土壤样品风干至含水量大约为8%时,先将较大的土块通过8 mm的标准筛轻轻破碎,并进一步去除石块和动植物残体。每次称取100 g风干土置于标准套筛中,垂直振动10 min,每分钟60次。分离出>2 mm,0.25~2 mm和<0.25 mm的3个粒级的土壤团聚体,其中>2 mm和0.25~2 mm(即>0.25 mm)粒径团聚体称为大团聚体,<0.25 mm粒径团聚体称为微团聚体。烘干后称量各个粒级团聚体的重量。取适量的团聚体研磨后过0.15 mm筛,用于土壤化学分析。另一部分未筛分的土样用于土壤常规分析。
土壤团聚体的稳定性用团聚体平均重量直径(mean weight diameter, MWD)来描述[26]:
式中:ri是第i个标准筛的孔径(mm),r0=r1,rn=rn+1,n为标准筛的数量,mi为第i个标准筛上团聚体所占百分比。采用水合热重铬酸钾氧化-比色法测定土壤有机碳;采用凯氏法,即硒粉-硫酸铜-硫酸钾-硫酸消煮法和流动分析仪(AA3,Bran+Luebbe Crop,德国)测定全氮;采用高氯酸-硫酸消煮-钼锑抗比色-紫外分光光度法测定全磷;采用氯化钙浸提法和流动分析仪测定土壤硝态氮、铵态氮;采用玻璃电极法(MP511 pH计)按土水比1.0∶2.5测定土壤pH[27]。
1.4 数据统计与分析
利用Excel 2013和SPSS 20.0对试验数据进行统计分析,不同氮添加处理土壤团聚体粒径分布的差异显著性采用单因素方差分析(One-way ANOVA),最小显著差异法(least significant difference, LSD)检验;通过双因素方差分析法(Two-way ANOVA)分析不同氮素添加处理、团聚体粒径及其交互作用对土壤化学计量学特征的影响,并用单因素方差分析(One-way ANOVA)和LSD法进行差异性检验。采用Excel 2013软件制作图表。
2 结果与分析
2.1 氮素添加对土壤团聚体粒径分布与稳定性的影响
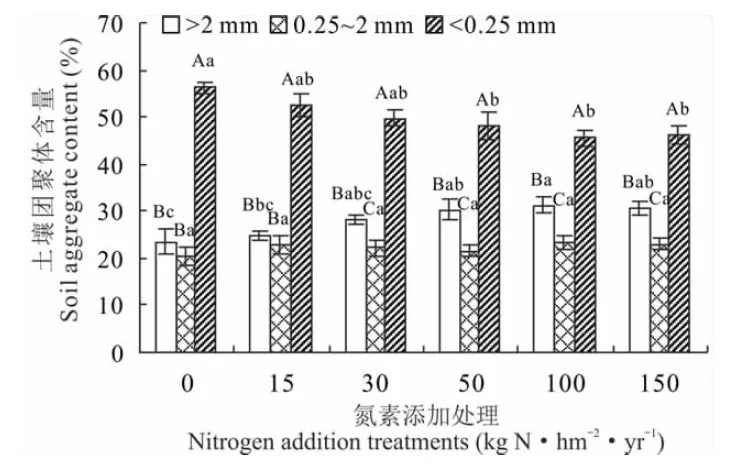
图1 不同氮添加处理土壤团聚体的粒径分布特征
2.1.1不同氮素添加水平土壤团聚体的粒径分布特征 不同氮添加处理对贝加尔针茅草原土壤团聚体组成的影响不同(图1),经过8年的模拟氮沉降试验,>2 mm的土壤团聚体含量随着氮添加量的增加先升高再下降,且在N50~N150处理时显著升高(P<0.05),其中N100处理极显著高于对照(P<0.01);0.25~2 mm的土壤团聚体含量随着氮添加量增加呈升高趋势,但各处理间差异不显著(P>0.05);<0.25 mm的土壤团聚体含量随着氮添加量的增加呈降低趋势,在N50~N150处理显著低于对照(P<0.05)。同一氮添加处理下土壤团聚体粒径分布均存在显著性差异(P<0.05),且<0.25 mm的土壤团聚体在土壤中占据大部分。
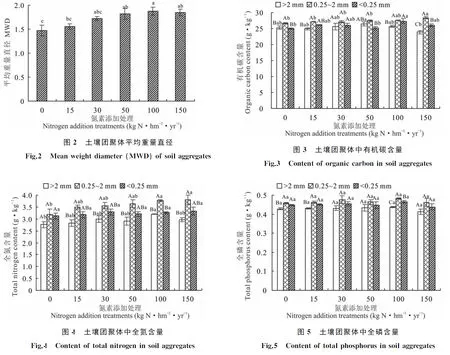
2.1.2不同氮素添加水平对土壤团聚体平均重量直径的影响 氮素添加促进了贝加尔针茅草原土壤团聚体平均重量直径的增加(图2),土壤团聚体平均重量直径在N100处理下较N0和N15处理显著增加,且与N0处理间存在极显著差异(P<0.01);N150处理较N100处理土壤团聚体平均重量直径减小,但处理间差异不显著(P>0.05)。
2.2 不同氮素添加处理土壤团聚体碳氮磷含量分布特征
2.2.1土壤团聚体有机碳含量 不同氮素添加处理贝加尔针茅草原的土壤团聚体有机碳含量变化范围为23.92~28.41 g·kg-1(图3),随氮素添加量增加有机碳含量呈增加的趋势,>2 mm的土壤团聚体在N50处理下有机碳含量达到最高,但与对照无显著差异(P>0.05),N150处理较N50处理显著降低了土壤团聚体的有机碳含量(P<0.05);0.25~2 mm土壤团聚体在N150处理下有机碳含量显著高于对照(P<0.05);<0.25 mm土壤团聚体在N100处理下有机碳含量最高,且显著高于对照(P<0.05)。
2.2.2土壤团聚体全氮含量 不同氮素添加处理贝加尔针茅草原土壤团聚体全氮含量变化范围为2.83~3.82 g·kg-1(图4)。N100处理显著提高了>2 mm和0.25~2 mm的土壤团聚体全氮含量(P<0.05);<0.25 mm的土壤团聚体全氮含量随着氮添加量增加也呈增加趋势,但处理间差异不显著(P>0.05);N150处理相较于N100处理降低了土壤团聚体全氮含量,但差异不显著(P>0.05)。
2.2.3土壤团聚体全磷含量 如图5所示,不同氮素添加处理贝加尔针茅草原土壤团聚体全磷含量变化范围为0.41~0.48 g·kg-1。各粒径土壤团聚体全磷含量随氮添加量增加的变化趋势与有机碳、全氮类似,但土壤团聚体全磷含量对氮添加处理的响应均未达到显著水平(P>0.05)。各粒径土壤团聚体在N100处理下全磷含量达到最高,N150处理下>2 mm、<0.25 mm土壤团聚体全磷含量与对照相比降低,但处理间差异不显著(P>0.05)。
同一氮素添加处理下土壤团聚体有机碳、全氮和全磷含量的分布均呈现中间高两边低的趋势,0.25~2 mm土壤团聚体的有机碳、全氮和全磷含量均高于其他粒径。除了N0、N50处理下>2 mm土壤团聚体有机碳含量高于<0.25 mm土壤团聚体外,其他处理<0.25 mm土壤团聚体有机碳、全氮和全磷含量均高于>2 mm土壤团聚体。
2.2.4土壤团聚体C、N和P 的相关性 对C、N和P 3种元素的相关性进行分析可知,不同氮添加处理各粒径土壤团聚体C、N和P之间呈极显著正相关关系(P<0.01)。C、N和P之间呈线性拟合关系(图6),且从斜率上看,P的变化相对于C和N的变化速度较慢,这可能是C和N是有机质结构性成分的原因。
2.3 不同氮素添加处理土壤团聚体碳氮磷生态化学计量特征
不同氮素添加处理土壤团聚体C/N变化范围为7.22~9.15,变异系数为6.9%(表2)。N100和N150处理下>2 mm土壤团聚体C/N与对照相比显著减小(P<0.05);不同氮添加处理土壤团聚体C/N在0.25~2 mm粒径中随着氮添加量的增加与对照相比呈减小趋势,N150处理较对照达到显著水平(P<0.05);<0.25 mm的团聚体C/N与对照相比无显著差异。同一氮素添加处理土壤团聚体各粒径C/N存在显著差异,除N100、N150处理外,>2 mm粒径土壤团聚体C/N均显著高于其他两个粒径(P<0.05)。

图6 土壤团聚体C、N和P 的相关性
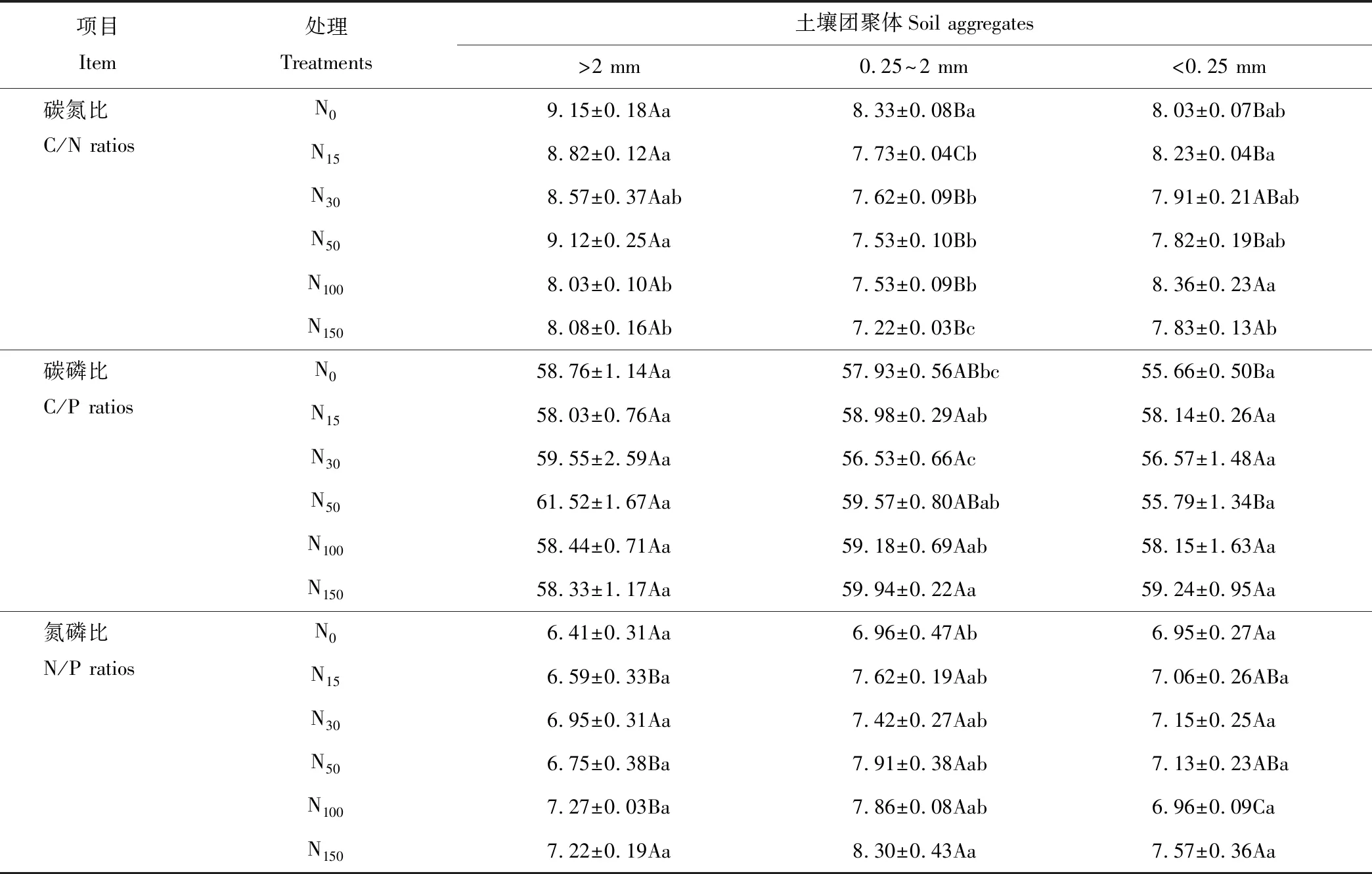
表2 不同氮素添加处理土壤团聚体碳、氮、磷化学计量学特征
注:同行不同大写字母表示团聚体粒径之间差异显著(P<0.05),同列不同小写字母表示团聚体处理之间差异显著(P<0.05)。
Note: The different uppercase letters in the same row mean significant differences at 0.05 level among aggregate sizes, the different lowercase letters in the same column mean significant differences at 0.05 level among treatment.
不同氮添加处理土壤团聚体C/P变化范围为55.66~61.52,变异系数为2.6%。不同氮添加处理,随氮素添加量增加>2 mm和<0.25 mm土壤团聚体C/P呈升高趋势,但差异均不显著;0.25~2 mm团聚体C/P随氮素添加量增加也呈增加趋势,并在N150处理下达到显著差异(P<0.05)。同一氮素添加处理,不同粒径团聚体C/P无显著差异(P>0.05)。
不同氮添加处理土壤团聚体N/P变化范围为6.41~8.30,变异系数为6.7%。不同氮添加处理随着氮添加量的增加,3个粒径的团聚体N/P均呈增加趋势,其中0.25~2 mm粒径团聚体显著增加(P<0.05),>2 mm和<0.25 mm团聚体增加差异不显著(P>0.05)。同一氮素添加处理下,0.25~2 mm团聚体N/P均高于>2 mm和<0.25 mm,N100处理使0.25~2 mm团聚体N/P显著高于其他粒径(P<0.05)。
氮素添加显著影响了土壤团聚体全氮、C/N和N/P(P<0.05),并对C/N有极显著的影响(P<0.01)(表3)。土壤团聚体粒径除对C/P有显著影响外,对有机碳、全氮、全磷、C/N和N/P均有极显著影响(P<0.01)。双因素方差分析显示,氮素添加和团聚体的交互作用显著影响了土壤团聚体有机碳(P<0.05),且对土壤团聚体C/N有极显著的影响(P<0.01)。

表 3 氮素添加和团聚体粒径对土壤团聚体生态化学计量特征影响的双因素方差分析
注:*在0.05水平上差异显著,**在0.01水平上差异极显著。
Note:*significant difference at the 0.05 level, **significant difference at the 0.01 level.
3 讨论
氮素添加可以使土壤生态系统的微环境发生改变,提高土壤肥力,同时也会使土壤pH降低[23]。本研究通过长期氮素添加处理,研究了氮素添加对贝加尔针茅草原土壤团聚体稳定性及生态化学计量学的影响,结果表明,氮素添加提高了土壤团聚体的稳定性及土壤肥力,同时氮素的添加也提高了土壤有机质的矿化速率。
3.1 氮素添加对土壤团聚体分布与稳定性的影响
团聚体的平均重量直径和>0.25 mm的土壤大团聚体含量是决定土壤抗侵蚀能力的两个重要指标,通常用来评价土壤团聚体的稳定性[28]。土壤团聚体的形成和稳定性与土壤中的有机质有着密切的联系,有机质是团聚体形成过程中重要的胶结物质[29]。适宜浓度的营养元素输入在一定的时期内对整个生态系统的发展是有利的[30]。郭虎波等[31]对杉木(Cunninghamialanceolata)人工林土壤团聚体的研究发现,中低氮处理可以提高土壤大团聚体含量以及团聚体平均重量直径,而高氮处理对土壤团聚体的含量与平均重量直径表现出一定的抑制作用。本研究结果显示,氮素添加处理提高了>0.25 mm土壤大团聚体含量以及团聚体平均重量直径。根据课题组前期研究显示氮素添加提高了地上植物的生产力,提高了凋落物及根系分泌物的量,从而促进了土壤中有机质含量,这可能是促进土壤大团聚体含量及团聚体平均重量直径增加的原因[32];土壤有机质含量的增加是促进土壤大团聚体形成的重要原因[31],刘晓东等[33]发现过高的氮元素添加会加速有机质的矿化,导致有机质含量下降,这可能是限制大团聚体形成的原因。
3.2 氮素添加对土壤团聚体碳氮磷含量的影响
土壤有机碳的含量是反应土壤肥力大小、衡量土壤质量的重要指标[32]。氮沉降对有机碳的影响存在很大的不确定性,有机碳含量主要决定于初级生产力对土壤的碳输入和微生物对土壤有机物质分解矿化作用引起的碳输出之间的平衡[34]。一般认为氮沉降促进土壤有机碳积累的主要原因是促进了植物的生长和凋落物积累从而增加了有机碳输入;氮沉降减少有机碳积累的原因主要是氮输入加剧了有机碳的分解矿化作用[34]。本研究结果显示,氮素添加显著提高了各粒径土壤团聚体的有机碳含量,但在N150处理时>2 mm粒径团聚体的有机碳含量较N50降低,这与郭虎波等[31]对杉木人工林土壤团聚体有机碳分布的研究结果相一致。同时也印证了Fornara等[35]的研究,即氮沉降增加对土壤有机碳的积累存在“碳饱和点”,土壤固定有机碳的能力在施氮量高于100 kg N·hm-2·yr-1处理下显著下降,阻碍了土壤有机碳的积累。
土壤团聚体全氮含量分布情况与有机碳有密切的联系,两者分布较为一致,这可能是由于土壤中的氮素95%以上都是有机态氮,主要为氨基酸、蛋白质、腐殖质等,和有机碳共同来源于植物凋落物或残体分解所形成的有机质[36-37]。氮添加后土壤有机质和全氮含量的升高主要是因为氮添加提高了草原植物的生产力,增加了凋落物和根系分泌物,进而丰富了土壤有机质[38]。随着氮素添加量的增加,大团聚体全氮含量显著增加,而微团聚体各处理间无显著差异,其原因是大团聚体在形成过程中结合了有机质,而微团聚体中有机质含量相对较低,导致微团聚体全氮含量随氮添加量的增加升高并不明显。
氮添加对土壤团聚体全磷含量变化影响较小,与李瑞瑞等[39]研究氮添加对墨西哥柏(Cupressuslusitanica)人工林土壤影响的结果相一致。Yang等[40]对落叶松(Larixspp.)人工林持续9年氮添加处理并未对土壤全磷含量产生显著影响。这是因为土壤磷素是一种沉积类的矿物,在土壤中的迁移率很低,土壤全磷含量主要受土壤母质的影响较大[32,36]。
在同一氮添加处理下,不同粒径团聚体有机碳、全氮和全磷的增加幅度也有所不同[41]。本研究结果显示,0.25~2 mm大团聚体中有机碳、全氮和全磷的含量高于>2 mm大团聚体和<0.25 mm微团聚体,这与Scott等[42]及Maysoon等[43]对农田土壤团聚体有机碳的研究结果相一致,可能与该粒径团聚体结合较多的轻组有机质有关[44],轻组有机质碳、氮含量高,周转快,易被土壤微生物分解,对提高土壤肥力具有重要的意义[45],所以0.25~2 mm粒径大团聚体最适合C、N和P积累,这可能是由于不同粒径的团聚体的物理性质和微生物特性的差异,造成大于该粒径的团聚体中的养分周转速度快,养分固定效率较低,小于该粒径的团聚体易分解的有机质含量低,造成总养分含量较低[46]。孔雨光等[44]在对土地利用方式变化对土壤团聚体影响的研究中也发现,各种土地利用方式下,均以0.25~2 mm大团聚体有机碳含量最高。有机质是团聚体形成过程中的主要胶结物质,有机分子与黏粒和阳离子相互胶结形成微团聚体,微团聚体再通过与周围基本粒子或微团聚体之间相互胶结形成大团聚体,同时在大团聚体内部由于颗粒有机质的分解,大团聚体解体形成微团聚体[44]。这可能是微团聚体有机碳含量低于大团聚体的原因。
3.3 氮素添加对土壤团聚体碳氮磷生态化学计量学特征的影响
土壤C/N是有机质或其他成分中碳素与氮素总质量的比值,是土壤有机质组成和质量的重要指标之一[36],通常被认为是土壤氮素矿化能力的标志[47]。土壤有机质和全氮是土壤质量中最为重要的指标,它们不仅反映土壤的肥力状况,也能印证区域生态系统的演变规律[48]。本研究中,氮添加处理下贝加尔针茅草原土壤团聚体C/N变化范围是7.22~9.15,低于中国陆地土壤C/N的平均值12.3[49]以及全球草地土壤C/N的平均值11.8[50],氮添加降低土壤大团聚体C/N。陈磊等[51]对黄土高原油松林(Pinustabulaeformis)土壤短期氮添加的研究结果发现土壤C/N随氮添加水平的增加而下降,本研究结果与其相一致。土壤氮素对土壤有机碳的调控有着重要的作用,较高的土壤氮素含量会加速土壤微生物对有机质的分解矿化,减少有机碳的积累,从而导致土壤C/N降低[51],这也是0.25~2 mm团聚体C/N比>2 mm团聚体C/N低的原因,由于0.25~2 mm团聚体中氮含量显著高于>2 mm团聚体,矿化速率高,导致C/N较低。
土壤C/P是衡量磷有效性的指标之一,C/P越低则磷的有效性越高[36]。此外,土壤C/P对植物的生长发育具有重要的影响[47]。氮添加处理下贝加尔针茅草原土壤团聚体C/P变化范围是55.66~61.52,高于中国陆地土壤C/P的平均值52.7,低于全球草地土壤C/P的平均值64.3[49-50]。氮添加提高了土壤大团聚体C/P,这与课题组前期研究结果一致,表明氮添加降低了土壤团聚体磷的有效性。
土壤N、P是植物生长所必需的矿质营养元素,也是生态系统中常见的限制性元素,土壤N/P在一定程度上间接预测群落养分的供给性水平和限制性水平[36]。氮添加处理下贝加尔针茅草原土壤团聚体N/P变化范围是6.41~8.30,高于中国陆地土壤N/P平均值3.9以及全球草地土壤N/P平均值5.6[49-50]。氮添加对草原各粒径土壤团聚体N/P均有所提高,表明氮添加减缓了草原植物氮限制,但是磷限制性增强[32]。相同氮添加处理,0.25~2 mm粒径土壤团聚体N/P最高,表明0.25~2 mm粒径大团聚体中养分限制类型以磷为主。
4 结论
氮素添加提高了草原土壤大团聚体的含量和团聚体的稳定性,对团聚体的稳定性和土壤有机碳的积累有着重要的意义,但高浓度的氮添加不利于有机碳的积累。氮添加提高了>0.25 mm土壤大团聚体有机碳、全氮的含量,其中以0.25~2 mm粒径团聚体最为显著,且在同一处理下,0.25~2 mm粒径团聚体有机碳、全氮、全磷含量最高,最适合养分的积累;氮添加未显著影响各粒径土壤团聚体全磷含量。氮添加导致团聚体C/N降低,C/P、N/P升高;提高了土壤团聚体有机质的矿化速率,随着氮添加量的增大,土壤团聚体中P元素成为限制草原植物生长的主要限制因素。氮素添加对贝加尔针茅草原土壤团聚体养分效应的影响、团聚体粒径分布和土壤生态化学计量学特征之间的内在联系有待更深入的研究。
猜你喜欢
云南化工(2021年5期)2021-12-21 07:41:34
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:32:44
中外文摘(2018年17期)2018-09-08 02:45:04
环球时报(2018-08-29)2018-08-29 04:10:59
现代农业(2016年3期)2016-04-14 12:35:28
创新科技(2015年9期)2015-12-15 07:04:09
土壤与作物(2015年3期)2015-12-08 00:47:01
云南中医学院学报(2014年4期)2014-07-31 18:22:23
郑州大学学报(理学版)(2012年4期)2012-03-25 13:59:10
对外经贸(2011年8期)2011-08-09 00:59:44
