
宁夏东部荒漠草原灌丛引入过程中土壤呼吸响应特征
2019-12-19 08:46郭天斗赵亚楠周玉蓉王红梅
草业科学 2019年12期
郭天斗,赵亚楠,周玉蓉,王红梅,
(1. 宁夏大学农学院,宁夏 银川 750021;2. 西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021)
草原灌丛化是当前干旱、半干旱地区面临的重要生态问题[1]。多年来,干旱、过度放牧、火烧、温室气体增加及降水格局变化等因素都会导致草原优势植被的变化以及水分、养分的空间异质性[2-3],进而破坏草地生态系统的稳定,改变草地生产力及生物多样性[4]。土壤呼吸作为陆地生态系统与大气交换碳素的主要环节,连接着植物-土壤-微生物之间的碳转化过程,对生态系统碳循环至关重要[5]。全球的碳收支过程对气候变化十分敏感,土壤呼吸是碳素损失的主要途径之一,而荒漠草原人工引入旱生灌木过程中,土壤干旱化和斑块化加剧,也可能会影响生态系统碳循环[6]。
自然降水是荒漠草原唯一的水分来源,因此水分是该地区土壤呼吸的主要影响因子[7],土壤呼吸随土壤温度和水分变化呈现季节变化,且凋落物和土壤微生物活动对土壤呼吸影响较大[8]。荒漠草原人工灌丛引入后,植被净光合效率和生长速率都升高[9],土壤呼吸对土壤湿度和温度的敏感性增加[10],会加速灌丛化进程。Felton 等[11]研究了不同降水对草地植物净初级生产力和土壤呼吸的响应,表明净初级生产力与降水呈线性关系,并且水分对土壤呼吸影响很大。已有学者对全球多年土壤呼吸数据进行收集并分析其时空变化,研究发现,温度、降水量和土壤有机碳是土壤呼吸的关键控制因素[12]。Brye 等[13]指出,气候变化会影响土壤有机质分解和土壤碳循环,进而导致碳储量的差异。在全球气候变化的背景下,国内外有关荒漠草原土壤呼吸响应特征的研究也成为干旱区生态学热点问题,主要集中在不同植被类型土壤呼吸特征的研究[10,14-15],而荒漠草原人工灌丛引入过程土壤呼吸特征研究相对较少。宁夏荒漠草原大面积人工引入柠条灌丛(Caragana korshinskii)进行植被恢复,引起植被多样性下降、深层土壤水分消耗以及斑块间土壤侵蚀加剧的现象[16]。虽有研究表明,“沃岛效应”会导致灌丛斑块的土壤碳氮和持水能力增强,但灌丛的引入也增加了荒漠草原水土资源的空间异质性[17],因此,荒漠草原灌丛引入下土壤特性的变化仍需进一步研究。
基于此,本研究以宁夏荒漠草原不同种植年限、间距人工柠条灌丛地为研究对象,分析灌丛引入过程中土壤呼吸与土壤水分、土壤碳氮以及土壤微生物的变化,旨在了解和掌握在降水有限的荒漠草原灌丛引入过程的土壤呼吸响应特征,以期对荒漠草原碳循环研究提供基础数据,进而为荒漠草原人工植被恢复的可持续性提供科学依据和数据支撑。
1 材料与方法
1.1 研究区概况
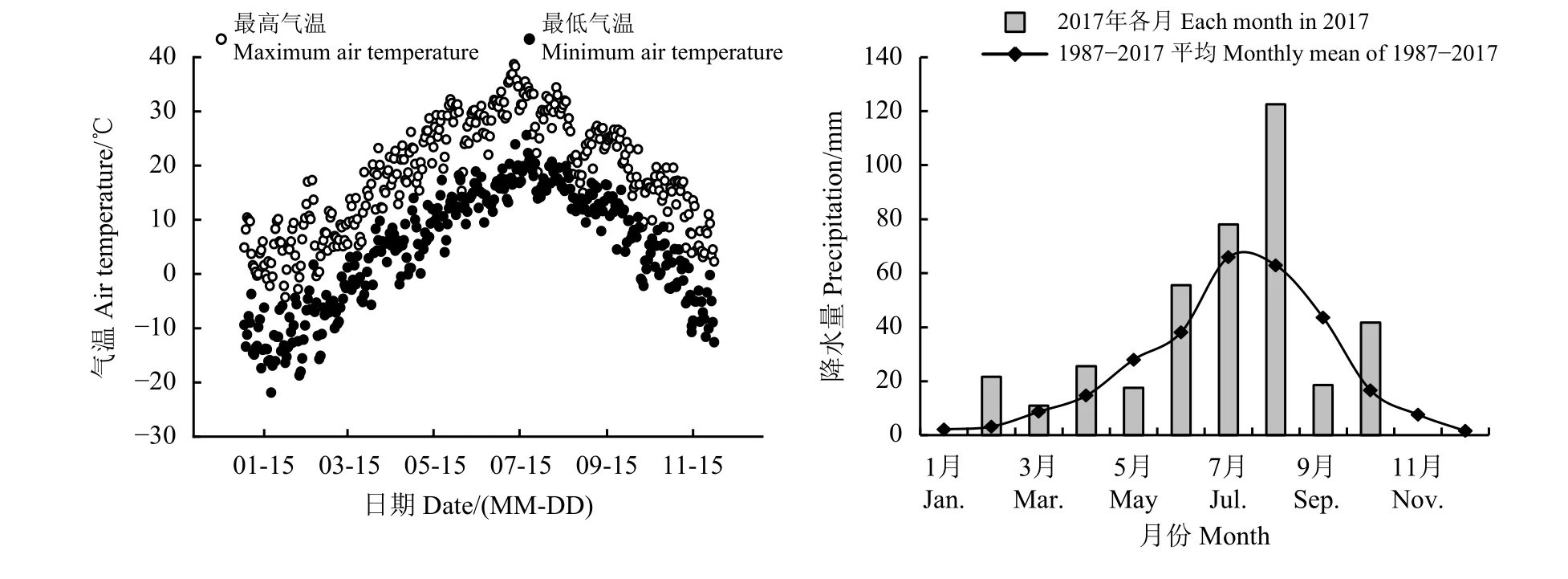
图 1 盐池县2017 年日气温变化与1987-2017 年月均及当年各月降水量分布Figure 1 The change of daily temperature in 2017 and the distribution patterns of monthly precipitation from 1987 to 2017 and every month in 2017 in Yanchi County
研究区位于宁夏回族自治区东部盐池县花马池镇(37°04′-38°10′ N,106°03′-107°04′ E)。该区北接毛乌素沙漠,南靠黄土高原,四季多风少雨,干燥炎热,属于典型的温带大陆性季风气候,年平均气温8.1 ℃,年平均温差28 ℃,平均海拔1 450 m,多年平均降水量300 mm 左右,降水主要集中在7 月- 9 月[14](图1)。该区是由半干旱向干旱区过渡的荒漠草原,其土壤类型以灰钙土为主,土质疏松,土壤肥力低,补水能力差,易受风蚀和沙化。植被类型以典型的沙生植物为主,主要有赖草(Leymus secalimus)、猪毛蒿(Artemisia scoparia)、白 草(Pennisetum centrasiaticum)、柠 条(Caragana korshinskii)、骆 驼 蓬(Peganum harmala)、针 茅(Stipa capillata)、 虫 实(Corispermum hyssopi folium)、牛枝子(Lespedeza potaninii)和刺蓬(Salsola ruthenica)等。
1.2 样地设置
在盐池县东北部荒漠草原区选取封育草地(enclosed grassland,EG)、放牧地(grazing grassland,GG)、不 同 种 植 年 限[3 年(SY3)、12 年(SY12)、22 年(SY22)]和年限相同不同间距[40 m (SI40)、6 m (SI6)、2 m (SI2)]的柠条灌丛地(其中SY22 和SI6为同一样地),利用空间梯度代替时间梯度的方法来代表荒漠草原退化及人工灌丛引入恢复的不同阶段和过程,即草原-放牧-灌丛引入。样地基本情况如表1 所列。
1.3 观测指标及方法
1.3.1 土壤呼吸观测
土壤呼吸速率测定采用动态密闭气室分析法,利用EGM-4 便携式环境气体检测仪观测,呼吸罩面积78 cm2。在7 个样地,距灌丛1.5 m 的范围内按间隔30 cm 的距离随机选取5 点观测,分别测定各点土壤呼吸强度及浓度,于每月中旬观测一次(生长季一月两次),单次观测时间为120 s,在封育草地及放牧草地进行正常间距随机观测。
1.3.2 土壤水分监测
土壤水分的观测时间在2017 年3 月-11 月,主要利用德国产TDR 剖面土壤水分测量系统定点长期观测,EG 和GG 随机设置5 个重复,不同种植年限、间距柠条灌丛地在带间、带内分别设置3 个重复,观测土壤深度为0-200 cm,每20 cm为一层,共10 层,每根TDR 管分别在4 个方向读表并记录,每月观测1~2 次;每次观测时用铝盒分别取0-5 和5-10 cm 土壤,采用烘干法对表层土壤水分进行测定。
1.3.3 土壤有机碳、全氮测定
土 壤 有 机 碳(soil organic carbon, SOC)、全 氮(total nitrogen, TN)的采样时间在荒漠草原植被生长季7 月中旬,按五点取样法用土钻取各个样点0-100 cm 土层的土壤,按5 层每层20 cm 取回土样,混匀后装入自封袋,每个样地3 次重复,以备后期室内试验测定有机碳及全氮的分析。采用重铬酸钾容量法测定土壤有机碳[18],半微量凯氏定氮法测定土壤全氮[18]。
1.3.4 土壤微生物数量及量碳的测定
土壤微生物土样采集时间在7 月中旬,微生物三大类群的测定采用稀释平板记数法[19],分别用琼脂培养基培养细菌、马丁氏培养基培养真菌、高氏一号培养基培养放线菌,每个样地做3 次重复,比较不同浓度梯度微生物数量,培养3~7 d 并准确计数[19]。土壤微生物量碳(microbial biomass carbon,MBC)的测定采用氯仿熏蒸浸提法[19]。

表 1 样地基本情况Table 1 Description of the sampling sites
1.4 数据分析
1.4.1 土壤微生物量碳及呼吸熵的计算
土壤微生物生物量碳:MBC = EC/kEC。
式中:EC 为熏蒸土壤与未熏蒸土壤有机碳的差值,kEC为MBC 的转换系数,其取值0.45[18]。
土壤呼吸熵:qCO2= SR/MBC。
式中:SR 为土壤呼吸强度,MBC 为土壤微生物生物量碳[19]。
1.4.2 其他数据分析
数据前期整理采用Excel 2010;文中的图表采用Origin 2018 绘 制;数 据 分 析 则 采 用SPSS 19.0,采用单因素方差分析来检验各样地之间及不同土层土壤因子间的差异性,采用最小显著差数法(LSD)对数据进行多重比较,α = 0.05;各因子与土壤呼吸之间的相关性用Pearson 相关分析进行。
2 结果与分析
2.1 荒漠草原灌丛引入过程土壤水分垂直特征
草原-放牧-不同年限灌丛引入过程,EG 和GG土壤水分随土层加深而增加,深层(100-200 cm)土壤水分分别为12.42%~24.85%、14.23%~16.90%,明显高于浅层(0-100 cm)土壤水分(图2A);而灌丛引入后垂直差异不明显,分别为6.14%~13.32%、4.44%~11.37%;EG 和GG 0-200 cm 平均土壤水分均高于不同年限灌丛地,分别为14.91%和13.07%。不同间距灌丛地与不同年限灌丛地相似(图2B),其0-200 cm 土壤水分均低于EG 和GG,且无明显垂直变化,100-200 cm 土壤水分高于0-100 cm,分别为7.83%~14.89%、4.44%~11.61%;不同间距灌丛地(SI40、SI6、SI2)平均土壤水分均低于EG和GG,其值分别为9.13%、8.74%和10.49%。
2.2 荒漠草原灌丛引入过程土壤水分季节变化
草原-放牧-不同年限灌丛引入过程(图3A),EG 和GG 0-200 cm 土壤水分存在明显季节动态,即3-4 月春季返潮期、5-8 月生长消耗期、9-11 月秋季雨水补充期。EG、GG 样地平均土壤水分分别为14.91%、13.14%,高于SY3 (8.51%)、SY12 (9.66%)、SY22 (8.67%),但EG 0-10 cm 土壤水分较低且无明显季节变化,其范围为0.44%~3.22%,其他样地季节波动较大。草原-放牧-不同间距灌丛引入过程土壤水分差异明显(图3B),0 -200 cm,EG、GG 土壤水分均高于灌丛地,不同间距与不同年限灌丛地土壤水分季节变化相似,3 个时期的土壤水分分别为7.80%~10.89%、7.80%~10.37%和8.83%~12.05%;而灌丛地0-10 cm 土壤水分季节变化明显,其范围在0.89%~6.53%。
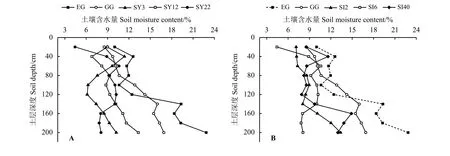
图 2 灌丛引入过程土壤含水量垂直特征Figure 2 Vertical characteristics of soil’s moisture content during shrub introduction
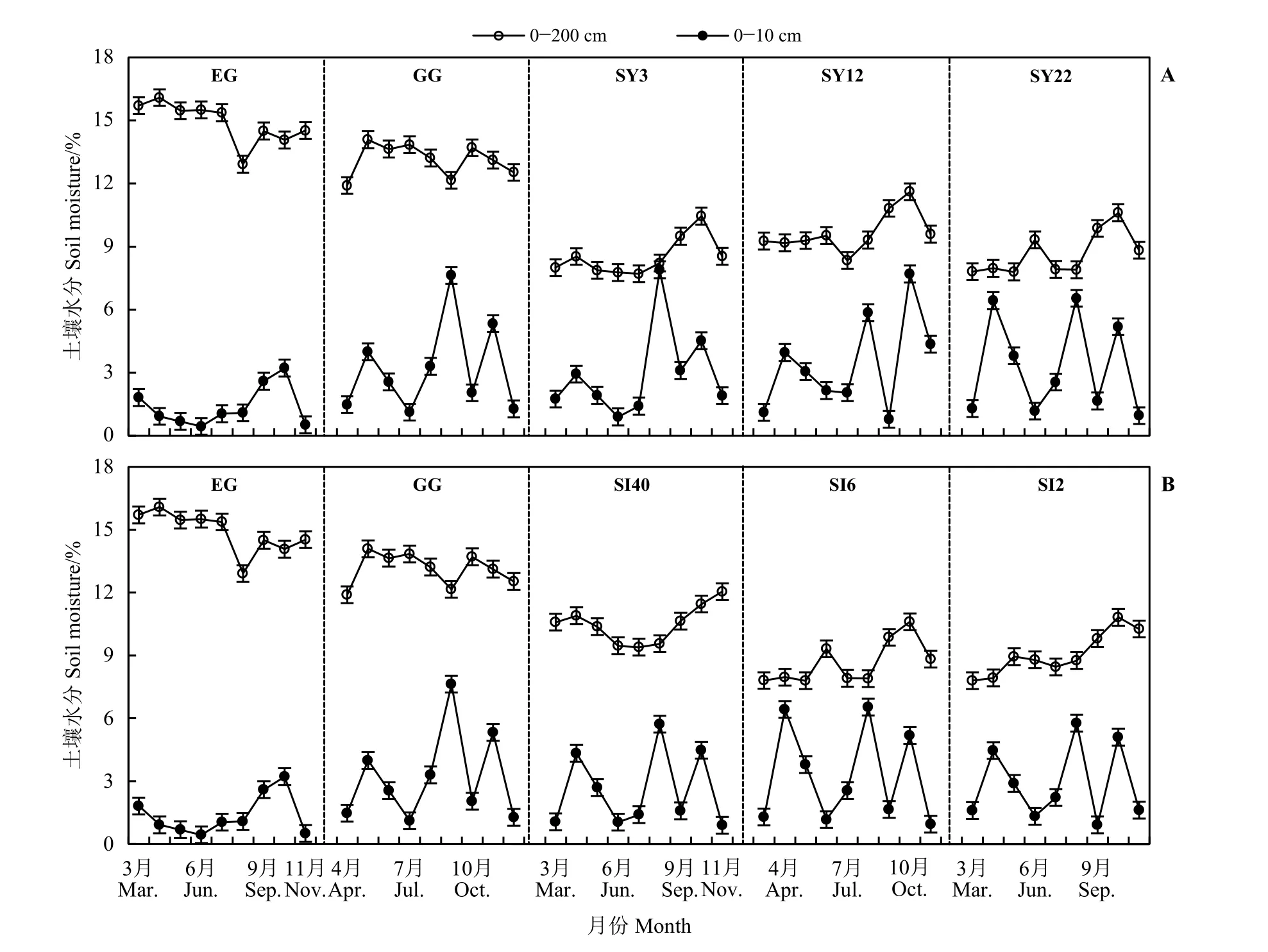
图 3 灌丛引入过程土壤水分季节动态Figure 3 Seasonal dynamics of soil moisture during shrub introduction
2.3 荒漠草原灌丛引入过程土壤呼吸特征
草原-放牧-不同年限灌丛引入过程,土壤呼吸速 率 在0.016~0.544 g·(m2·h)-1,在3 月-5 月EG 土壤呼吸速率逐渐降低,而GG 土壤呼吸略升后又逐渐降低,在5 月-11 月土壤呼吸呈现先升后降再升又降的双峰波动,EG、GG 在8 月份有最大值,分别为0.506、0.470 g·(m2·h)-1;灌丛地生长季土壤呼吸表现为SY3<SY22<SY12,其土壤呼吸速率峰值也在8 月份,分别为0.436、0.468、0.544 g·(m2·h)-1(图4A)。此外,随着种植年限的增加,年均土壤呼吸速率降低,SY22 最低,为0.126 g·(m2·h)-1,显 著 低 于 其 他 样 地(P<0.05) (图5A)。草原-放牧-不同间距灌丛引入过程,土壤呼吸速率范围在0.008~0.506 g·(m2·h)-1,土壤呼吸速率随季节波动明显且均出现两个峰值,SI40、SI6、SI2 最 大 值 分 别 为0.301、0.468、0.234 g·(m2·h)-1;随着灌丛密度的增大表现出先增后减的趋势,在11 月土壤呼吸速率最低,分别为0.008、0.016、0.020 g·(m2·h)-1(图4B)。不同间距灌丛地显著低于EG、GG (P<0.05),且随着密度的增大,年均土壤呼吸速率降低,SI2 最低,为0.103 g·(m2·h)-1,显著低于SI40、SI6 (P<0.05) (图5B)。
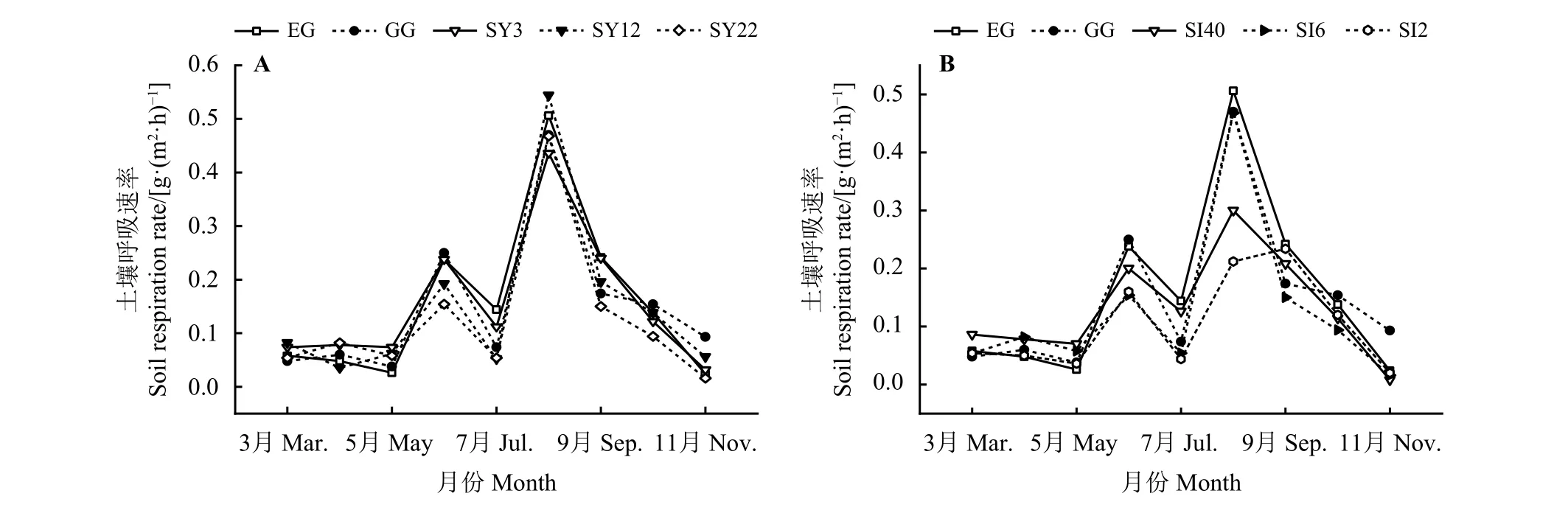
图 4 灌丛引入过程土壤呼吸季节动态变化Figure 4 Seasonal dynamics of soil respiration during shrub introduction
2.4 荒漠草原灌丛引入过程土壤有机碳、全氮变化
草原-放牧-不同年限灌丛引入过程(表2),各样地土壤有机碳在不同土层均存在不同程度的差异,EG、GG 样地SOC 随土层加深呈现先升高后降低趋势,其范围分别为1.87~4.80、1.09~3.08 g·kg-1;随着年限的增加0-60 cm,SOC 表现为先降低后升高,而60-100 cm 则表现出持续降低的趋势,SY3 在40-60 cm 土层与SY22 有显著差异(P<0.05),而与SY12 各土层之间均无显著差异(P > 0.05)。土壤全氮(TN)除EG 样地在20-40 cm与80-100 cm 间有显著差异(P<0.05)外,其余各样地不同土层间均无显著差异(P > 0.05),且放牧地土壤全氮含量较低,其范围在0.19 ~0.32 g·kg-1;而随着灌丛年限的增加各土层TN 也呈增大趋势,SY22 在40-80 cm 显著高于SY3、SY12,在其他土层之间均无显著差异(P > 0.05)。
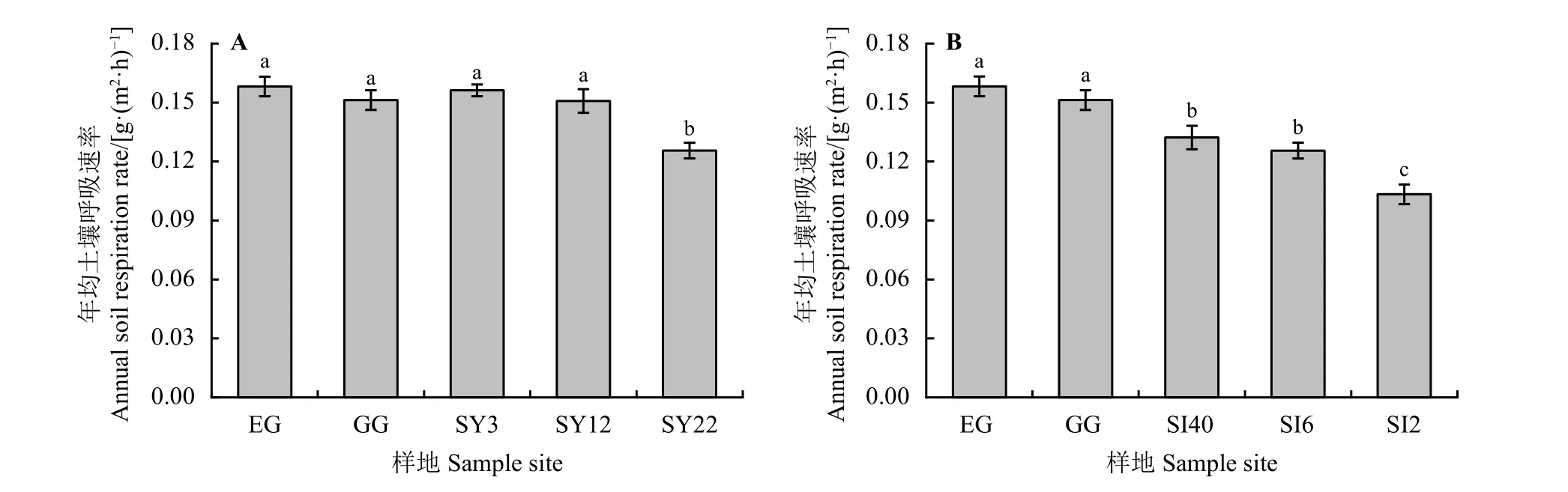
图 5 灌丛引入过程年均土壤呼吸速率变化Figure 5 Changes of annual soil respiration rate during shrub introduction
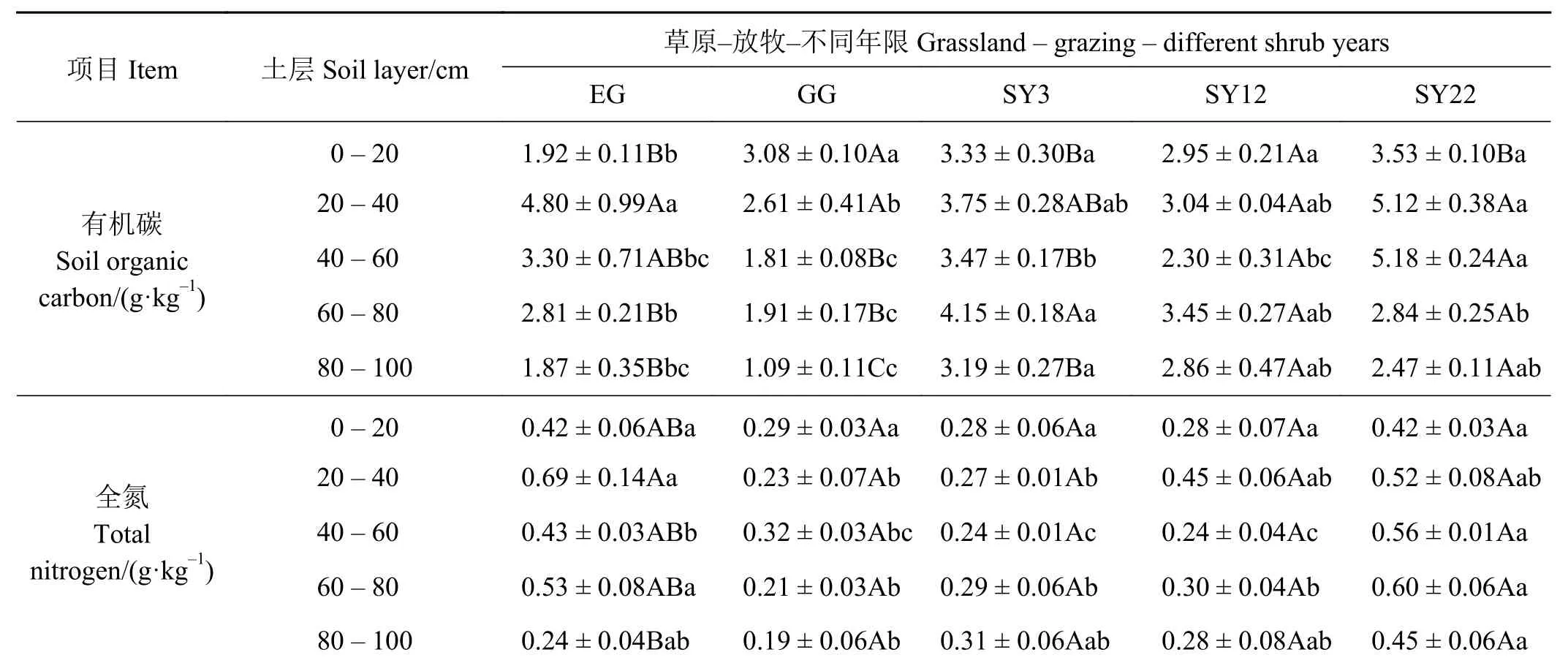
表 2 草原-放牧-不同年限灌丛引入过程土壤有机碳、全氮变化Table 2 Soil organic carbon and total nitrogen during grassland-grazing-different shrub years
草原-放牧-不同间距灌丛引入过程(表3),在0 -20 cm 土层EG 样地SOC 最小,显著低于其他样地(P<0.05),但GG 样地在0-20 cm 最大,其含量为3.08 g·kg-1,各样地SOC 最低值均在80-100 cm,且随着间距的减小,0-20 cm 土层SOC 含量逐渐降低,20-60 cm 表现为先升高后降低,60-100 cm则表现为先降低后升高的趋势。相较于其他样地,EG 样地土壤全氮含量在各层都较高,其范围在0.24 ~ 0.69 g·kg-1,且80-100 cm 显著低于其他土层(P<0.05);随着灌丛间距的减小,TN 均表现出先升高后降低的趋势,且同一样地不同土层之间TN 差异均不显著(P > 0.05)。
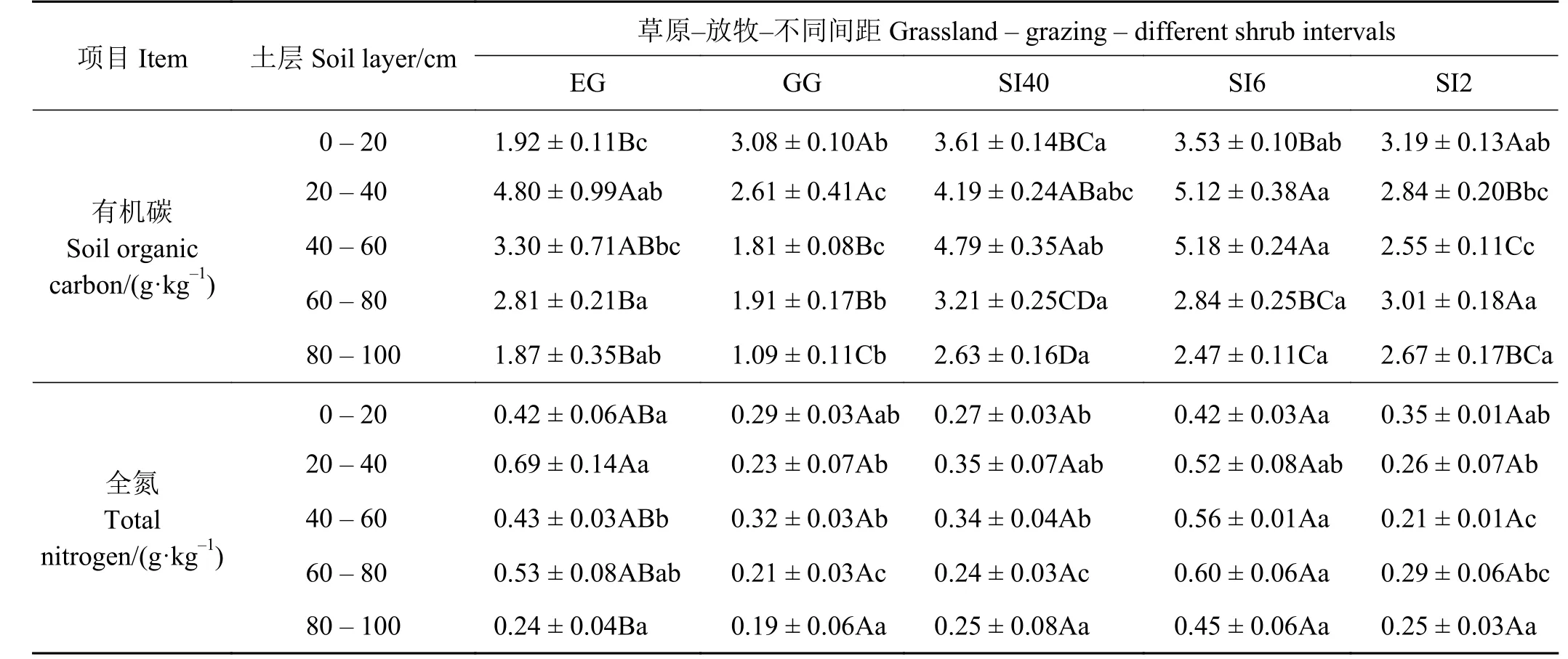
表 3 草原-放牧-不同间距灌丛引入过程土壤有机碳和全氮变化Table 3 Soil organic carbon and total nitrogen during grassland-grazing-different shrub intervals
2.5 灌丛引入过程中土壤微生物特性变化
草原-放牧-不同年限灌丛引入过程(图6A),微生物总量随年限增加而增加,SY22 最多,为3.63 ×106cfu·g-1,显著高于其他样地(P<0.05);其中真菌数量整体较低,灌丛地高于EG、GG,其中SY3最多,为1.13 × 105cfu·g-1,EG 最少,为1.40 × 104cfu·g-1;放线菌、细菌数量整体呈增加趋势,SY22 显著高于 其 他 样 地(P<0.05),分 别 为2.59 × 106、9.53 ×105cfu·g-1,而其他样地之间差异不显著(P > 0.05),分 别 为1.28× 106~1.49 × 106、3.38× 105~5.97 ×105cfu·g-1。封育-放牧-不同间距灌丛引入过程(图6B),微生物总量随种植间距的减小呈先增加后减 少的趋势,SI6 最多,为3.63 × 106cfu·g-1,SI2最少,为1.78 × 106cfu·g-1;细菌、真菌、放线菌数 量,SI6 最 多,分 别 为9.53 × 105、8.80 × 104、2.59 × 106cfu·g-1,均显著高于其他样地(P<0.05),SI2 各菌群数量明显减少。
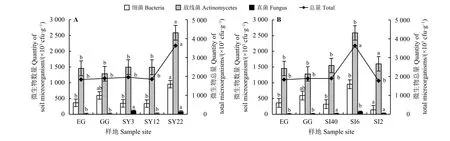
图 6 灌丛引入过程土壤微生物数量特征Figure 6 Quantitative characteristics of soil microorganisms during shrub introduction
草原-放牧-不同年限灌丛引入过程(图7A),EG、GG 样地的MBC 分别为548.80、486.40 mg·kg-1,显著高于SY3、SY12 (P<0.05),且随年限的增加MBC 整体呈增加趋势,SY22 最高为403.20 mg·kg-1;而qCO2则有降低趋势,SY22 最低为0.38。草原-放牧-不同间距灌丛引入过程(图7B),SI40 样地MBC 最高,为637.93 mg·kg-1,显著高于SI6、SI2(P<0.05),而与EG、GG 样地差异不显著(P > 0.05);且MBC 随着种植间距的缩小而降低,SI2 最低,为371.20 mg·kg-1,但qCO2则有增加趋势,SI2 最高,为0.43。
2.6 灌丛引入过程土壤呼吸与土壤水分、有机碳、全氮和土壤微生物的相关性
相关性分析得出(表4),草原-放牧-灌丛引入过程中,土壤呼吸与土壤水分、土壤微生物生物量碳正相关,且与土壤水分极显著正相关(P <0.01),而与土壤有机碳、全氮、微生物数量呈负相关,其中与放线菌数量呈显著负相关(P<0.05)。
3 讨论
3.1 荒漠草原灌丛引入对土壤水分及土壤呼吸的影响
在荒漠草原区域,土壤水分与植被生长密切相关,抗旱灌木的引入是生态修复的主要措施之一,灌木的引入可以改善土壤物理结构、增加土壤养分含量及植被多样性[20]。然而,本研究发现,封育草地0-10 cm 土壤水分含量很低,这是因为封育草地以草本为主,植被根系较浅且破坏程度轻,表层水分循环利用率高,加之研究区降水有限,无法补给深层水分,而使土壤水分较低。灌丛引入后浅层土壤水分波动较大,深层土壤水分明显降低,这是因为浅层土壤更易受降水、光 照、风力等气候因子的影响,而随着灌丛年限和密度的增加,木本植物在草地群落中的优势越来越大,其对深层土壤水分的需求也增大,因此会加快荒漠草原深层水分的消耗[17]。且本研究中,9 月-11 月随温度降低,土壤水分又有升高且波动较小,此时植物生长速率减慢导致根系对水分的利用减少[14]。潘占兵等[21]发现,灌丛密度增大的同时土壤水分亏缺越大,土壤碳损失也越大,这与本研究的结果相似。因此,宁夏东部荒漠草原合理的灌丛种植间距还需进一步探究。
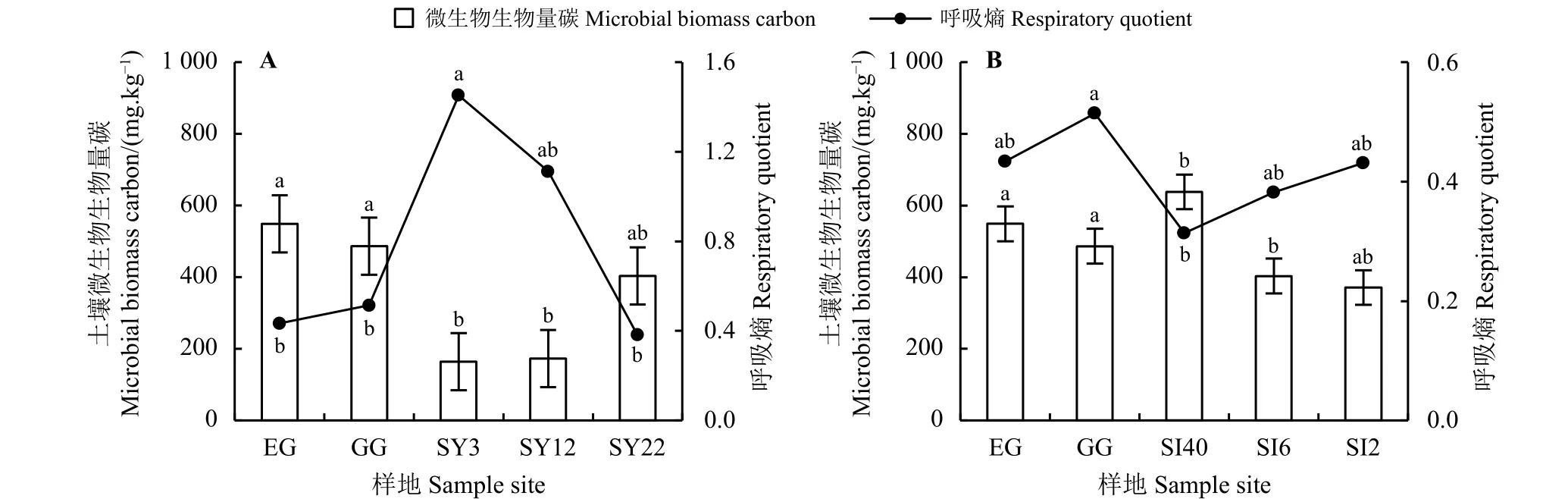
图 7 灌丛引入过程土壤微生物量碳及呼吸熵变化Figure 7 Changes of soil microbial biomass carbon and respiratory quotients during shrub introduction
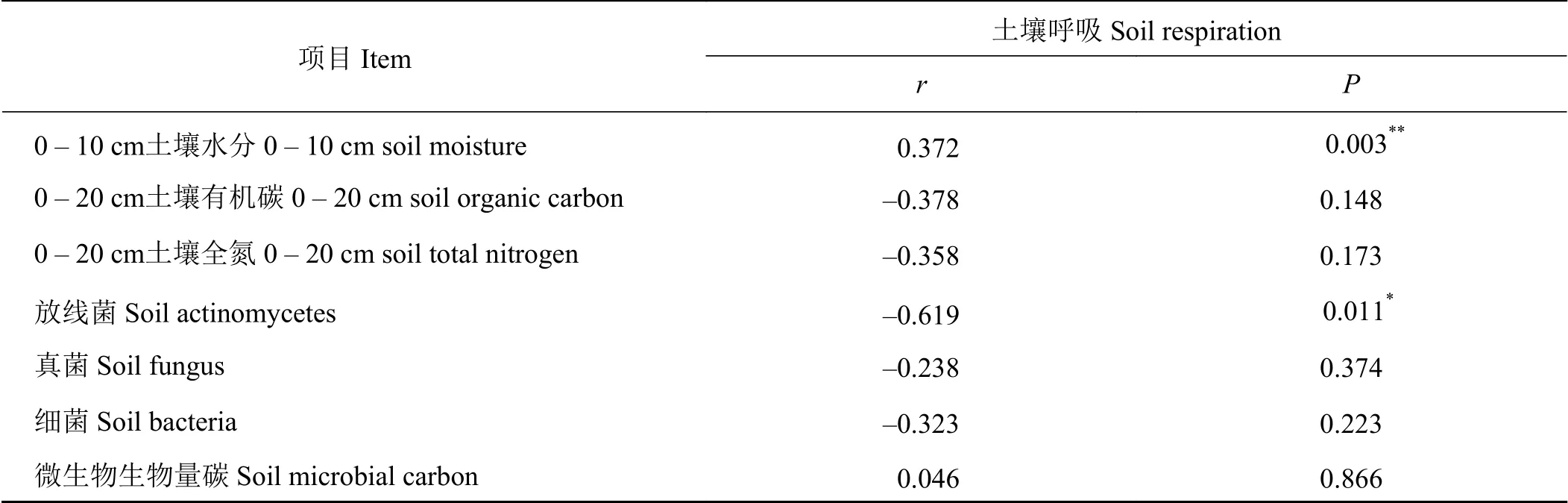
表 4 灌丛引入土壤呼吸与土壤水分、有机碳、全氮及土壤微生物的相关分析Table 4 Correlation analysis of soil respiration, soil moisture, organic carbon, total nitrogen, and soil microorganism under different shrub introduction conditions
荒漠草原土壤呼吸在草地碳循环机制中非常关键,该地区土壤-植物-大气之间的碳循环研究对生态系统稳定具有重要的科学指导意义[16],降水影响植物根系生长及微生物活动,进而对根际土壤呼吸的激发作用十分强烈[22]。已有研究指出,气候和生物因子共同控制着土壤呼吸在小尺度上的空间变异性[23]。Ying 等[24]研究得出灌木会与草本植物竞争浅层土壤水分,同时还会利用深层土壤水分,而也有研究表明以木本植物为主的林地、灌丛地的土壤呼吸强度明显高于草地区[25]。荒漠草原灌丛引入后土地斑块化和破碎化加剧[17],土壤呼吸异质性增大,灌木根系在生长过程中形成的大孔隙会加快降雨入渗至深层土壤,有助于灌木利用更多的水分[26],加速了灌丛化进程。在植被生长季内土壤呼吸呈现出明显的季节变化,且受降水影响较大[27],而本研究发现,不同年限、间距灌丛地8 月土壤呼吸强度都强于其他月份,这与此阶段该地区温度适宜且降水集中有关,但灌丛地土壤呼吸随着年限的增加而降低,虽然灌丛地凋落物多,但分解产物无法及时被利用就会产生土壤碳流失[28],而封育、放牧草地对水分变化更加敏感,在干旱时土壤呼吸较低,碳损失也低。
3.2 荒漠草原灌丛引入对土壤碳氮及微生物的影响
灌丛引入会改变荒漠草原原有的物种组成、地上植被及土壤资源空间分布,引起植被多样性和群落结构的变化,从而产生生境异质性[29]。灌丛附近的有机碳和全氮含量比邻近的草地空斑高,灌木层下形成“沃岛效应”导致水分和养分的富集,加剧资源的空间异质性和土地斑块化[16,30-31]。Cable等[10]在美国灌丛化草地的土壤呼吸研究中得出,灌木的扩张会引起呼吸的异质性,大灌丛附近土壤呼吸速率和温度敏感性更高。本研究中封育草地表层有机碳、全氮含量整体较低,而放牧地则有表层聚集的现象。随着灌丛的引入,豆科植物与根瘤菌固定了更多空气中的游离态氮,如果氮沉降持续增加,就会严重威胁生态系统健康[29],使土壤微生物生物量下降活性降低,进而引起土壤微生物与植物养分分配不均,对土壤微生物呼吸和根系呼吸产生影响[32]。盐池荒漠草原夏季光照充足,植物生长快,这使得植被覆盖和生物多样性增大,土壤呼吸和微生物活动增强,从而加快光合产物与土壤之间的转化,因此夏秋季土壤呼吸速率要高于冬春季,这与熊莉等[33]的研究结果相似。此外,与本研究的结果相似,灌丛斑块土壤表层有机碳和全氮都高于草地斑块且随灌丛由近到远其含量逐渐降低[34],根际土壤中全氮、有机质的含量均高于非根际土壤[35],而灌丛下凋落物分解更快从而增加了灌丛周围的土壤有机质含量,这可能是因为灌木叶片中木质素和纤维素含量较低[36]。灌丛引入有利于有机质的转化和土壤微生物活性的增强,从而改变土壤肥力[37]。
草地生态系统中养分循环利用存在复杂性和不确定性,灌丛地土壤变化对土壤细菌、真菌影响显著,而对放线菌影响较小,草本植物对细菌活动影响更加密切[38]。而本研究中,封育草地和放牧地细菌数量较低,这可能与采样期气候干旱、降雨少、放牧等因素有关。Ochoa-Hueso 等[39]在对北美和澳大利亚两个区域的7 个草地进行模拟降水试验得出,干旱条件对微生物群落组成影响显著,细菌群落比真菌对水分的限制更加敏感,而放线菌适应干旱条件的能力强,菌落的相对丰度会因干旱而增加,这与本研究结果相似。此外,本研究发现封育、放牧草地微生物生物量碳高于灌丛地,且随年限的增加和间距的缩小也呈整体增加趋势,但到一定年限后其随种植间距的缩小而降低,这可能与土地利用方式、土壤有机碳含量、土壤温湿度、季节波动等有关[40]。草地有助于调节全球土壤碳库的年际动态变化,微生物群落结构和功能对干旱、气候变化和碳循环的响应至关重要[16]。因此,为应对全球气候变化,未来对荒漠草原土壤呼吸的研究应该重点关注不同环境因子作用下不同物种根系呼吸和根际微生物活动,从而进一步探究荒漠草原土壤微生物对土壤呼吸的影响。
4 结论
在草原-放牧-灌丛引入过程中,灌丛地深层土壤水分低于封育、放牧草地,无明显垂直动态。各样地中土壤呼吸均在8 月有最高值,封育、放牧草地高于显著高于不同间距灌丛地(P<0.05),但与不同年限灌丛地差异不显著(P > 0.05)。封育、放牧草地土壤有机碳随土层的加深而降低,而灌丛地深层有机碳含量高于放牧地(P<0.05),与封育草地无显著差异(P > 0.05);土壤全氮在封育草地各土层含量均较高,放牧地较低,且全氮随着年限的增加和间距的缩小而增大,而当种植间距最小时又有降低趋势。各菌群数量随灌丛的引入而显著增加,土壤微生物量碳随种植年限和间距的增加而增大,但呼吸熵则降低。土壤呼吸与土壤水分、土壤微生物量碳正相关,且与土壤水分极显著正相关(P<0.01),而与土壤有机碳、全氮、微生物数量负相关,其中与放线菌数量显著负相关(P<0.05)。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
小哥白尼(趣味科学)(2022年5期)2022-08-15
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
绿色中国(2019年14期)2019-11-26
现代园艺(2017年13期)2018-01-19
植物资源与环境学报(2017年3期)2017-10-12
儿童故事画报·智力大王(2016年7期)2017-02-08
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
文史春秋(2016年3期)2016-12-01
土壤与作物(2015年3期)2015-12-08
