
miR-186介导的PTTG1下调对膀胱癌细胞增殖、凋亡、侵袭和迁移的影响研究
2019-12-10 06:10余洋刘斌波熊飞
现代泌尿生殖肿瘤杂志 2019年5期
余洋 刘斌波 熊飞
膀胱癌是临床上常见的恶性肿瘤,其发病率居泌尿系统首位[1]。同其他恶性肿瘤一样,其发生、发展涉及癌基因激活、抑癌基因失活等系列复杂过程[2-4]。而微小核糖核酸(microRNA, miRNA)通过转录后水平降解mRNA分子或抑制其表达参与肿瘤生成、细胞凋亡、发育调控、器官形成、新陈代谢等[5-7]。它在进化上高度保守,是一种长度约19~24 nt的短序RNA,本身不具有蛋白编码功能。作为其重要成员的miR-186定位于人类1号染色体,前体序列长约70 nt,其与多种肿瘤的发生、发展密切相关。Yao等[8]的研究显示,在膀胱癌组织和细胞系中,miR-186表达下调,可靶向抑制核小体结合蛋白1的表达进而抑制肿瘤细胞增殖和侵袭能力。Yang等[9]的研究显示,miR-186可通过下调磷酸化的蛋白质磷酸酶1B而影响膀胱癌细胞周期进展。另有He等[10]的研究显示,miR-186可通过血管内皮生长因子C调控膀胱癌细胞侵袭和转移能力。可见,其参与膀胱癌的发生、进展和结局调控,但其具体信号通路或机制尚待进一步研究。
垂体肿瘤转化基因1(pituitary tumor transforming gene 1, PTTG1)由Pei等[11]于1997年在大鼠垂体肿瘤组织中首次发现,1999年Zhang等[12]首次将其从人胚胎干细胞中分离。后续研究证实PTTG家族至少包括PTTG1、PTTG2和PTTG3 3个成员。而人体肿瘤中主要表达PTTG1,其具有促进肿瘤细胞转化、诱导肿瘤新生血管形成、导致肿瘤复发和转移等生物学功能[13-16]。多项研究表明,在前列腺癌、乳腺癌、卵巢癌、宫颈癌等多种肿瘤组织和细胞系中PTTG1表达升高,且与这些恶性肿瘤的发生、进展、预后等密切相关,可能成为重要的肿瘤标志物和治疗靶点[17-20]。但目前PTTG1在膀胱癌中的研究相对较少。Xiang等[21]的研究提示,miR-146a-3p可调控PTTG1的表达,促进膀胱癌的迁移、侵袭、转移和生长。关于miR-186和PTTG1在膀胱肿瘤细胞中的表达情况及其靶向关系,以及两者对膀胱肿瘤细胞增殖、凋亡、侵袭和迁移影响的研究也甚少。
本研究采用qRT-PCR检测50例膀胱癌及相应的癌旁组织中miR-186和PTTG1核酸含量,Western blotting检测PTTG1蛋白含量,通过生物信息学预测miR-186和PTTG1结合位点,并利用双荧光素酶报告实验验证两者的靶向关系。为了证实miR-186是否通过靶向调控PTTG1而影响膀胱癌细胞的生物学行为,我们将miR-186抑制物或miR-186抑制物+设计合成的PTTG1特异性siRNA转染膀胱癌细胞J82,观察抑制miR-186或同时抑制miR-186和PTTG1后细胞增殖、侵袭、迁移和凋亡的变化,同时监测PCNA、Bcl-2、Bax、MMP-2和E-cadherin以探讨其对上述各信号分子的作用,以期加深对膀胱癌发生、发展及结局机制的了解,为膀胱癌早期诊疗、基因治疗及预后判断提供更多理论依据。
材料与方法
一、材料
1.膀胱癌组织来源:收集2017年7月至2018年6月期间在宜昌市中心人民医院泌尿外科行膀胱全切术的膀胱癌患者癌组织标本共计50例,详细记录手术时间、年龄、性别、病理资料等。所有患者术前均未行放、化疗和免疫治疗。其中男32例,女18例;年龄39~80岁,平均年龄(60.02±7.51)岁。行膀胱全切术时,同时取距离肿瘤组织2 cm以上区域的正常膀胱组织(癌旁组织)。所有标本的采集经过医院允许,术前所有患者签订《标本处理知情同意书》,并自愿提供膀胱组织标本。
2.膀胱癌细胞来源:膀胱癌J82细胞购自中国科学院上海细胞库,按照细胞的培养条件进行培养。
3.主要试剂和仪器:总RNA提取试剂盒、逆转录试剂盒、PCR扩增试剂盒,购自大连TAKARA公司;BCA试剂盒、HRP标记的羊抗鼠IgG、ECL试剂盒,购自美国Santa Cruz公司;小鼠抗人PTTG1抗体,购自美国Abcam公司;PVDF膜、ECL增强发光液,购自美国Millipore 公司;双荧光素酶报告基因检测试剂盒,购自美国BioAssay Systems公司;Lipofectamine 2000试剂盒,购自美国Invitrogen公司;MTT细胞活性检测试剂,购自中国碧云天生物科技有限公司;Annexin Ⅴ-FITC细胞凋亡试剂盒、基质胶,购自美国BD公司;Transwell小室,购自美国Coring公司;鼠抗人PCNA、Bcl-2、Bax、MMP-2和E-cadherin抗体,购自美国Abcam公司;电泳仪、紫外分光光度计,购自北京六一生物科技有限公司;凝胶成像系统、垂直型蛋白电泳装置,购自美国Bio-Rad公司;PCR仪、CO2细胞培养箱,购自美国Thermo公司;大型台式恒温摇床,购自美国NBS公司;酶标仪,购自美国Bio-Rad公司;流式细胞仪,购自美国BD公司;细胞计数板,购自上海生化试剂仪器公司;倒置荧光显微镜,购自日本Olympus公司。
二、实验方法
1.膀胱癌组织和细胞系中miR-186和PTTG1核酸含量及蛋白水平测定:选取生长状态良好的膀胱癌细胞,依据总RNA提取试剂盒的说明提取细胞总RNA,紫外分光光度计检测提取的RNA浓度,测定260 nm和280 nm处的光密度(optical density,OD)值的比值(OD260/OD280),估计核酸的纯度,OD260/OD280为1.8~2.0的RNA用于后续实验。检测合格的RNA放置在-80 ℃冰箱中保存。依据反转录试剂盒的说明将总RNA逆转录cDNA,反转录程序:37 ℃,60 min;85 ℃,5 min;4 ℃,终止反应。将cDNA模板放于-20 ℃冰箱中保存。依据引物设计原则进行引物的设计,引物序列如下:miR-186引物(上游-5′-CCCGATAAAG-CTAGATAACC-3′,下游-5′-CAGTGCGTGTCGT-GGAGT-3′);内参U6引物(上游-5′-GCTTCGGCAGCACATATACTAAAAT-3′,下游-5′-CAG-TGCGTGTCGTGGAGT-3′);PTTG1引物(上游-5′-TGATCCTTGACGAGGGAG-3′,下游-5′-GGT-GGCAATTCAACATCCAGG-3′);内参GAPDH引物(上游-5′-AGAAGGCTGGGGCTCATTT-3′,下游-5′-AGGGGCCATCCACAGTCTTC-3′)。反应体系:总RNA 1 μg,5×缓冲液 4 μl, 反转录酶混合物 1 μl,引物混合物 1 μl,去离子水补足至25 μl。反应条件:37 ℃,60 min;85 ℃,5 min,反应终止后,将cDNA模板放置于-20 ℃冰箱中保存。然后根据不同反应条件进行定量扩增,最后针对产物进行荧光信号定量分析,以所得溶解曲线及Ct值,按照公式2-△△Ct计算miR-186(以U6为内参)和PTTG1(以GAPDH为内参)的表达量,其中△Ct=Ct目的基因-Ct内参基因。选取备用组织和细胞,加入RIPA裂解液,冰盒中裂解反应30 min,12 000 rpm,4 ℃离心10 min,取上清至离心管中,部分用于后续实验,剩余放置于-20 ℃冰箱保存。BCA法定量蛋白。经SDS-PAGE电泳,转膜,膜封闭和一、二抗体孵育过程。再应用 ECL显色液显色,Image J软件分析图像。蛋白相对表达量为目的蛋白灰度值与内参GAPDH灰度值。
2.双荧光素酶报告实验:通过TargetScan、miRBase和miRanda生物信息学分析软件发现miR-186和PTTG1存在可结合的位点。24孔板中培养膀胱癌J82细胞,将构建的miR-186 mimics与PmiR-GLO-PTTG1-3′UTR共转染至J82细胞中,PTTG1基因的3′UTR 区克隆至载体海肾荧光素酶基因下游位点。构建结合miR-186 mimics与对照(miR-scramble)的野生型(WT)报告质粒及突变型(MUT)报告质粒。依照Lipofectamine 2000试剂盒说明,转染构建好的质粒至J82细胞。转染48 h后,分别以海肾荧光素酶基因及萤火虫荧光素酶作为内参基因和报告基因,通过双荧光素酶报告检测系统进行检测。
3.细胞增殖实验:使用MTT试剂检测J82细胞活力。收集转染后的细胞,制备成单细胞悬液,并将细胞浓度调整为1×104个/ml。在96孔板中加入细胞悬液,每孔100 μl,每组设置5个复孔,并设置加入培养液的孔为零孔。分别收集培养24、48、72和96 h的细胞,避光条件下在每孔中加入5 mg/ml的MTT溶液20 μl,培养箱中放置4 h,将孔内液体吸出,再在每孔中加入DMSO溶液150 μl,摇床低速震荡10 min,结晶完全溶解后,使用酶标仪测定490 nm波长处各组各个孔的OD值,记录结果,取均值。以所测的时间为横坐标,检测的OD值为纵坐标,绘制生长曲线。实验重复3次。
4.细胞迁移实验:将J82细胞分为anti-NC组、anti-miR-186组和anti-miR-186+si-PTTG1组。胰蛋白酶消化转染48 h的细胞,无血清培养液重悬细胞,制备成单细胞悬液,并调整细胞浓度为5×105个/ml。取200 μl细胞悬液,接种于Transwell小室的上室,然后沿小室外侧孔缓慢的加入600 μl含10% FBS的DMEM培养液。依据细胞迁移能力不同,调整细胞迁移时间,到达时间后,从培养箱中将小室取出,棉签轻轻拭去培养室内培养液,小室放于多聚甲醛(4%)中固定,20 min后,使用棉签将多聚甲醛拭去。转移小室至结晶紫液中,避光染色25 min。冲洗掉结晶紫液,棉签擦干小室。显微镜下随机选择5个视野(上、中、下、左、右),计数,记录数据。取均值,对各组数据进行统计学分析。
5.细胞侵袭实验:铺板前1 h,将基质胶于4 ℃环境解冻,24孔板4 ℃提前预冷。取100 μl基质胶平铺在Transwell小室的上室,37 ℃条件下放置过夜,使基质胶能聚合成凝胶。按照3∶1比例将无血清培养液与基质胶混匀,取稀释后的基质胶40 μl,铺在Transwell小室上室,4 ℃环境放置30 min,然后转移至37 ℃放置1~2 h。后续实验步骤同细胞迁移实验。
6.细胞凋亡实验:采用Annexin Ⅴ-FITC双染法,通过流式细胞术检测各组细胞的凋亡率。
三、统计学方法
采用SPSS 21.0进行数据的处理与分析,结果采用均数±标准差表示。所有实验均重复3次,使用单因素方差分析及t检验进行组间比较,以P<0.05为差异有统计学意义。
结 果
一、miR-186和PTTG1在膀胱癌细胞中的表达
为选择最适宜的用于后续实验研究的膀胱癌细胞系,本研究以人膀胱上皮永生化细胞SV-HUC-1为对照,检测了膀胱癌J82、T24、HT1376和RT4细胞miR-186和PTTG1的表达情况,结果显示,SV-HUC-1、J82、T24、HT1376和RT4细胞miR-186 mRNA表达水平分别为1.000±0.019、0.189±0.022、0.255±0.031、0.416±0.042、0.527±0.056,PTTG1 mRNA表达水平分别为1.000±0.057、7.112±0.531、5.987±0.501、5.112±0.311、4.121±0.325,PTTG1相对蛋白表达水平分别为0.081±0.013、0.652±0.059、0.448±0.052、0.372±0.043、0.187±0.027,与SV-HUC-1细胞比较,miR-186 mRNA表达降低,PTTG1 mRNA表达及蛋白表达升高,差异有统计学意义(P<0.05)。见图1。
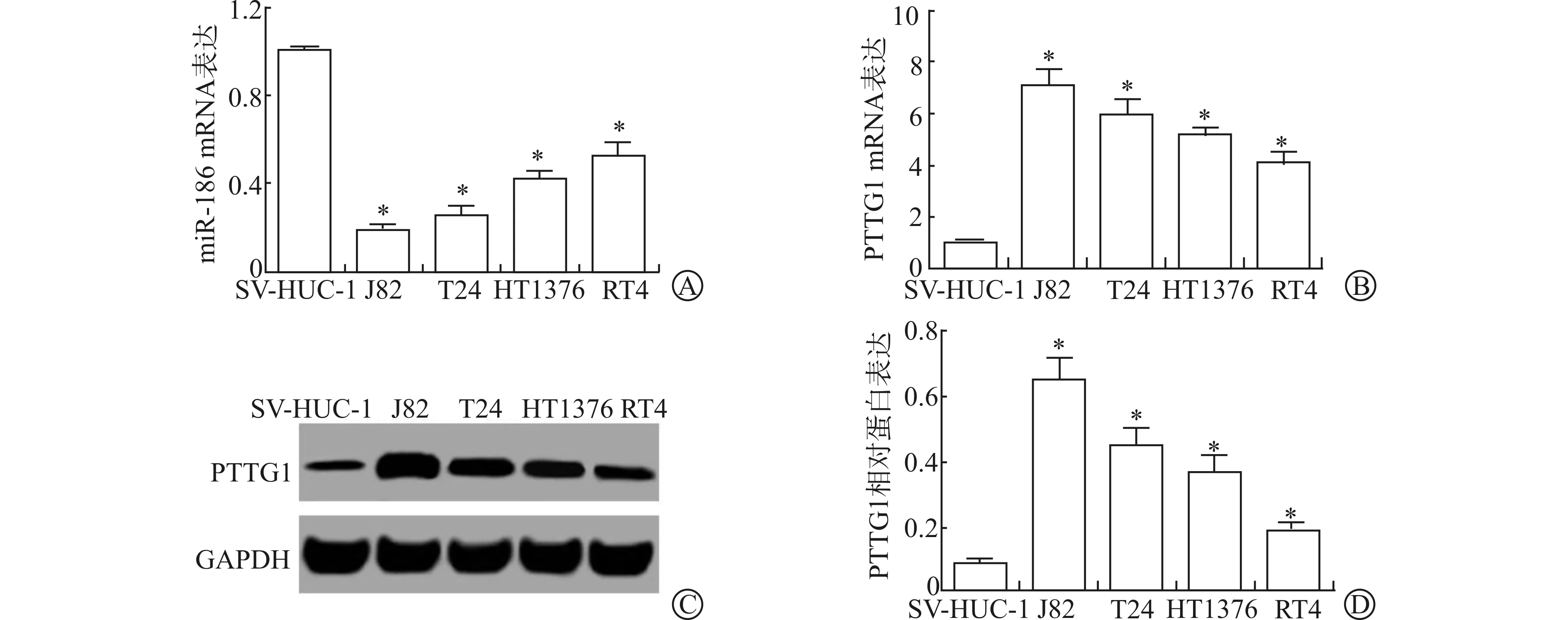
A、B:qRT-PCR检测不同膀胱癌细胞miR-186和PTTG1表达水平;C:Western blotting检测不同膀胱癌细胞PTTG1表达;D:不同膀胱癌细胞PTTG1相对蛋白表达量(与SV-HUC-1细胞比较 * P<0.05)图1 不同膀胱癌细胞中miR-186和PTTG1表达水平
二、双荧光素酶活性检测结果
生物信息学软件预测显示miR-186和PTTG1存在可结合位点(图2A)。为了证实miR-186和PTTG1之间是否存在靶向关系,构建含结合位点的PTTG1-WT和PTTG1-MUT质粒,并与miR-186共转染J82细胞,通过双荧光素酶活性检测转染后的荧光素酶活性。结果显示,miR-186 mimics与PTTG1-WT质粒共转染后相对荧光素酶活性为0.302±0.046,miR-scramble与PTTG1-WT质粒共转染后相对荧光素酶活性为1.000±0.034,两者比较差异有统计学意义(P<0.05)。miR-186 mimics与PTTG1-MUT质粒共转染后相对荧光素酶活性与miR-scramble+PTTG1-MUT组比较差异无统计学意义(P>0.05)。见图2B。
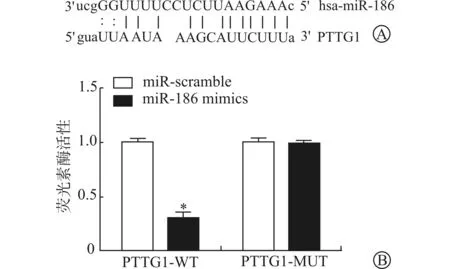
A:miR-186和PTTG1的结合位点;B:双荧光素酶报告实验检测PTTG1-WT/MUT与miR-186 mimics共转染后的荧光素酶活性(与miR-scramble组比较 * P<0.05)图2 miR-186和PTTG1的结合位点及双荧光素酶报告实验检测荧光素酶活性
三、抑制miR-186表达减弱PTTG1下调对J82细胞增殖的影响
将miR-186抑制物转染J82细胞,qRT-PCR检测转染后细胞中miR-186的表达,结果显示,anti-miR-186转染J82细胞后,miR-186表达水平为0.351±0.045,其表达水平明显低于anti-NC组(1.111±0.026)(P<0.05)。将J82细胞分为anti-NC组、anti-miR-186组和anti-miR-186+si-PTTG1组,通过MTT法检测抑制miR-186和同时抑制miR-186、PTTG1表达后J82细胞增殖能力的变化。结果显示,与anti-NC组比较,anti-miR-186组在转染后24~96 h的增殖能力均明显升高(P<0.05);而与anti-miR-186组比较,anti-miR-186+si-PTTG1组在转染后24~96 h的增殖能力均明显降低(P<0.05)。见图3、表1。
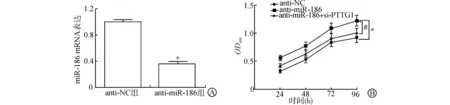
A:anti-miR-186转染后的J82细胞中miR-186表达;B:抑制miR-186和PTTG1表达后J82细胞OD值变化(与anti-NC组比较 * P<0.05;与anti-miR-186组比较 # P<0.05)图3 抑制miR-186和PTTG1表达后J82细胞增殖曲线
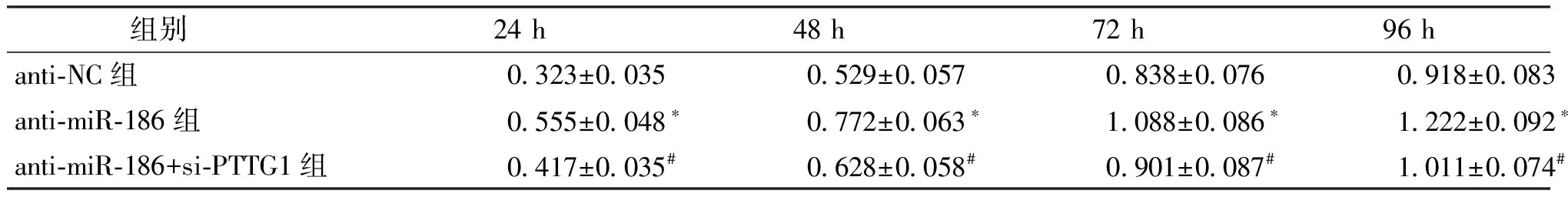
组别24 h48 h72 h96 hanti-NC组0.323±0.0350.529±0.0570.838±0.0760.918±0.083anti-miR-186组0.555±0.048∗0.772±0.063∗1.088±0.086∗1.222±0.092∗anti-miR-186+si-PTTG1组0.417±0.035#0.628±0.058#0.901±0.087#1.011±0.074#
与anti-NC组比较*P<0.05;与anti-miR-186组比较#P<0.05
四、抑制miR-186表达减弱PTTG1下调对J82细胞侵袭和迁移的影响
Transwell小室检测抑制miR-186和同时抑制miR-186、PTTG1表达后J82细胞侵袭和迁移数目的变化。结果显示,anti-NC组、anti-miR-186组和anti-miR-186+si-PTTG1组J82细胞侵袭数分别为(122.5±4.9)、(172.9±6.2)和(144.4±4.2)个,迁移细胞数分别为(152.8±6.1)、(214.3±7.9)和(182.4±6.8)个。与anti-NC组比较,anti-miR-186组细胞侵袭及迁移数明显升高(P<0.05);与anti-miR-186组比较,anti-miR-186+si-PTTG1组细胞侵袭及迁移数明显降低(P<0.05)。见图4。
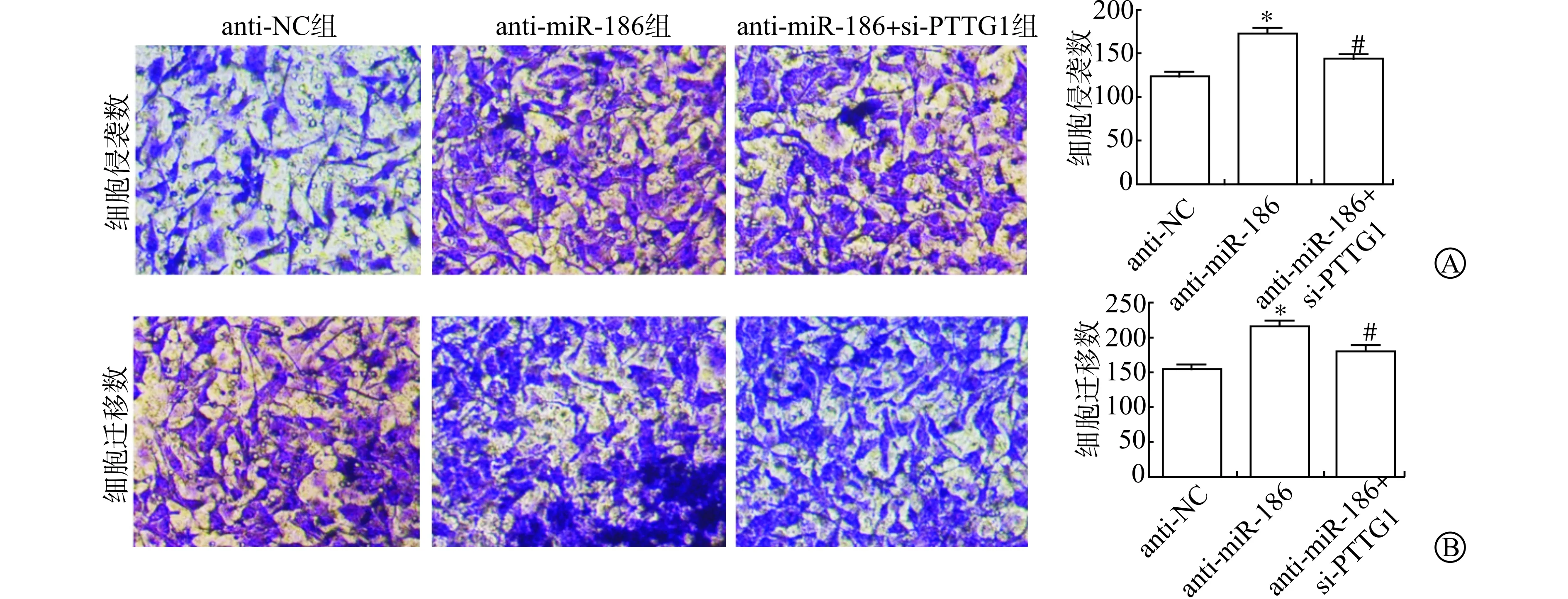
A:Transwell小室检测抑制miR-186和PTTG1表达后J82细胞侵袭变化;B:Transwell小室检测抑制miR-186和PTTG1表达后J82细胞迁移变化(与anti-NC组比较 * P<0.05;与anti-miR-186组比较 # P<0.05)图4 抑制miR-186和PTTG1表达后J82细胞侵袭和迁移变化
五、抑制miR-186表达减弱PTTG1下调对J82细胞凋亡的影响
流式细胞术检测抑制miR-186和同时抑制miR-186、PTTG1表达后J82细胞凋亡率的变化,结果显示,anti-NC组、anti-miR-186组和anti-miR-186+si-PTTG1组细胞凋亡率分别为(26.41±1.52)%、(11.13±0.86)%和(20.20±1.45)%。与anti-NC组比较,anti-miR-186组细胞凋亡率明显降低(P<0.05);与anti-miR-186组比较,anti-miR-186+si-PTTG1组细胞凋亡率明显升高(P<0.05)。见图5。
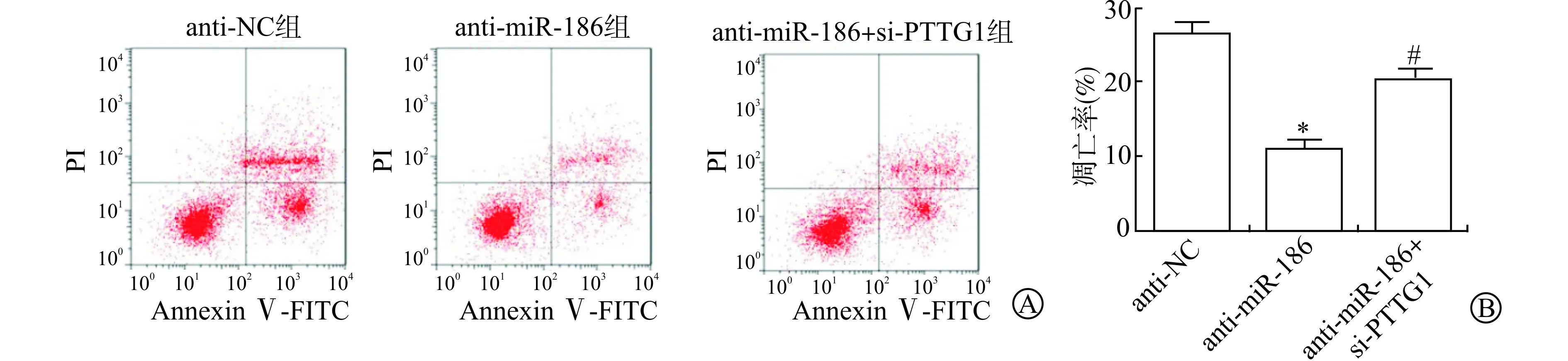
A:Annexin Ⅴ-FITC/PI双染法检测抑制miR-186和PTTG1表达后J82细胞凋亡情况;B:各组细胞的凋亡率(与anti-NC组比较 * P<0.05;与anti-miR-186组比较 # P<0.05)图5 抑制miR-186和PTTG1表达后J82细胞凋亡率的变化
六、抑制miR-186表达减弱PTTG1下调对J82细胞PCNA、Bcl-2、Bax、MMP-2和E-cadherin表达的影响
抑制miR-186和同时抑制miR-186、PTTG1表达后,Western blotting检测J82细胞中PCNA、Bcl-2、Bax、MMP-2和E-cadherin的表达,结果显示,anti-NC组、anti-miR-186组和anti-miR-186+si-PTTG1组PCNA蛋白表达分别为 0.104±0.013、0.655±0.071、0.312±0.034,Bax/Bcl-2比例分别为3.556±0.123、0.216±0.023、0.678±0.051,MMP-2蛋白表达分别为 0.078±0.010、0.289±0.032、0.163±0.015,E-cadherin蛋白表达分别为 0.499±0.051、0.124±0.018、0.266±0.030。与anti-NC组比较,anti-miR-186组细胞PCNA和MMP-2表达明显升高,Bax/Bcl-2比例和E-cadherin表达明显降低(P<0.05);与anti-miR-186组比较,anti-miR-186+si-PTTG1组细胞PCNA和MMP-2表达明显降低,Bax/Bcl-2比例和E-cadherin表达明显升高(P<0.05)。见图6。
讨 论
近年来,膀胱癌的发病率呈现出逐年升高的趋势,严重威胁着膀胱癌患者的生命健康。目前,膀胱癌的主要治疗途径为手术切除、辅助免疫治疗及放、化疗,但治疗效果并不理想[22]。
miRNA发挥癌基因或原癌基因活性主要通过与靶基因相互作用实现,对于miRNA靶基因研究一直是miRNA研究中的一个重点和热点,只有明确了miRNA的靶基因,才能揭示肿瘤发生、发展过程中miRNA发挥作用的机制。PTTG1是一种原癌基因,黄庆锋[23]的研究发现,PTTG1在胶质瘤组织中的表达水平随恶性程度增加而升高,胶质瘤细胞系中PTTG1也呈现高表达,其表达与肿瘤分级呈正相关,抑制PTTG1表达可降低胶质瘤细胞的侵袭和迁移能力,miRNA干扰PTTG1表达可明显降低Ki-67及c-myc表达,降低细胞增殖能力。Wang等[24]的研究发现,上调miR-655可通过靶向PTTG1抑制食管癌侵袭能力。以上研究提示miRNA可通过对PTTG1的调节来影响肿瘤细胞的生长。另有研究表明,miR-186-5p可通过靶向抑制PTTG1降低肺腺癌细胞侵袭和迁移能力,抑制细胞EMT发生[25];miR-186可靶向PTTG1参与结直肠神经内分泌肿瘤的浸润和侵袭[26]。
在本实验中,我们收集了50例膀胱癌及癌旁组织标本,qRT-PCR检测显示,miR-186在膀胱癌组织中的表达明显低于癌旁组织,qRT-PCR及Western blotting检测PTTG1表达结果显示,膀胱癌组织中PTTG1表达明显高于癌旁组织。多个靶基因预测软件找到miR-186和PTTG1的结合位点,通过双荧光素酶报告实验证实miR-186和PTTG1确实存在靶向关系。
PCNA是一种分子量为36 kD的蛋白质,在细胞核内合成,并存在于细胞核,在DNA链损伤、修复及复制等多个过程中发挥作用,其表达水平随细胞周期改变而变化,与恶性肿瘤生长、转移、预后等存在密切关系,是一个可反映细胞增殖状态的理想标志物,在多种肿瘤研究中应用广泛[27-28]。
EMT是一种生物学现象,为上皮细胞历经复杂生化改变后获得间质细胞表型的过程,参与肿瘤发生和发展、组织纤维化及慢性炎症。大量研究发现,EMT与肿瘤的侵袭和转移存在密切关系,恶性肿瘤细胞的上皮细胞可通过EMT过程而得到转移能力[29-30]。E-cadherin是一个经典的上皮细胞分子标志物,在维持组织完整性及上皮细胞极性过程中发挥重要作用。E-cadherin表达降低或丢失可使肿瘤上皮细胞极性消失,细胞间黏附性降低,从而使细胞侵袭和转移能力增强。
凋亡过程的启动可对细胞“命运”产生直接影响,在细胞凋亡信号转导过程中,Bcl-2家族成员比例是调控细胞凋亡的一个关键因素,尤其是Bcl-2家族成员抑凋亡基因Bcl-2和促凋亡基因Bax,两者比例是启动细胞凋亡的“分子开关”,Bcl-2和Bax通过形成同源二聚体或异源二聚体发挥细胞凋亡调控作用,Bax与Bcl-2产生异源二聚体时,细胞凋亡受到抑制,当Bax同源二聚体形成时,可引起细胞凋亡发生[31-32]。Bcl-2和Bax常用于细胞凋亡机制的研究,大量研究表明,肿瘤细胞凋亡与Bcl-2和Bax表达密切相关[33-34]。
为了进一步证实miR-186靶向PTTG1对膀胱癌细胞生长的影响,使用miR-186抑制物或miR-186抑制物和miR-186 siRNA同时处理J82细胞,发现抑制miR-186表达可明显增强J82细胞增殖、侵袭和迁移能力,抑制细胞凋亡,上调PCNA、Bcl-2和MMP-2表达,下调Bax和E-cadherin表达,且减弱PTTG1会下调对J82细胞生长的影响。提示miR-186可靶向PTTG1影响膀胱癌细胞生长。通过以上研究,可为膀胱癌早期诊疗、基因治疗及预后判断提供新的策略和理论依据。
猜你喜欢
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
天津医科大学学报(2021年4期)2021-08-21
东坡赤壁诗词(2021年2期)2021-06-01
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
Drug Combination Therapy(2020年1期)2020-02-14
环球时报(2019-01-29)2019-01-29
畅谈(2018年6期)2018-08-28
中国卫生标准管理(2015年25期)2016-01-15
