
麦角菌的研究进展
2019-12-19 08:46张海娟何雅丽李秀璋李春杰
草业科学 2019年12期
张海娟,何雅丽,李秀璋,2,李春杰
(1. 兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 / 兰州大学草地农业教育部工程研究中心 / 兰州大学甘肃省西部草业技术创新中心 / 兰州大学草地农业科技学院,甘肃 兰州 730020;2. 青海大学畜牧兽医科学院,青海 西宁 810016)
广义上的麦角菌(Claviceps purpurea)包含所有有性和无性的近缘类群,被系统发育学归类为麦角菌科[1],该科包括常见的瘤座菌属(Balansia)、白僵菌属(Beauveria)、麦角菌属(Claviceps)、青茅斑穗病菌属(Ephelis)、香柱菌属(Epichloe)、亚肉座菌属(Hypocrella)、绿僵菌属(Metarhizium),还包含一些罕为人知的属,例如Hyperdermium、Munkia、Myriogenospora、 Paecilomyces、 Tolypocladium和Ustilaginoidea等。狭义的麦角菌仅指麦角菌属(Claviceps)真菌,由大约60 种专门寄生禾草子房部位的真菌组成[2]。由于该属真菌主要寄生在黑麦(Secale cereale)、 小 麦(Triticum aestivum)、 大 麦(Hordeum vulgare)、燕麦(Avena sativa)、披碱草属(Elymus)和鹅观草属(Roegneria)等禾本科植物的子房内,菌丝体充满子房使之成为菌核,形状如同麦粒,故称之为“麦角”。本文所指“麦角菌”仅为麦角菌属真菌。
麦角菌与人类有着长期密切的关系。一方面,麦角菌产生的生物碱在医学上得到了广泛的应用;另一方面,麦角菌产生的毒素,给草原畜牧业生产造成巨大的损失。最近有关人类麦角中毒的报道发生在20世纪70 年代的印度、1978 年和2001 年的埃塞俄比亚[3]。正是由于麦角菌的双重特性,引起了国内外相关科研工作者的广泛关注,并开展了大量的研究,近年来逐渐成为了该领域重要的研究热点之一,因此很有必要对麦角菌的研究进展加以综述。
1 麦角菌的发展历史及其生物学特征
1.1 麦角菌的发展历史
自从欧洲-地中海农业开始以来,麦角菌很可能就与人类一起存在(6 000-2 000 B.C.)。然而,第一次记录在案的由麦角菌引起的人类毒性传播发生在公元857 年的德国。随后在欧洲的许多地方发生了其他流行病,给人类带来重大损失[4]。人类麦角中毒主要是由于食用含麦角的粮食产品所致,麦角中毒的症状主要有两类[5],即坏疽性麦角中毒和痉挛型麦角中毒。坏疽性麦角中毒是由于麦角生物碱的血管收缩特性导致,这种特性会减少血液流向手和脚,由于血液流动不足而腐烂,严重时可导致断肢。痉挛性麦角中毒的症状是神经失调,主要包括麻木、抽搐、运动失协、呼吸困难、脉搏加快、流涎、呕吐、失明、瘫痪和痉挛等症状,此外,产生幻觉和痴呆也是麦角中毒症状的一部分[6]。
历史上的人类流行病,例如黑瘟病。有学者认为是由于长期存在于面包中的麦角生物碱和其他霉菌毒素导致人类抵抗力减弱,从而削弱免疫系统所致。1350 年至1490 年期间,欧洲人口的减少也被认为是由于人类食用了麦角感染的黑麦面包而导致女性生育率下降[7]。
目前,世界大部分地区已不再受麦角中毒的影响。但近年来,家畜麦角中毒仍然时有报道,重新引起了科学界的关注[8]。喂食麦角侵染的谷物会导致家畜缺血性肢端坏死、饲料摄入量减少、生长和生殖障碍等[9]。
早在19 世纪和20 世纪初,Atanasoff 就对麦角菌的生物学、多样性和分类地位进行了研究总结。20 世纪中期,Giles 和Langdon[10]在澳大利亚和非洲共发现并描述了26 种麦角菌,对麦角菌的分类学做出了重大贡献。后来,Loveless 等研究了津巴布韦的麦角菌,描述了几个新种,并强调了分生孢子状态和宿主植物在麦角菌鉴定中的作用[11-12]。在日本,Tanda 于1977 年至1992 年对麦角菌进行了报道,共描述了5 个新种[13]。此外,Pazoutová等[14-15]描述了来自美洲和非洲的15 种麦角菌,并对较为罕见的几种麦角菌进行了深入研究。
Tulasne[16]第一次明确了麦角菌的所有生活史阶段都属于同一种真菌,并以麦角菌的名称对它的有性阶段进行了描述。与此同时,有80 多种麦角菌属真菌和超过10 种小头孢菌属真菌(Cephalosporium)[17]被归类到麦角菌属,而且对其形态特征进行了详细描述。然而,后期研究发现这些真菌中仅有59 种属于麦角菌属真菌,其余被划分到其他真菌类群。近年来,随着国际命名规则的变化,每种真菌只允许有一个科学名称,因此作为有性态麦角菌的蜜孢霉属(Sphacelia)和无性态麦角菌属的取舍成为了大家讨论的焦点。尽管蜜孢霉属的命名相对较为古老,但由于麦角菌属这个名称被广泛使用且受到社会和众多研究者的高度重视,最终这一名称得以保留[18]。
1.2 麦角菌的生物学特性
麦角菌属属于子囊菌亚门麦角菌科,此属的大部分物种均可引起动物中毒,例如雀稗麦角菌(Claviceps paspali)的麦角可引致畜禽中毒。早期关于麦角菌生物学特征的描述主要通过其侵染宿主种子形成的菌核[19-20]和无性繁殖形成的分生孢子进行描述[21]。“蜜露”内无性世代在寄主子房内的菌丝垫中形成不规则腔室,产生分生孢子,分生孢子卵形至椭圆形,单胞,无色,(3.5~6) μm ×(2.5~3) μm[21]。病原菌的菌核呈香蕉形、柱状,表层紫黑色,内部白色,质地坚硬,大小因寄主而异,一个病穗可产生几个或几十个麦角[19-20]。麦角成熟后落入土壤中越冬。翌年条件适宜时萌发出1~50 个肉色有细长柄的子座。子座直径1~2 mm,球形,肉红色,外缘生许多子囊壳;子座柄白色,长5~25 mm;上有许多突起,即子囊壳的孔口,子囊壳埋生于子座表皮组织内,烧瓶状,内有若干个细长棒状的子囊,子囊壳大小 (150~175) μm ×(200~250) μm。子囊透明无色,细长棒状,稍弯曲,大小为4 μm × (100~125) μm,有侧丝,子囊内含8 个丝状孢子,后期有分隔,大小(0.6~0.7) μm ×(50~76) μm。子座产生5~7 d 后子囊壳成熟,空气相对湿度为76%~78%时,子囊孢子可以发射出来,随粘性物质排出。发射出的子囊孢子借气流传播,粘液中的分生孢子借飞溅的水滴和昆虫传播到其他小花上[22]。
2 麦角菌的遗传进化和分布
2.1 麦角菌在进化过程中的生物地理学分布
Píchová等[2]提出,麦角菌属、竹针孢座囊菌属(Aciculosporium)和珊座孢新属(Corallocytostroma)的共同祖先起源于南美洲-南极洲-澳大利亚地区的上白垩纪,它们共同形成了一个紧凑的区块。巴塔哥尼亚从上白垩纪到晚古新世连接到南极半岛[23],南极洲-澳大利亚的分裂发生在4 500 万年前[24]。这次分裂对应与植物相关的麦角菌科(Balansia、Myriogenospora、Atkinsonella 和Periglandula)真菌的当前分布。麦角菌始于古新世,它的原始宿主可能是早熟禾亚科(BEP)和PACMAD[黍亚科(Panicoideae)、三芒草亚科(Aristidoideae)、虎尾草亚科(Chloridoideae)、百生草亚科(Micrairoideae)、芦竹亚科(Arundinoideae)和扁芒草亚科(Danthonioideae)]的共同祖先。
基础物种Claviceps citrina可能起源于南美洲,但目前仅在墨西哥有报道[25]。虽然其唯一已知的宿主海滨盐草(Distichlis spicata)在南美洲也有分布[26],但是无法确定南美洲是否存在C. citrina。由于缺乏南美洲麦角菌属的最新记录,阻碍了该属进化和分布的重建。此外,仅在南美洲发现C. flavella、C. diadema和C. orthocladae,其菌核未分化,认为这是一种原始特征[27-28],这进一步支持了南美洲起源的理论。有学者研究雀稗麦角菌(C. paspali)原产于南美洲[29],其传播依赖于寄主,雀稗属(Paspalum)是新世界的一个大属,起源中心在南美洲,物种多样性大[30]。麦角菌是BEP 枝中唯一发生在禾本科植物上的物种,麦角菌的寄主可能是莎草(Cyperus rotundus)。莎草科起源于上白垩世的南美洲[31],促进了麦角菌由原始寄主向莎草转移。这种真菌与新宿主一起传播到北美洲,在中新世与早熟禾亚科接触后分枝。早熟禾亚科在其祖先从非洲迁移到亚欧大陆后起源于亚欧大陆,随后通过北大西洋传到北美洲[32]。C. cyperi进化枝仍与莎草科保存在一起,并随莎草来到亚欧大陆和非洲。剩下的两个分枝都转移到了早熟禾亚科,后来又有几个物种转移到了稻亚科或PACMAD 禾草。早熟禾亚科的另一种共生体,在渐新世(2 800 万年前)开始分枝,时间较长,未发生任何分枝事件。这与核心区蒲亚科是一致的。核心早熟禾亚科的冠节点被认为在2 500 万年到3 200 万年前[32-34],在始新世-渐新世边界经历全球变冷后(2 600 万年到3 300 万年前)出现了应激反应基因家族的扩增,使其能够适应寒冷的气候,并向较冷的栖息地传播[35]。麦角菌与其宿主一起从封闭的栖息地转移到了气候稳定,开放、凉爽的栖息地,其菌核变黑和拉长。麦角菌的进化很明显依赖了早熟禾亚科宿主的进化。麦角菌的寄主谱很广,覆盖了许多禾本科植物亚科。虽然主要宿主是早熟禾亚科,但是几种物种,例如C. arundinis,C. humidiphila,C. macroura和C. spartinae,可 以 以PACMAD 作 为宿主。假设这些物种通过寄主转移在其区域中发生宿主移位,而C. arundinis和C. humidiphila可以感染芦竹亚科和早熟禾亚科(BEP)。麦角菌保留了感染莎草的能力,在苔草属(Carex)上感染麦角菌[36]。
C. pusilla及其宿主[32](PACMAD)的起源估计分别在非洲5 000 万年以前和4 500 万年以前,麦角菌从南美洲向非洲转移。当时非洲和南美洲是分开的,因为这两个大陆的分裂发生在一亿三百万年至一亿一千三百万年前[37]。然而,有学者认为新旧世界之间是分散的。这一假设得到了一些植物或昆虫类群例子的支持,这些类群的进化枝太年轻,它们的分布没有受到冈瓦纳碎片的影响。因此,如果长距离传播(long distance distribution, LDD)发生在始新世期间,则假定是在大西洋或通过陆地热带路径的长距离扩散[37-40]。麦角菌可能与宿主一起或通过昆虫媒介传播到旧世界,后者被认为更有可能。昆虫在麦角菌向健康宿主植物或新地区的传播过程中发挥着重要作用,它们以含大量麦角菌的含糖甘露为食[41]。蜜露吸引蝇科(Muscidae)[42-44]、 夜 蛾 科 (Noctuidae)、 蚜 总 科(Aphidoidea)、 叶 蝉 科(Cicadellidae)、 蓟 马 亚 科(Thripinae)[42]和臭虫科(Cimicidae)[45]。昆虫可能在古新世晚期至始新世中期通过跨大西洋长距离传播将麦角菌的分生孢子转移到非洲。在非洲,这种真菌开始侵染新出现的PACMAD,后来主要侵染黍亚科。黍亚科在中新世早期从非洲传播到北美洲[32]。C. etariiphila来自巴西,但可能起源于非洲。它的宿主是幽狗尾草(Setaria parviflora),发现在新世界,但起源于旧世界[46]。据推测,C.setariiphila可能与小花鬼针草(Bidens parviflora)一起引入北美。由于C. tenuispora在非洲的狼尾草属(Pennisetum)[47]和蒺藜草属(Cenchrus)[48]上有所发现,所以,尽管有来自南美洲和北美洲的历史记录,比如姊妹种C. setariicola是一种美国麦角菌[49]。但仍有学者认为C. tenuispora原产于非洲。虽然Píchová等[2]的分析表明麦角菌起源于南美洲,但最近没有发现麦角菌属的南美洲物种,因此无法确定。
在我国,1984 年刘若和马振宇对甘肃中部榆中县贡井乡、中国农科院兰州畜牧研究所实验基地附近的野生禾草的麦角病进了调查[50]。2003 年,李春杰对醉马草麦角病的病原(Sphaceliasp.)及发病症状进行了描述[51]。2008 年,孙相辉对芨芨草(Achnatherum splendens)、醉马草(Achnatherum inebrians)、披碱草(Elymus dahuricus)等7 种禾草麦角病的发生及其病原真菌的特征进行了描述[52]。国内对麦角菌的相关研究相对较少,因此很有必要对麦角菌的起源进行更深一步的研究。
2.2 现代麦角菌在全球的分布
麦角病在世界各地均有分布,在气候比较冷凉潮湿的地区发生尤为严重。麦角病主要分布在潮湿的温带地区,加拿大和北欧一些国家生产的禾本科牧草种子中含有较多的麦角(即麦角菌的菌核)[2]。全球分布主要为6 个区域:非洲、南美洲、欧亚大陆、北美洲、澳大利亚和印度[46]。在我国几乎到处都有分布,南至贵州,北达黑龙江,东自浙江,西抵青海,但正式报道的以北方各省(区)为主,其中又以新疆受害草种为最多[22]。
3 麦角菌的寄主范围和麦角病
3.1 寄主范围
麦角菌可以侵染7 个科或亚科70 多属的400余种禾本科植物,分别为黍亚科(Panicoideae)、画眉草亚科(Chloridoideae)、芦竹亚科(Arundinoideae)、早熟禾亚科(Pooideae)、竹亚科(Bambusoideae)、稻亚科(Ehrhartoideae)和莎草科(Cyperaceae)。其中为畜牧业带来巨大经济收益的属有冰草属(Agropyron)、剪股颖属(Agrostis)、看麦娘属(Alopecurus)、燕麦属(Avena)、野牛草属(Buchloe)、雀麦属(Bromus)、拂子茅属(Calamagrostis)、鸭茅属(Dactylis)、披碱草属(Elymus)、羊茅属(Festuca)、洽草属(Koeleria)、猬 草 属(Hystrix)、 黑 麦 草 属(Lolium)、 虉 草 属(Phalaris)、黍属(Panicum)、猫尾草属(Synthyris)、芦苇属(Phragmites)、甜茅属(Glyceria)、绒毛草属(Holcus)、早熟禾属(Poa)、碱茅属(Puccinellia)、狗尾草属(Setaria)、黑麦属(Secale)、针茅属(Stipa)、小麦属(Triticum)和结缕草属(Zoysia)等[2]。
3.2 麦角病的发生及其防治
麦角菌侵染寄主引起的病害称之为麦角病,是草地早熟禾(Poa annua)上的一种重要的花絮部病害[53-54]。麦角病菌主要寄生在粮食作物及禾草的穗头上,当冬天来临时,菌核落入土中过冬。麦角菌侵染在开花期,菌核萌发,单细胞的子囊孢子借助风力传播到寄主的花穗上,立刻萌发出芽管,由雌蕊柱头侵入子房。菌丝滋长蔓延,发育成白色、棉絮状的菌丝体并充满子房。破坏子房内部组织后逐渐突破子房壁,生出成对短小的分生孢子。同时菌丝体分泌一种具有甜味的粘性物质,引诱苍蝇、蚂蚁和蚜虫等昆虫把分生孢子传至其他健康的花穗上,麦角病随之重复传播。当禾草种子快成熟时,受害子房不再产生分生孢子,子房内部的菌丝体逐渐收缩成一团,进而变成黑色坚硬的菌丝组织体成为菌核[55]。麦角病菌侵染植物子房过程和植物花粉受精的形式相似。一般情况下,植物花粉受精只需要几个小时,而麦角病菌侵染子房需要一天的时间,麦角菌的感染只能在子房受精前进行,受精后的子房不会受麦角病菌的侵染[56]。
在麦角病的防治方面,抗病育种已被证明是主要的防治方法。在谷物中,对子房感染的抗性一般在受精后的几天内产生。由于花是植物唯一易受感染的器官,因此需要花期短的品种[57]。适当使用杀菌剂可以降低麦角菌侵染的严重程度。为了达到最佳效果,必须在开花初期施用杀菌剂。虽然杀菌剂可以减轻麦角菌侵染的严重程度,但目前还不清楚杀菌剂的应用是否具有经济效益。Pérez 等[58]研究表明,施用土壤杀菌剂可以减少麦角菌菌核的萌发。加强田间管理在麦角病的防治过程中有重要的作用,由于菌核是侵染的主要来源,在收获期尽可能多地摘除菌核可减少来年春季潜在的孢子数量。也可通过露天焚烧麦角来减少初侵染来源。
昆虫,尤其是苍蝇和飞蛾以麦角蜜露作为食物来源。当这些昆虫在种子周围爬行时,它们可以将蜜露转移到其他花朵上,从而使麦角病传播[42]。然而,关于昆虫在麦角病流行病学中所起作用的定量资料还很缺乏。目前还不清楚昆虫是如何有效地进行麦角病的二次传播以及控制昆虫是否会减少麦角病的二次传播。
4 麦角菌次生代谢产物
4.1 麦角菌次生代谢产物的多样性
麦角类生物碱是麦角菌的主要次生代谢产物。目前,已从麦角菌中分离出了70 多种生物碱,根据C8 位取代基结构的不同可将麦角碱分为 4 类(表1):棒麦角生物碱、简单麦角酸衍生物、肽型生物碱及酰胺类麦角生物碱[54,59-60]。许多麦角菌科物种缺乏生物碱,对于一些麦角菌属个体也是如此[60],表明麦角生物碱(ergot alkaloids, EA)不是病原菌致病所必需的。
值得注意的是,Langdon 等[61]研究发现,麦角菌属部分菌核总是色素沉着,有几组色素,如黑色素和类胡萝卜素,可增强真菌的抗逆性(例如抗UV 辐射或氧化应激)。麦角菌(C.purpurea)含有结构多样的紫色色素、红色色素和黄色色素,色素在菌核胁迫耐受中的作用还有待研究。
麦角生物碱EAs 对细菌、真菌[62]、昆虫和食草动物都有毒性,特别是clavines 和ergopeptines 中含量丰富的黑麦酮酸(secalonic acid) A、B 和C,菌核中高水平的黑麦酮酸会严重影响哺乳动物细胞代谢能力。Wäli 等[63]研究表明,麦角菌侵染会导致寄主植物种子减少,但是麦角菌能通过预防食草动物采食而增强寄主对环境的适应性。事实上,麦角菌与寄主的相互作用是一种有条件的防御互惠关系。C.spartinae的存在增强了寄主对环境的适应性,因为补偿作用受侵染小穗(小于10%)的花序比未受侵染的花序有更多的种子数量,尽管种子重量较低[64]。部分麦角菌种类,如非洲C.africana、 C. cynodontis、 C.fusiformis、 C. gigantea和C. paspali,通常具有高水平的麦角生物碱(EAs)(0.1~10 mg·g-1)[65-68],产生生物碱的区域更适合建立保护性互惠共生体。
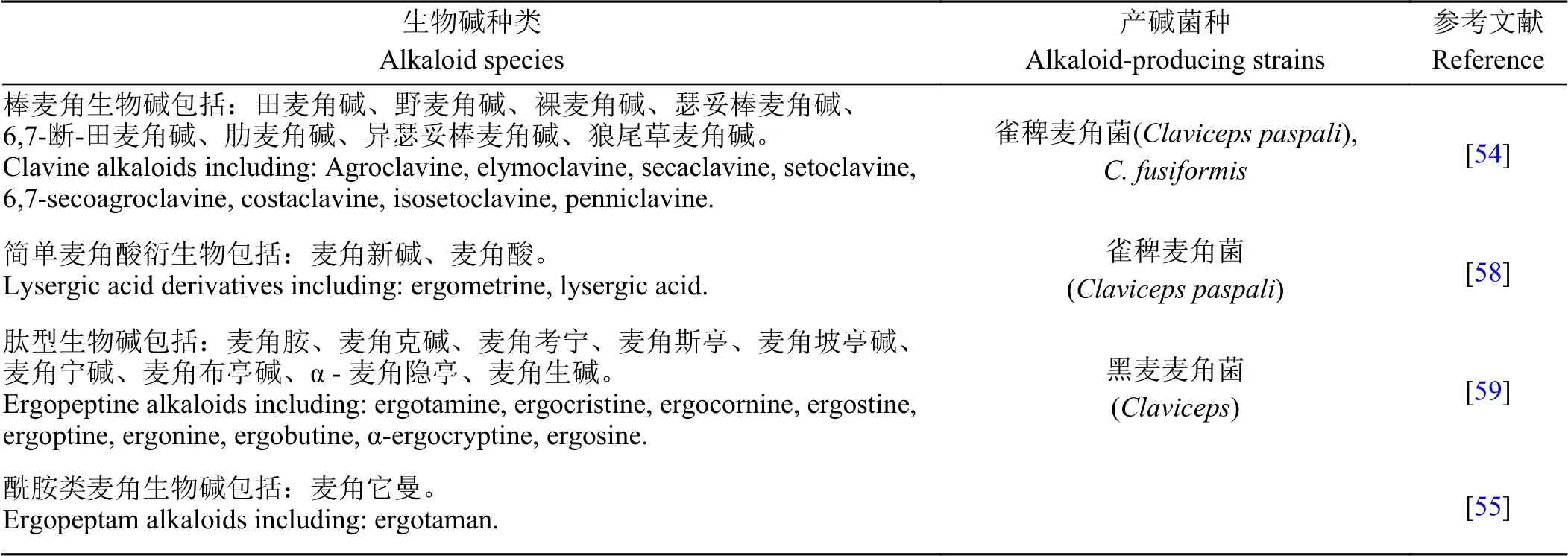
表 1 常见的麦角生物碱Table 1 Common ergot alkaloids
4.2 麦角菌次生代谢产物生物碱在医学上的应用
麦角生物碱是已知最早的天然药用产物之一。由于该类生物碱具有多种生物活性,在药理学和医学上得到了广泛的应用。欧洲助产士用麦角来加速分娩,少数菌核的使用常常使分娩时间缩短几个小时[69]。麦角胺及其衍生物用于治疗偏头痛,溴麦角环肽用于帕金森病和Ⅱ型糖尿病[70-72]。麦角溴烟酯适用于治疗各种血管疾病如脑血栓和动脉粥样硬化,动脉阻塞[73-74]。在目前使用的半合成生物碱中,卡麦角碱和尼可胆碱被广泛应用于各种适应症,包括高泌乳素血症、帕金森病和衰老[75]。在转移性乳腺癌和前列腺癌的治疗中,通常使用低剂量的溴隐亭与经典的抗肿瘤疗法一起[76]。海胆碱有改善记忆和大脑功能的效果[77]。人们还发现了许多其他的化学物质,其中大部分来自麦角菌,这使该物种成为在次生代谢物生产方面探索的最有价值的真菌之一。
5 展望
广义上的麦角菌指麦角菌科真菌,狭义上的麦角菌仅指麦角菌属真菌。本文只综述了狭义上麦角菌的生物学特性、遗传进化、麦角病以及麦角生物碱在医学上的应用。目前,在抗麦角病牧草的培育、麦角病杀菌剂的筛选、麦角菌与内生真菌互作关系的研究方面还很缺乏。
首先,家畜采食受麦角菌侵染的牧草会引起中毒,给畜牧业带来了巨大的经济损失,因此,很有必要培育出抗麦角病的牧草新品种。其次,由于麦角菌能产生多种有生物活性的麦角生物碱,同时其衍生物可以治疗衰老和痴呆。因此,很有必要筛选出高产碱量的菌株,开发新药剂,为其在医学上的应用提供药物来源。此外,尽管在作物开花期喷施广谱性杀菌剂可以一定程度上防治麦角病[6],但是开发低毒高效且环境友好型生物源杀菌剂迫在眉睫。
笔者调查发现禾草-内生真菌共生体上常伴有麦角菌的侵染,且禾草内生真菌亦隶属于麦角菌科、也可产生麦角类生物碱。因此,对比分析这两类菌的进化关系和麦角生物碱合成途径,明确二者互作对宿主禾草生长的影响,揭示它们对宿主禾草生长的影响机制,探讨麦角菌、禾草内生真菌和宿主禾草之间的互作机制。
猜你喜欢
食品与发酵工业(2022年24期)2022-12-29
第二课堂(课外活动版)(2022年5期)2022-05-25
中国典型病例大全(2022年10期)2022-05-10
西北农业学报(2021年5期)2021-06-28
世界最新医学信息文摘(2021年12期)2021-06-09
中国食用菌(2021年1期)2021-03-12
武夷科学(2019年2期)2019-12-20
中国食用菌(2019年9期)2019-10-21
中成药(2018年3期)2018-05-07
中国当代医药(2015年36期)2015-03-11
