
信号肽酶复合体催化亚基SPC18 敲除细胞系的建立与鉴定
2019-11-27 08:48张子豪曹涵钰董宁征
中国畜牧杂志 2019年11期
张子豪,李 慧,刘 萌,曹涵钰,董宁征*
(1.江苏省血液研究所,苏州大学医学部,江苏苏州 215123;2.苏州大学唐仲英血液学研究中心,江苏苏州 215123)
乳腺生物反应器是一种转基因动物乳腺个体表达系统,利用哺乳动物乳腺特异性启动子使目的基因在乳腺中特异性表达,通过乳腺的正常分泌获得转基因动物乳汁中的重组蛋白[1]。世界上很多团队已利用该技术生产药用蛋白,Macias 等[2]利用HEK293 细胞生产具有抗凝血活性的人重组蛋白C(rhPC),用于脓毒症的治疗;Gordon 等[3]将组织型纤溶酶原激活剂基因(tPA)插入小鼠乳清酸蛋白基因(WAP)中并成功表达具有天然活性的tPA;Wright 等[4]在绵羊乳汁中将药用蛋白人抗胰蛋白酶(hAAT)表达量提高到35.0 mg/mL;Echelard等[5]通过建立乳腺转基因牛并将人血清白蛋白(hSA)表达量提高到40.0 mg/mL。乳腺生物反应器所获得的重组蛋白大多为分泌蛋白,分泌效率是影响乳腺生物反应器发展的重要因素。
哺乳动物分泌蛋白信号肽由信号肽酶复合体(Signal Peptidase Complex,SPC)水解切除,被切除信号肽的分泌蛋白经过内质网和高尔基体进一步的加工修饰才能顺利分泌到乳汁中,所以SPC 是分泌蛋白合成后蛋白转运过程中的重要调控蛋白。SPC 结构上由5 个亚基构成,分别为催化亚基SPC18 和SPC21,结构亚基SPC12、SPC22/23 和SPC25[6-15]。但迄今尚不清楚催化亚基SPC18 在蛋白分泌中的具体作用,因此深入了解哺乳动物SPC18 的作用机制有助于探究分泌蛋白的合成转运等关键问题。
本实验运用CRISPR-Cas9 基因编辑技术在人胚肾细胞HEK293 的基础上构建SPC18 敲除(KO)细胞系,研究SPC18 缺失对HEK293 细胞行为、细胞增殖以及对内源分泌蛋白表达和分泌的影响,为进一步探究SPC对分泌蛋白胞内转运与分泌机制的影响以及SPC 在乳腺生物反应器中的应用提供了理论基础和实验依据。
1 材料与方法
1.1 主要材料 pGL3-U6-gRNA-Puromycin mutBsaI ACCG gRNA 载体和Cas9 载体由本实验室保存,BsaI 内切酶、T4 DNA 连接酶、NEB Buffer 2.1 购自美国New England Biolabs(NEB)公司,基因组DNA 抽提试剂盒购自德国Qiagen 公司,DEME、Opti-MEM、胎牛血清(FBS)购自美国Gibco 公司,转染试剂PolyJet reagents 购自SignaGen 公司,SPC18 antibody 购自美国Sigma 公司,辣根过氧化酶(HRP)标记的羊抗兔抗体购自Bioworld 公司,MMP9(N-Terminal)antibody购自美国Proteintech 公司,CCK-8、MTT 细胞增殖试剂盒购自碧云天生物技术有限公司,5 ku 分子截留超滤管购自美国Millipore 公司。
1.2 构建稳定敲除SPC18 的HEK293 细胞系
1.2.1 寡核苷酸的合成及载体构建 运用pGL3-U6-gRNAPuromycin mutBsaI ACCG 作 为gRNA 载 体。根 据NCBI GenBank 公布的基因序列(SEC11A:NM-001271921.1),在CRISPR/Cas9 guide RNA design 网 站(https://zlab.bio/guide-design-resources)设 计guide RNA 引 物,序 列:sgRNA-SPC18-F:CCGGCGGCACTAATGATCTGGAAG;sgRNA-SPC18-R:AAACCTTCCAGATCATTAGTGCCG。在引物的5´加BsaI 酶切位点(CCGG-AAAC)。通过退火、酶切、胶回收、酶连反应将sgRNA 片段与载体相连;转化DH5α感受态细胞扩增并测序鉴定。
1.2.2 细胞培养、转染及SPC18 KO 单克隆细胞系的筛选 HEK293 细胞购自美国ATCC 公司,在37℃、5%CO2的培养环境下,用含10% 胎牛血清的DMEM 高糖培养基进行培养[16]。
将上述构建好的载体质粒与Cas9 载体(pSt1374-NNLS-3xflag-Cas9-ZF-TM)按照1∶1 共同转染HEK293 细胞,转染试剂PolyJet 与质粒按照1∶3 浓度混合,6 h 换液,培 养24 h 后 加0.5 µg/mL Puromycin 和16 µg/mL Blasticidin S,筛选3 代之后,分单克隆细胞。筛选获得的单克隆KO 细胞系提取基因组DNA,测序鉴定,同时用Western blotting 检测其蛋白水平。
1.3 细胞形态学实验 培养SPC18 KO 及HEK293 细胞至汇合率为80% 时,加入0.25% 胰蛋白酶进行消化,37℃消化1 min 后细胞变成悬浮状态,加入含有10%胎牛血清的DMEM 终止消化。细胞悬液800 r/min 离心5 min,弃上清,重悬细胞,按照1∶5 的比例将细胞传代。在36、72 h 用倒置显微镜随机选取4 个视野进行观察并拍照。
1.4 MTT 细胞增殖实验 在96 孔板中每孔接种5 000个细胞,加入10 µL MTT 溶液,37℃、5% CO2条件下孵育4 h。每孔加入100 µL Formazan 溶解液,适当混匀溶解,多功能酶标仪在570 nm 检测吸光度(OD),用GraphPad 进行数据分析。
1.5 CCK-8 细胞增殖实验 在96 孔板中每孔接种5 000个细胞,加入10 µL 增强型CCK-8 溶液,37℃、5%CO2条件下孵育2 h。多功能酶标仪在450 nm 检测吸光度(OD),用GraphPad 进行数据分析。
1.6 蛋白质免疫印迹实验(Western blotting) 收集HEK293 和SPC18 KO 细胞,加入含蛋白酶抑制剂的细胞裂解液[50 mmol/L Tris-HCl(pH 8.0),150 mmol/L NaCl,1% Triton X-100(v/v)][16],用二喹啉甲酸(BCA)法测定细胞裂解液蛋白浓度。还原条件下,进行SDSPAGE,200 mA 恒流转膜120 min。5% 脱脂奶粉室温封闭2 h。用0.05% TBST 稀释SPC18 或MMP9 抗体,室温孵育2 h,洗涤后加HRP-羊抗兔抗体,室温孵育2 h,GE AI600 化学发光成像仪显影,用软件ImageJ 对蛋白条带进行统计分析[17]。
1.7 内源分泌蛋白pro-MMP9 的表达及分泌 将HEK293和SPC18 KO 细胞种于培养皿中,至汇合率到80%,加入10 mL 无血清的Opti-MEM 培养基,37℃培养36 h。收集上清及细胞,上清3 000 r/min 离心5 min,5 ku 分子截留超滤管超滤浓缩,同时制备细胞裂解液。用Western blotting 检测MMP9 在HEK293 和SPC18 KO 细胞系中的表达和分泌情况。
1.8 统计分析 用Prism 8(GraphPad)软件分析数据,所有数据均以平均值(Mean)和平均数标准误(SEM)的形式表示。用t检验(Student'st-test)比较2 组变量的差异,P<0.05 时认为具有统计学差异,P<0.01 时认为统计学差异显著。
2 结果
2.1 SPC18 敲除细胞系的建立 设计guide RNA 引物,合成后的寡核苷酸单链通过退火形成双链,然后通过BsaI 酶切位点将其插入pGL3-U6-gRNA 载体(图1)。
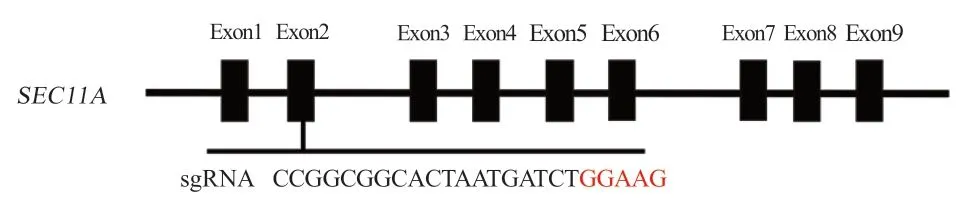
图1 SEC11A 基因敲除质粒sgRNA 设计
测序结果显示,SPC18 KO 与野生型HEK293 细胞相比,2 号外显子缺失GGAAG 5 个碱基(图2-a),导致移码突变,理论上可以使SPC18 蛋白不表达,达到基因敲除的目的。Western blotting 结果也可以看出,在HEK293 细胞中可以检测到18 ku 的蛋白条带,与SPC18 蛋白分子量一致,SPC18 KO 细胞系则检测不到(图2-b)。结果提示,SPC18 KO 细胞系构建成功。
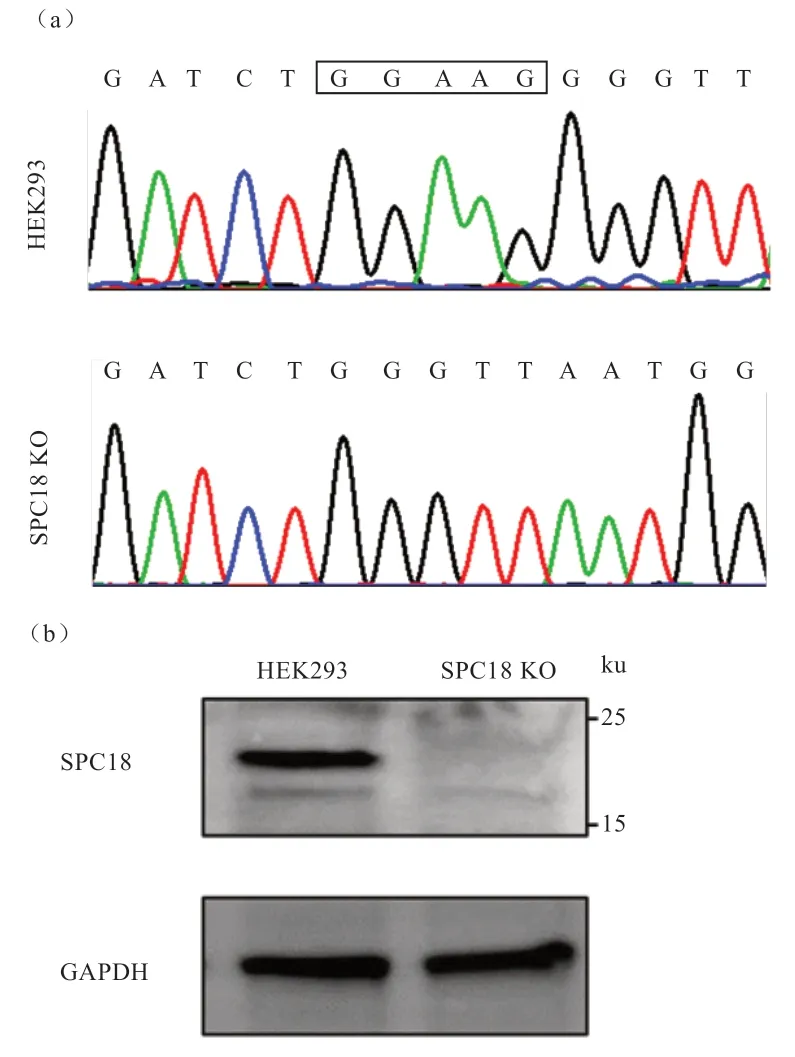
图2 SPC18 KO 细胞系DNA 测序鉴定及蛋白水平鉴定
2.2 SPC18 缺失对细胞形态的影响 由图3 可以看出,传代培养36、72 h 后,HEK293 和SPC18 KO 的细胞形态无显著不同,均显示扁平、多角样贴壁生长。因此,在本实验条件下,信号肽酶复合体催化亚基SPC18 缺失没有导致细胞形态的明显改变。
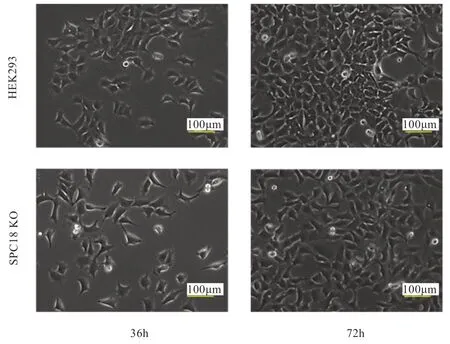
图3 HEK293 和SPC18 KO 细胞形态
2.3 SPC18 缺失对细胞增殖的影响 为了检测SPC18 缺失对HEK293 细胞增殖的影响,采用了MTT 和CCK-8 2 种方法。从图4-a 可以看出,2 组细胞的吸光度值随着培养时间的延长而升高,与对照HEK293 细胞相比,在48 h 时间点SPC18 KO 细胞系的570 nm 吸光度显著降低,提示催化亚基 SPC18 的缺失可能使细胞的增殖能力显著下降。CCK-8 法也得到了与MTT 类似的检测结果(图4-b)。
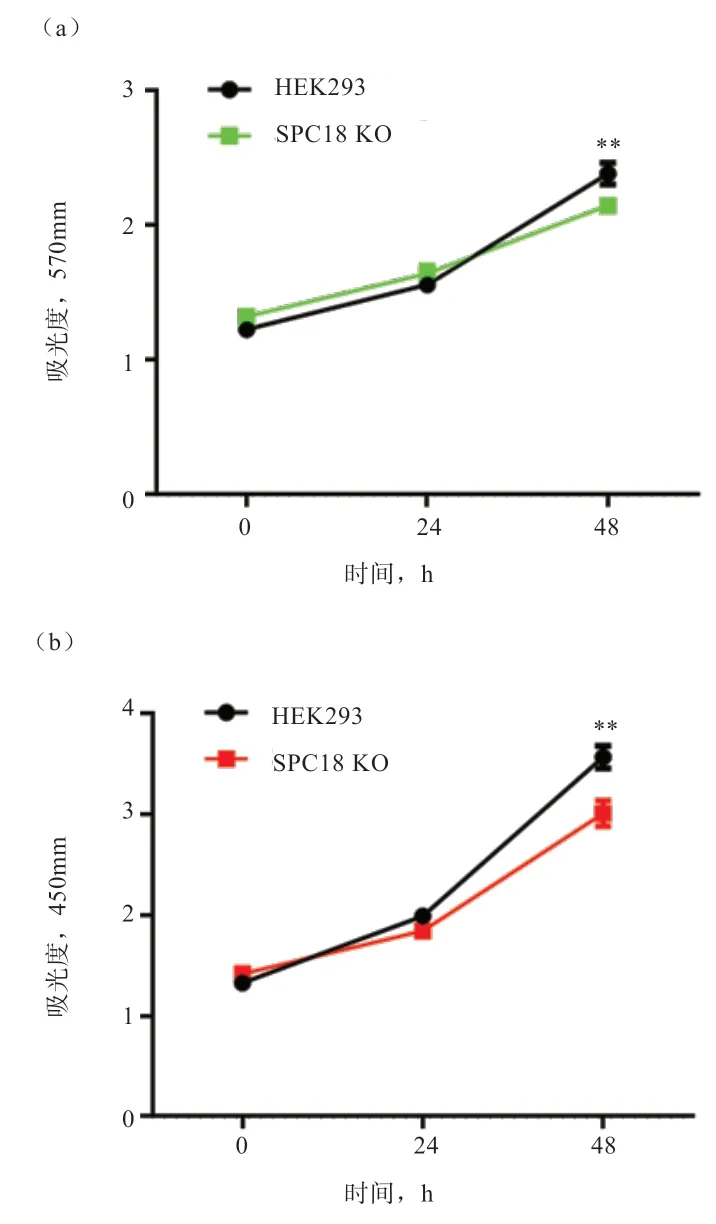
图4 HEK293 与SPC18 KO 细胞增殖曲线
2.4 内源性分泌蛋白pro-MMP9 在SPC18 KO 及HEK293细胞中的表达和分泌 pro-MMP9 蛋白结构包含氨基端信号肽、前导肽、催化结构域、铰链区以及血色素结合蛋白样结构域(图5-a)。在SPC18 KO 细胞中,pro-MMP9 在上清中的表达显著少于HEK293 细胞,而细胞裂解液中pro-MMP9 的表达与对照HEK293 细胞无显著差别(图5-b)。通过Image J 光密度分析pro-MMP9 在上清中的表达量(图5-c),SPC18 KO 细胞上清中pro-MMP9 表达量为野生型HEK293 细胞表达量的17.26%(P<0.01)。结果提示,信号肽酶复合体催化亚基SPC18 缺失后,内源性分泌蛋白pro-MMP9在细胞中的表达不受影响,但削弱了SPC 的功能,导致分泌蛋白分泌受阻。

图5 pro-MMP9 在HEK293 和SPC18 KO 细胞中的表达和分泌
3 讨 论
本实验中,SPC18 的缺失并不影响细胞形态,但细胞增殖能力显著下降。此前有研究报道,催化亚基Sec11(与真核细胞SPC18/21 同源)为酵母细胞生存所必须[18]。酵母细胞SPC 只有1 个催化亚基,而哺乳动物SPC 有2 个催化亚基,所以本研究中1 个催化亚基(SPC18)的缺失仅造成细胞增殖能力下降,同时,分泌蛋白中包含很多细胞生长因子[19-20],其信号肽的切割受SPC 调控,催化亚基SPC18 的缺失可能通过影响细胞生长因子的分泌来造成细胞增殖能力下降,这点还需进一步实验验证。
MMPs 是一类内源性蛋白水解酶家族,以无活性的酶原形式分泌,能够降解细胞外基质中所有蛋白成分。MMP9 主要降解破坏细胞外基质中主要成分IV、V 型胶原和明胶,在细胞分化、组织损伤修复过程中发挥重要作用,参与了胚胎形成、伤口愈合、组织修复等生理过程。SPC18 的缺失并不影响pro-MMP9 在细胞中的正常合成与表达,但显著影响其分泌。此前研究报道,果蝇催化亚基Spc18/21(与哺乳动物SPC18 同源)缺失对个体发育是致命的,缺失后会造成细胞分化不全,相关蛋白分泌受阻,导致翅膀、眼、腿及触角等组织发育障碍[21-22]。在昆虫赤拟谷盗中,催化亚基TcSPC18(与哺乳动物SPC18 同源)缺失会造成肠细胞凋亡,蜕皮蛋白酶分泌受阻,幼虫蜕皮困难[23]。东亚飞蝗中,SPC催化亚基(与哺乳动物同源)的缺失造成蜕皮蛋白酶分泌受阻,肠腔内包括消化酶在内的总蛋白分泌均受阻,胚胎发育紊乱,69% 的幼虫死于蜕皮异常[24]。这与本研究SPC18 对蛋白分泌影响的结论相一致,催化亚基SPC18 对于SPC 发挥作用是至关重要的,SPC18 促进蛋白分泌,缺失后蛋白分泌受阻。
4 结 论
本研究利用CRISPR-Cas9 基因编辑技术,在HEK293 细胞基础上成功构建信号肽酶复合体催化亚基SPC18 敲除细胞系。SPC18 的缺失不改变细胞形态,但抑制细胞增殖。同时,敲除SPC18 后,内源分泌蛋白合成与表达不受影响,但分泌严重受阻,提示SPC18有促进分泌蛋白分泌的作用,为SPC 在动物乳腺生物反应器中调控蛋白分泌奠定了研究基础。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年10期)2021-12-02
科学与生活(2021年16期)2021-11-25
健康之家(2021年19期)2021-05-23
昆明医科大学学报(2021年1期)2021-02-07
浙江医学(2020年19期)2020-10-20
中国生殖健康(2019年5期)2019-01-06
CHINESE JOURNAL OF AERONAUTICS(2016年3期)2016-11-23
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年7期)2015-06-22
