
弱光下不同水深对长喙毛茛泽泻生长及生理特性的影响
2019-11-07 06:55朱光冕曾柏全曾文斌陈介南吕贤良
生态学报 2019年19期
朱光冕,曾柏全,*,曾文斌,沈 燕,詹 鹏,陈介南,吕贤良
1 中南林业科技大学生命科学与技术学院,长沙 410000 2 浙江省丽水市莲都区林业局,丽水 323000
在野外由于季节性降雨,水文条件较为复杂,湿生植物常生活在一个动态的环境中,经历洪水期、常水期和枯水期,水深的周期性变化对植物生长具有重要影响[1- 2]。不同植物对水深变化的响应不同,在水深不适宜时,适应性强的植物可以通过改变其生长情况和渗透调节物质的含量来适应不良的水深环境[3- 4]。光是植物生存和生长发育重要的环境因子之一,过度光照不仅会抑制植物的光合作用,还会导致光合器官的损伤[5]。光照不足则会对植物产生不利的影响,弱光胁迫下植物的形态建成和生理生化过程往往会发生重大改变,其抗氧化物酶[6]、光合生理特性[7]、生物量及叶型特征[8]等生理生态特征都将出现显著性变化。
长喙毛茛泽泻(Ranalismarostratum)为泽泻科(Alismataceae)毛茛泽泻属(Ranalisma),莲座状沼生小草本[9]。该属有2种,在我国仅长喙毛茛泽泻1种,其天然种群仅分布在湖南茶陵,浙江丽水和江西东乡局部生境[10]。由于人类对其生境的破坏和植物本身生存、竞争能力较差,湖南茶陵和江西东乡野外种群已消失[11]。该种在分类学上具有特殊地位,为国家一级保护野生植物[12]。野外调查表明,长喙毛茛泽泻分布于林下遮荫地带的浅水沼泽中,能忍受一定时间的水淹,但不能在深水中长期生存;在光照强度较高的地方,植株叶片往往发生叶片发黄的现象,而光照不足又导致植株矮小、生长缓慢。目前,对长喙毛茛泽泻的研究主要集中在生物学特性[9-11]和繁殖特性[13-14]等方面,但从水深和光照强度对植物的生长和生理特性的影响未见报道。本试验通过在人工气候箱内进行盆栽控水模拟不同水深和光照强度对于长喙毛茛泽泻生长和生理特性的影响,探讨植物野外濒危原因及其对环境的适应能力,为迁地保护和种群恢复提供理论依据。
1 材料与方法
1.1 试验地概况
供试材料采自浙江省丽水市莲都区南明山。试验地位于湖南省中南林业科技大学生命科学与技术学院(27°6′—28°10′N,111°6′—113°10′E),属亚热带季风气候区,年降水量1300—1500 mm,年日照数1450—1686 h,年平均气温15.9—18.2℃。
1.2 试验材料
选用生长旺盛、长势基本一致的长喙毛茛泽泻作为供试材料,平均苗高(6±0.5)cm,叶片数为8叶,叶片呈卵圆形。实验采用盆栽法,盆钵规格为33 cm×19 cm×14 cm(长×宽×高)。
1.3 弱光和水深处理
2017年8月将盆栽苗移入人工气候箱内进行缓苗处理,培养条件为光照时间13 h,白天温度为(25±2)℃,夜间温度为(18±2)℃,昼夜湿度(80±5)%,光照强度(400±10)μmol m-2s-1(在晴天9:00—10:00,采用LI-COR 6400便携式光合作用测定系统测定其光合特性)。实验采用双因素完全随机设计,共设置3个不同水深梯度:0 cm(水面与土面持平且保持表面湿润,形成气生环境)、3 cm(形成挺水环境)、6 cm(形成沉水环境)。根据长喙毛茛泽泻光合作用的特点设置400 μmol m-2s-1(标准光照)和240 μmol m-2s-1(低于其光饱和点,形成低光照)。每个处理设置5个重复。9月上旬开始处理,持续40 d,之后进行相关指标的测定。
1.4 生长和生物量的测定
不同水深和光照强度处理40 d后,从各处理组中随机选取5株植株,测定其叶长(游标卡尺测定)、叶宽、叶柄长度和叶面积(Li-3000型叶面积测定仪,美国LI-COR),并将根部泥土冲洗干净后,按根、茎、叶分别置于80℃烘箱内烘干至恒重,测定其生物量。
1.5 植物生理指标测定
取其叶片用于植物生理指标的测定(包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、丙二醛(MDA)、脯氨酸、可溶性蛋白和可溶性糖)。
其中SOD活性测定采用抑制氮蓝四唑的光还原作用来测定酶的活力[15]、POD活性采用愈创木酚法测定[15]、MDA含量采用硫代巴比妥酸法测定[15]、脯氨酸采用酸性茚三酮法测定[16]、可溶性糖采用蒽酮法测定[15]、可溶性蛋白采用考马斯亮蓝G- 250染色法测定[15]。
1.6 数据处理
数据统计分析采用SPSS 18.0软件,作图采用Origin 2018。平均值间的比较采用单因素方差分析(one-way ANOVA),多重比较采用Duncan新复极差法。水深与光照强度间的交互作用检验采用双因素方差分析(two-way ANOVA)。
2 结果与分析
2.1 不同水深和光照强度对长喙毛茛泽泻各器官生物量及其分配的影响
从表1中可看出,40 d后,在同一光照强度下,不同水深对长喙毛茛泽泻的叶生物量、茎生物量、根生物量和总生物量都具有极显著性影响(P<0.01),随着水深的增加,都呈现出先升高后降低的趋势,并在水深为3 cm、标准光照下叶生物量(0.7 g)、茎生物量(0.2 g)、根生物量(0.24 g)和总生物量(1.15 g)分别出现最大值。在同一水深下,光照强度对植物叶生物量和总生物量具有极显著性影响(P<0.01),对植物茎生物量具有显著性影响(P<0.05),且水深与光照强度的交互作用对植物叶生物量和总生物量也产生显著性影响(P<0.05),在标准光照下植物叶生物量、茎生物量和总生物量都显著高于低光照下植物叶生物量、茎生物量和总生物量。但在同一水深条件下,不同光照强度对于植物根生物量没有显著性影响(P=0.063)。
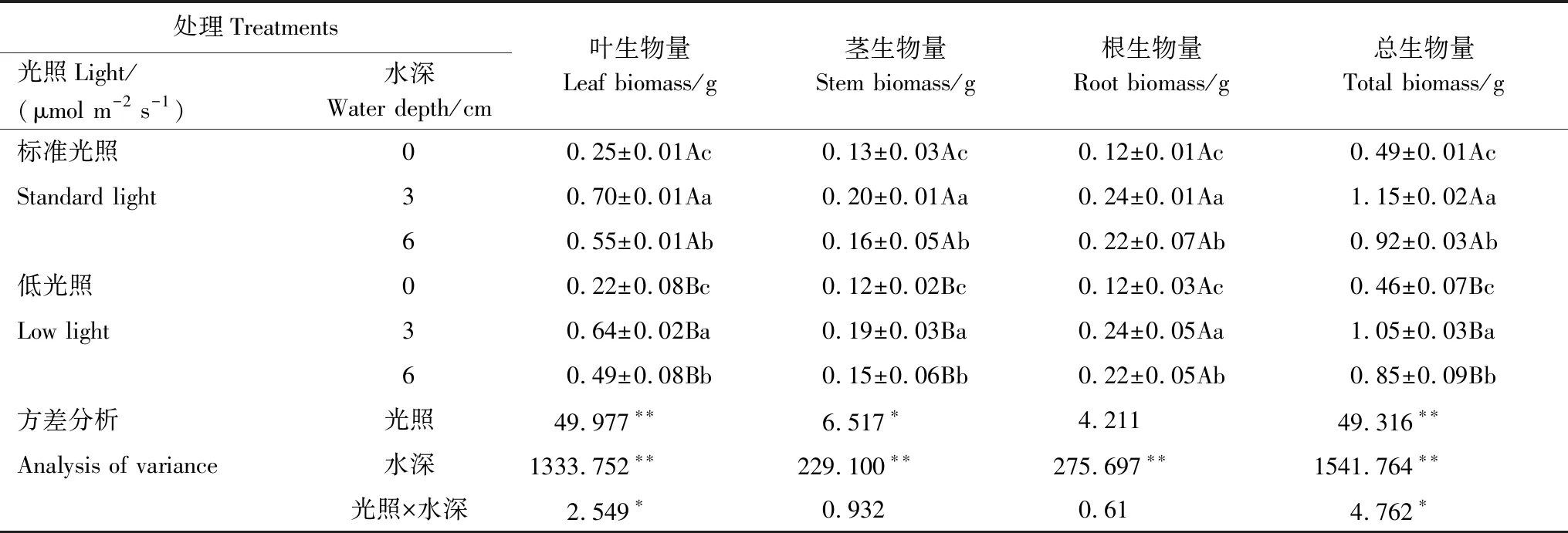
表1 不同水深和光照强度对长喙毛茛泽泻各器官生物量及其分配的影响
同列中标有不同的大写字母表示不同光强间的差异,不同小写字母表示相同光强下不同水深间的差异(P<0.05);*和**表示在0.05和0.01水平上差异显著
2.2 不同水深和光照强度对长喙毛茛泽泻叶形态特征的影响
从表2中可看出,水深与光照强度对长喙毛茛泽泻叶型特征的影响各不相同。在同一光照强度下,不同水深对于植物叶片的叶宽和叶面积都具有极显著性影响(P<0.01),且水深与光照强度的交互作用对植物叶宽和叶面积也产生极显著性影响(P<0.01),随着水深的增加,植株叶宽和叶面积都呈现先升高后降低的趋势,并在水深处理为3 cm时,叶宽(2.16 cm)和叶面积(3.24 cm)出现最大值。水深与光照强度对植物叶长没有显著性影响(P=0.061)、(P=0.487),但水深与光照强度的交互作用对植物叶长有极显著性影响(P<0.01)。在同一水深条件下,光照强度对植物叶柄长度没有显著性影响(P=0.859),但在同一光照强度下,水深对于植物叶柄长度有显著性影响(P<0.05),植株的叶柄长度随水深的增加而增加。
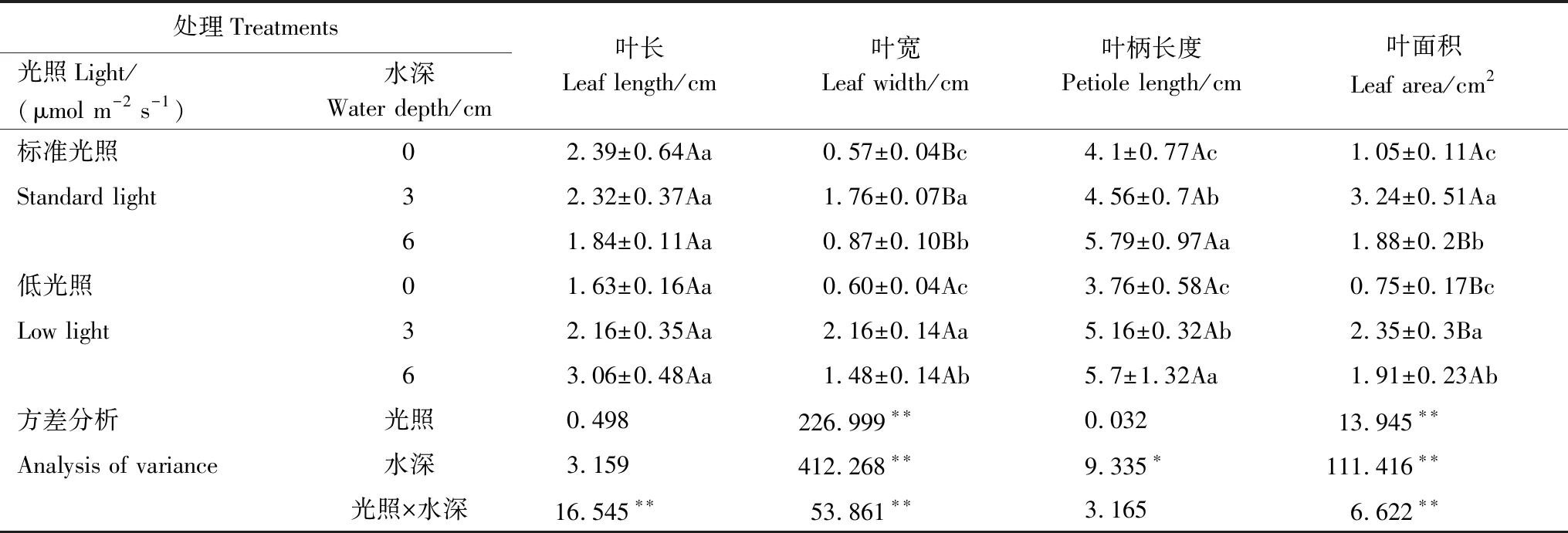
表2 不同水深和光照强度对长喙毛茛泽泻叶形态特征的影响
2.3 不同水深和光照强度对长喙毛茛泽泻生理指标的影响
如图1所示,在水深为3 cm、标准光照下长喙毛茛泽泻叶片中SOD活性最高(124.78 U g-1min-1)。在同一光照强度下,水深为0 cm和6 cm时植株叶片SOD活性差异不大。在同一水深条件下,不同光照强度对植物叶片中SOD活性变化没有显著性影响(P=0.06)。
在同一光照强度下,不同水深对于植物叶片中POD活性具有极显著性影响(P<0.01),随着水深的增加,呈现先升高后降低的趋势,水深为3 cm时叶片中POD活性最高(175.36 U g-1min-1),随着水深的变化POD活性有所下降。在同一水深下,不同光照对植物叶片中POD活性也具有极显著性影响(P<0.01),标准光照下的POD活性显著高于低光照下POD活性。
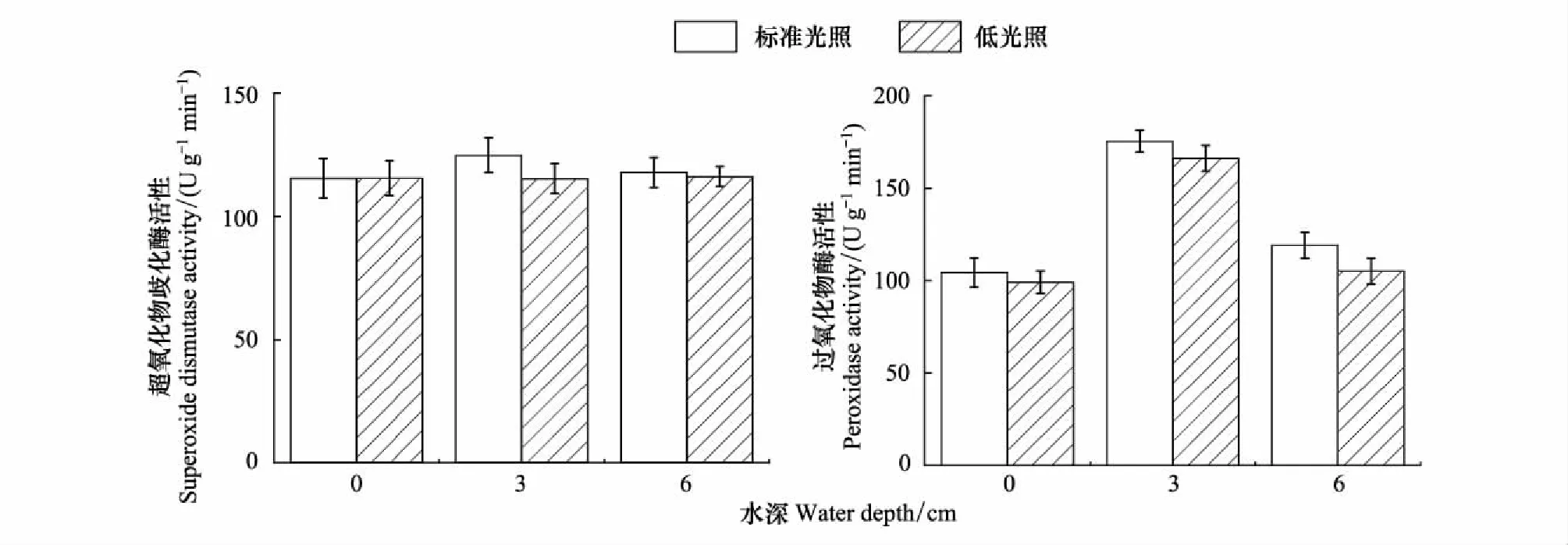
图1 不同水深和光照强度对长喙毛茛泽泻抗氧化物酶活性的影响Fig.1 Effects of water depth and light intensity on activity of antioxidant enzyme in Ranalisma rostratum
如图2所示,在同一光照强度下,不同水深对于植物叶片中可溶性蛋白具有极显著性影响(P<0.01),随着水深的增加呈现先升高后降低的趋势,水深为3 cm时叶片中可溶性蛋白含量最高(5.94 mg/g),随着水深的变化可溶性蛋白有所下降。在同一水深下,不同光照强度对植物叶片中可溶性蛋白的含量变化具有显著性影响(P<0.05),标准光照下可溶性蛋白的含量显著高于低光照下可溶性蛋白的含量。
在同一水深条件下,光照强度对植物叶片中可溶性糖的含量变化具有极显著性影响(P<0.01)在标准光照下的可溶性糖的含量显著高于低光照下可溶性糖的含量。在同一光照强度下水深的变化对植物叶片中可溶性糖的含量变化有显著性影响(P<0.05),随着水深的增加呈现先升高后降低的趋势,在水深为3 cm时植物叶片中可溶性糖的含量最高(9.67 mg/g)。
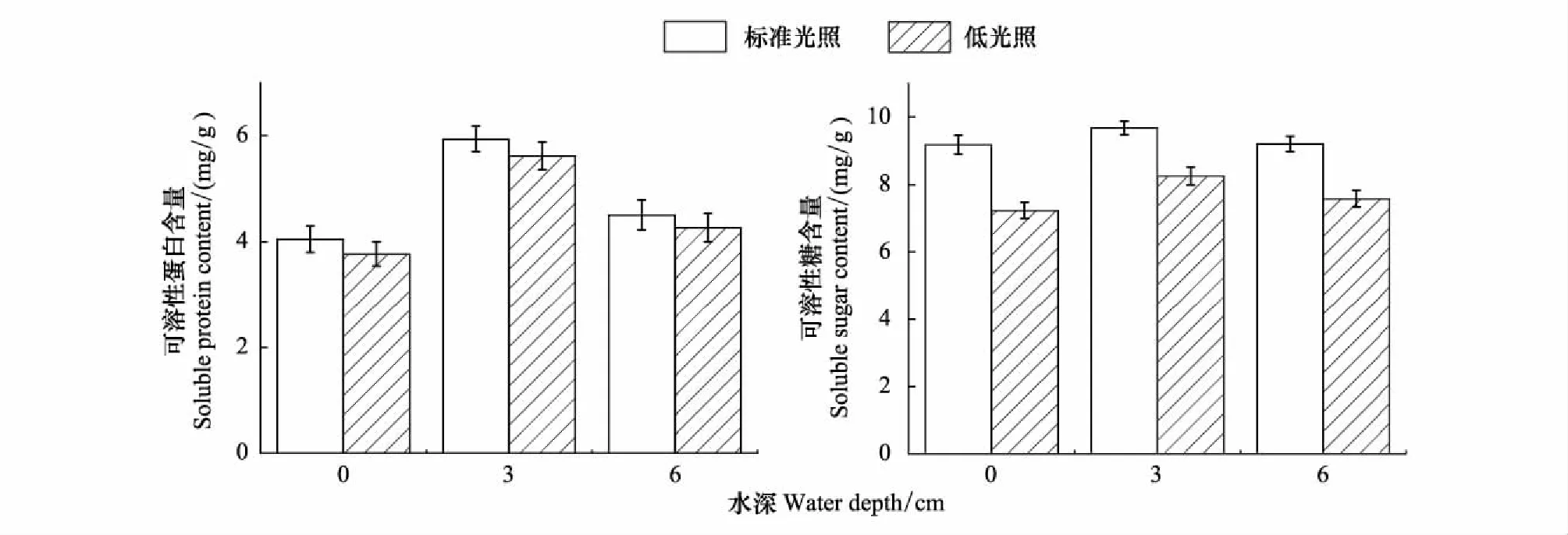
图2 不同水深和光照强度对长喙毛茛泽泻可溶性蛋白和可溶性糖含量的影响Fig.2 Effects of water depth and light intensity on content of soluble protein and soluble sugar in Ranalisma rostratum
如图3所示,在水深为0 cm时,不同光照强度对植物叶片中MDA的含量没有显著性差异(P=0.857),在水深为3 cm时,标准光照下MDA的含量要显著高于低光照下MDA含量(P<0.05),而在水深为6 cm时,情况则相反,低光照下MDA的含量显著高于标准光照下MDA的含量。
在同一水深条件下,光照强度对植物叶片中脯氨酸的含量变化具有极显著影响(P<0.01)。标准光照下脯氨酸的含量显著高于低光照下脯氨酸的含量。在同一光照条件下,水深的升高对于植物叶片中脯氨酸含量的变化有极显著影响(P<0.01),随着水深的升高,脯氨酸的含量呈现下降的趋势,水深为0 cm时脯氨酸的含量显著高于水深为3 cm和水深为6 cm时脯氨酸的含量。
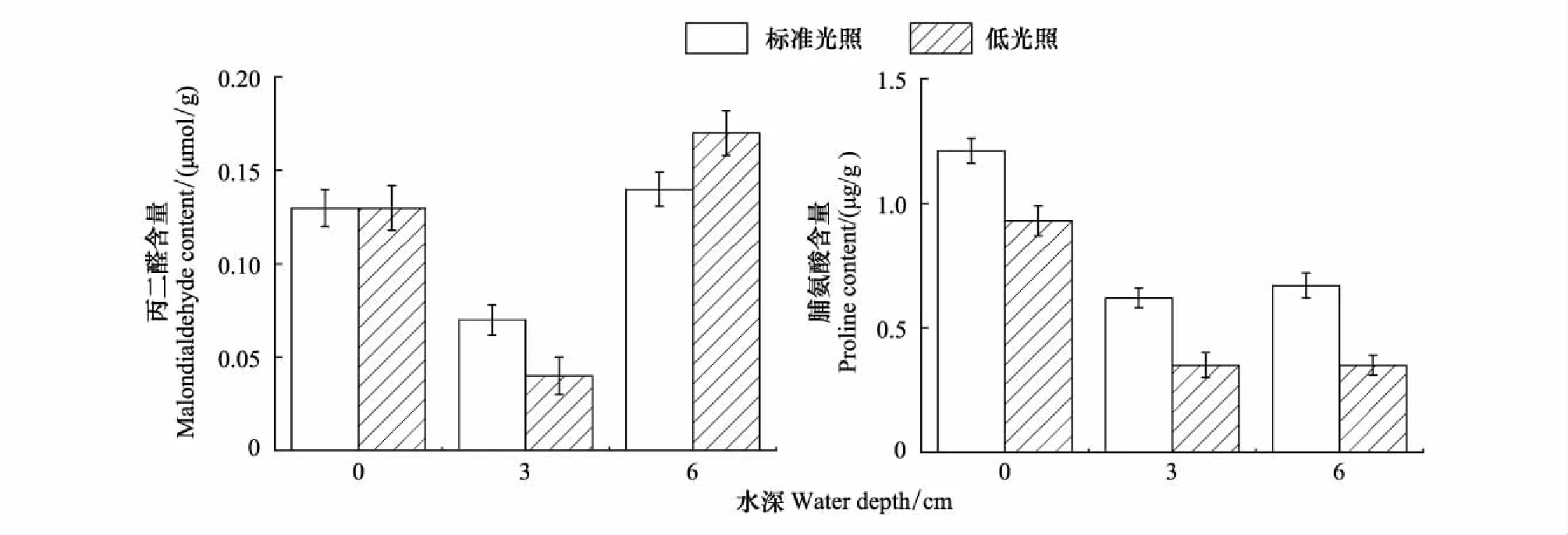
图3 不同水深和光照强度对长喙毛茛泽泻丙二醛和脯氨酸含量的影响Fig.3 Effects of water depth and light intensity on content of malondialdehyde and proline in Ranalisma rostratum
3 讨论
本研究通过弱光和水深双重处理,发现长喙毛茛泽泻的生长和生理特性受弱光和水深的双重影响。生物量作为植物对环境适应性的直接表现,在适宜的水深条件下植物的生物量往往会发生显著性的提高[17-18]。在同一光照条件下,长喙毛茛泽泻的叶生物量、茎生物量、根生物量和总生物量随水深的增加都呈现先升高后降低的趋势,并在水深为3 cm时出现最大值。这与其他水生植物生物量的变化趋势相同[19],过浅和过深的水深都将抑制长喙毛茛泽泻的生长,这表明其生长“机会窗”较狭窄[20],植株对于小范围内的水深变化反应较敏感,且植株不耐干旱,但能忍耐一定程度的水淹。当光照强度降低时,长喙毛茛泽泻的叶生物量、茎生物量和总生物量分别出现下降的趋势,而根生物量则没有变化,这可能是由于长喙毛茛泽泻为多年生的湿生小草本,为适应植株在水环境中的生长,植株的根系发育微弱,所以光照强度对于植株根生物量的影响不显著。长喙毛茛泽泻为叶基生的湿生植物,其地上部分主要为叶。因此,水深和光照对于植株叶生物量有显著性影响,适宜的水深和光照强度使植物叶生物量不断增大,从而使植株总生物量也出现显著性变化。
光照强度和水深不仅对于植株的生物量及其分配有很大影响,也对植株的叶型变化有很大的影响,叶型指标的变化体现了植株对环境的适应能力[21]。在本研究中,不同水深和光照强度对植株的叶长并无显著性影响,但植株的叶宽受水深和光照强度的双重影响。在同一光照条件下,随着水深的增加,长喙毛茛泽泻的叶宽和叶面积呈现先升高后降低的趋势,由于其叶长变化不显著,而叶宽出现显著性变化,这导致长喙毛茛泽泻叶型出现显著性变化,在气生和沉水环境下,其叶片呈披针形。据王建波等[11]观察,长喙毛茛泽泻在沉水情况下,叶片呈披针叶形,这与本研究结果一致,但其观察到长喙毛茛泽泻在气生状态时,叶片呈卵圆形,这与本研究中结果不一致。这可能是由于,在本研究中长喙毛茛泽泻受弱光和浅水双重胁迫,导致植株整体叶片偏小,植株光合作用减弱,致使植株生长不足,叶片呈现披针形。而叶面积的变化也主要源于植株叶宽的变化。在本研究中,长喙毛茛泽泻叶柄长度不受光照强度的影响,但受水深变化的影响并随水深的增加而增加,这可能是因为长喙毛茛泽泻为适应水深变化,使叶片始终处于水面之上,而发生的适应性改变[22]。
在不同水深和光照条件下长喙毛茛泽泻叶片中生理指标的变化进一步证实了上述结果,SOD和POD作为植物体内的抗氧化物酶,起着清除体内自由基和过氧化物,缓解逆境损伤的作用[23-24]。可溶性蛋白作为植物体内重要的渗透调节物质,对于植物缓解逆境损伤有重要作用[25]。可溶性糖作为植物体内重要的非结构性贮藏碳水化合物,不仅对于植物抵抗逆境环境有重要作用,同时也在一定程度上表征了植物固定能量的水平[26-27]。MDA作为植物膜脂过氧化的产物,是植物遭受逆境胁迫的重要检测指标,逆境对于植物的胁迫越严重,MDA的含量越高[28-29]。脯氨酸作为植物体内重要的游离氨基酸,对于植物抵御水分亏缺和盐分胁迫等逆境环境具有重要作用[30-31]。
在曹永慧等[32]研究中,抗氧化物酶活性和渗透调节物质含量的提高,可以提高幼苗对弱光的适应性。而本研究中,各处理组中SOD活性没有显著性差异,而POD活性和可溶性蛋白的含量,在同一光照条件下,都呈现先升高后降低的趋势,并在水深为3 cm、标准光照下含量最高,这说明长喙毛茛泽泻体内抗氧化物酶系统和渗透调节系统没有对水深变化和光照强度的降低产生积极的响应,以适应不同水深和光照强度。在各处理中,水深变化对植株体内可溶性糖含量的变化有显著性影响。且同一水深条件下,不同光照强度对植株可溶性糖含量变化有显著性影响,光照强度的降低显著降低了可溶性糖含量,这与唐中华等[28]研究结果相似,弱光显著性的降低了植物能量的固定。在本研究中,同一光照条件下,随着水深的增加MDA含量呈现先降低后升高的趋势,这有别于其他耐淹植物,MDA含量越高,表示膜脂破坏程度越高,植株遭受胁迫越严重。这与上述生物量和叶型指标结果相似,长喙毛茛泽泻在水深为0 cm和6 cm时,遭到了不同程度的胁迫,且弱光加剧了长喙毛茛泽泻的水淹胁迫。在水深为0 cm时,长喙毛茛泽泻处于水分亏缺的状态,气生环境不利于其生长,植株为抵御不良生境,故脯氨酸含量较高。
4 结论
在同一光照条件下,随着水深的增加其叶生物量、茎生物量、总生物量、叶宽、叶面积、POD活性、可溶性蛋白和可溶性糖含量都呈现先升高后降低的趋势;MDA含量呈现先降低后升高的趋势;叶柄长度随水深的增加而增加;脯氨酸含量随水深的增加而减少。在同一水深条件下,随着光照强度的降低其叶生物量、茎生物量、总生物量、POD活性、可溶性蛋白、可溶性糖和脯氨酸的含量都有所下降。
在野外,长喙毛茛泽泻种子发芽率偏低、幼苗存活率不高、种群扩散困难以及生境的人为破坏都是导致其濒危的重要原因[9]。根据本研究发现长喙毛茛泽泻对光照强度和水深变化较敏感,对动态环境适应性较差。一定范围内水深变化和上层植物的荫蔽可能也是导致其濒危的重要原因。
猜你喜欢
河北水利(2022年10期)2022-12-29
海洋通报(2022年4期)2022-10-10
中老年保健(2022年2期)2022-08-24
海洋信息技术与应用(2022年1期)2022-06-05
中国机械工程(2022年8期)2022-05-09
中国民族民间医药(2022年4期)2022-03-24
儿童时代·幸福宝宝(2021年1期)2021-03-29
贵州农业科学(2020年10期)2020-11-22
小资CHIC!ELEGANCE(2019年40期)2019-12-10
浙江国土资源(2015年4期)2015-05-09
