
Zn对水稻吸收转运Cd的影响
2019-10-23 06:00:54陈仕淼辛子兵陆覃昱郑富海
农业环境科学学报 2019年10期
陈仕淼,辛子兵,陆覃昱,郑富海,何 冰*
(1.广西大学,广西农业环境与农产品安全重点实验室,南宁 530005;2.广西壮族自治区中国科学院广西植物研究所,广西喀斯特植物保育与恢复生态学重点实验室,广西 桂林 541006;3.广州市林业和园林科学研究院,广州 510405)
锌(Zn)是植物所需的必要元素,镉(Cd)与Zn同族,具有相近的化学性质。多数研究发现,Zn可减少植物对Cd的吸收[1-4],减轻Cd毒害进而缓解农作物减产[2,5-6]。Zn的加入可以有效抑制水稻Cd的吸收和向上运输[7]。施用100 mg·L-1纳米ZnO,水稻穗和根中的Cd含量下降了30%和31%[8]。研究表明Zn可以显著降低水稻根系中Cd含量,提高水稻地上部Cd含量,促进Cd向上运输[9]。游离的Zn2+浓度在亚植物毒性水平(10-7.6至10-6.1)不会抑制水稻对Cd的吸收[10]。因此Zn对水稻Cd吸收转运的影响机制尚不清楚。
植物对重金属的吸收及转运过程包括质外体途径和共质体途径。水稻根系铁膜以及细胞壁是阻隔质外体中重金属进入共质体的主要屏障。根表铁膜增加了水稻对Zn的吸收[11],水稻对Zn的吸收与水稻根系铁膜厚度成正比,而水稻对Cd的吸收与根系铁膜厚度成反比[12]。有研究指出,水稻根部致密的根表铁膜阻遏了水稻对Cd的吸收,减轻了Cd对水稻幼苗的毒害[13-14]。水稻根系中有30%以上的Cd被细胞壁所阻隔,使Cd不能进入细胞并向上转运[15-17]。细胞壁组分影响水稻对Cd和Zn的吸收,有研究认为不同品种的油菜对Cd的吸收及耐性差异来源于细胞壁组分差异[18]。果胶是影响细胞壁对重金属吸收的重要因素之一,有研究表明果胶促进芹菜细胞壁对Cd的吸附[19],也有研究表明,果胶抑制水稻对Zn的吸附,而半纤维素则促进细胞壁吸收Zn[20]。Yang等[21]对拟南芥细胞壁组分的研究表明,半纤维素1(HC1)是拟南芥细胞壁组分中Al吸收的主要成分。Zhu等[22]研究表明,Cd2+固定在根细胞壁组分HC1中。Li等[23]对东南景天细胞壁组分的研究表明,高Cd条件下东南景天果胶和半纤维素2(HC2)含量显著增加,但细胞壁中Cd含量却显著降低。Cd、Zn处理影响细胞壁组分,果胶与半纤维素在柳树根细胞壁吸附Cd中发挥重要的作用[24]。Zn缺乏会导致甜茶细胞壁中果胶、HC1与HC2的下降[25]。Cd可诱导水稻品种日本晴细胞壁中果胶含量增加,使果胶甲酯酶活性提高[26]。综上所述,水稻根表铁膜和细胞壁对阻碍水稻吸收Cd和Zn具有重要作用。细胞壁中HC1能促进重金属在细胞壁中的积累,而果胶及HC2抑制细胞壁对重金属的吸收。我们试图分析Zn是否通过改变根表铁膜和细胞壁成分从而影响Cd的吸收及转运,探究水稻中Zn与Cd的相互作用规律,为复合污染农田中水稻生产的农产品质量安全控制提供理论依据。
1 材料与方法
1.1 实验设计
供试水稻品种为盐两优888(水稻Cd高积累品种)。水稻种子采用常规方法播种催芽,待水稻幼苗生长至三叶一心时移栽至3 L塑料桶中,使用国际水稻营养液培养,营养液成分如表1。保持营养液pH 5.0,每3 d更换一次,待幼苗生长至四叶一心时进行处理。实验设 CK(不添加 Cd、Zn)、Cd(5 μmol·L-1CdCl2)及 Cd+Zn(5 μmol·L-1CdCl2+30 μmol·L-1ZnCl2)3个处理,每个处理4次重复。每3 d处理1次,共处理3次,9 d后采样。

表1 国际水稻研究所常规营养液成分[27]Table 1 International rice research institute conventional nutrient solution composition[27]
1.2 测定项目及方法
1.2.1 植株生物量及Cd、Zn含量分析
植株分地上部和根部两部分,洗净,20 mmol·L-1EDTA-Na2浸泡20 min,去离子水冲洗3次,擦干,105℃杀青30 min,65℃烘干至恒质量,称干质量。取烘干的植物样品,粉碎,过100目筛,准确称取0.300 g样品,加入优级纯硝酸8 mL,微波消解法消解(Mars6 CEM,美国),原子吸收分光光度法测定Zn、Cd含量(PinAAcle 900T珀金埃尔默,美国)。金属积累量的计算方式为:
Zn、Cd积累量(mg·plant-1,以干物质计)=生物量(g·plant-1)×金属含量(mg·kg-1)。
1.2.2 根表铁膜提取及金属含量测定
根表铁膜的提取采用连二亚硫酸钠-柠檬酸钠-碳酸氢钠(DCB)法[28],待测液用原子吸收分光光度法测定Fe、Mn、Cd、Zn含量,按照Taylor等[29]方法计算铁膜厚度。
1.2.3 细胞壁组分提取、金属含量测定及吸附解析特性分析
细胞壁提取参照唐剑锋等[30]的方法,将提取细胞壁分成两份:一份用于组分分析,细胞壁组分提取及测定参照Yang等[31]的方法,果胶、HC1和HC2中半乳糖醛酸含量的测定采用分光光度法;一份用于细胞壁对Cd和Zn的吸附解吸动力学分析,参照何冰[32]的方法。
细胞壁组分中半乳糖醛酸的测定方法为:取已提取的果胶、HC1和HC2待测液各1 mL,3组分别以三者的提取液1 mL作为空白对照进行比色,加入5 mL四硼酸钠/硫酸溶液,混匀,沸水浴20 min,冷却至室温,在520 nm比色。再加入100 μL 1.5 mg·mL-1间羟基联苯溶液,混匀,5 min后520 nm再读取一次,吸光度的增加量表示各组分中糖醛酸的含量。
1.3 数据统计及分析
实验数据采用SPSS 22.0进行单因素方差分析,Duncan法多重比较,使用Origin 9.0对细胞壁吸附解吸数据进行非线性拟合及作图。
2 结果与分析
2.1 不同处理对水稻幼苗生长的影响
如表2所示,单Cd处理及Cd+Zn处理显著提高了水稻幼苗根部的生物量,与CK相比二者的增幅分别为27.22%和22.36%。地上部Cd+Zn处理显著降低了水稻幼苗地上部生物量,而单Cd处理与CK没有显著差异。结果说明在供试范围内,单Cd处理显著促进水稻根系生长,但对地上部无影响;而Cd+Zn处理,显著抑制地上部生长。

表2 水稻地上部及根部生物量(g·plant-1)Table 2 Shoot and root biomass of rice seedling(g·plant-1)
2.2 不同处理对水稻幼苗Cd、Zn含量的影响
各处理中,根部Cd含量均远高于地上部Cd含量(图1),相较于单Cd处理,Cd+Zn处理显著提高了水稻幼苗地上部及根部的Cd含量,其根部的增幅为单Cd处理的8.28%。相较于根部,Cd+Zn处理条件下地上部的Cd含量增加更为明显,增幅为单Cd处理的223%。各处理在积累量上表现出相同的趋势(表3),但由于Cd+Zn处理后,地上部的生物量有所下降,地上部Cd积累量的增幅有所下降,为单Cd处理的181%。

图1 水稻地上部及根部Cd含量Figure 1 Shoot and root Cd concentration of rice seedling
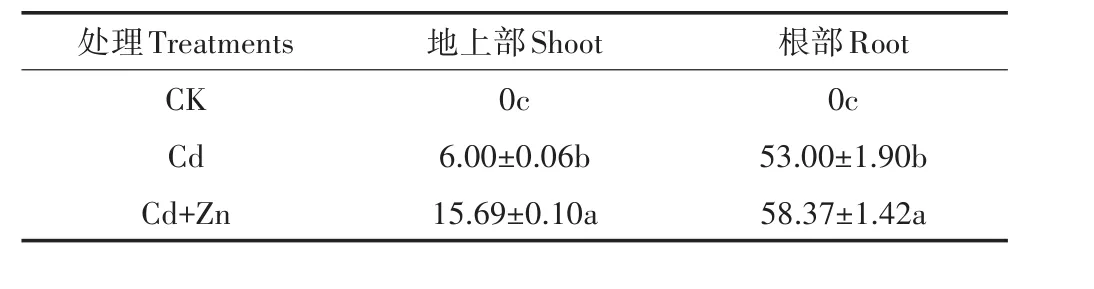
表3 水稻地上部及根Cd积累量(μg·株-1)Table 3 shoot and root Cd accumulation of rice seedling(μg·plant-1)
Cd+Zn处理各部分Zn含量及总积累量均远高于CK,其地上部相较于单Cd处理增幅分别为270%和222%,而其根部Zn含量及总积累量则分别是单Cd处理的19.46倍及18.76倍。值得注意的是,单Cd处理的地上部Zn含量(图2)及地上部Zn积累量(表4)均低于CK,表明Cd的加入会抑制水稻对Zn的吸收,暗示Zn、Cd间存在拮抗关系。
2.3 不同处理对水稻幼苗根表铁膜成分的影响
不同处理下水稻幼苗形成的根表铁膜成分分析结果表明(表5):与CK相比,单Cd处理降低了铁膜中Fe、Mn的含量,而Cd+Zn处理中的Fe、Mn含量与CK无显著差异。单Cd处理降低了根表铁膜中Zn的含量,Cd+Zn处理的根表铁膜中Cd含量比单Cd处理增加29.99%。

图2 水稻地上部及根部Zn含量Figure 2 Shoot and root Zn concentration of rice seedling

表4 水稻地上部及根部Zn积累量(μg·株-1)Table 4 Shoot and root Zn accumulation of rice seedling(μg·plant-1)
2.4 不同处理对水稻幼苗细胞壁组分及Cd吸附解析的影响
研究表明[33],半乳糖醛酸与细胞壁组分之间关系相对稳定,半乳糖醛酸的含量能表征细胞壁组分的多少。细胞壁作为Cd进入共质体的质外体屏障,对Cd具有拦截、阻隔的作用。试验结果表明(图3),相较于单Cd处理,Cd+Zn处理根系细胞壁中果胶糖醛酸含量下降11.58%,HC1糖醛酸含量下降11.45%,HC2糖醛酸含量增加14.82%。

图3 水稻幼苗根细胞壁组分中半乳糖醛酸含量Figure 3 Galacturonic acid concentration in different composition of root cell wall of rice seedling
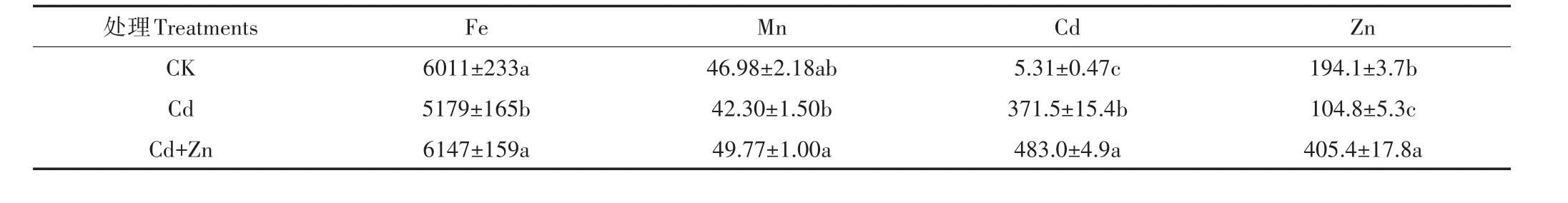
表5 水稻根表铁膜成分(mg·kg-1)Table 5 Iron plaque element of rice seedling(mg·kg-1)
根细胞壁Cd吸附时间动力学曲线如图4,根细胞壁对Cd的吸附随着处理时间的延长而不断增加。在1~150 min,两个处理的细胞壁吸附量迅速增加,单Cd处理的细胞壁吸附量增加幅度大于Cd+Zn处理,在180 min左右,两个处理的Cd吸附进入平台期,细胞壁对Cd的吸附增速极其缓慢。一级扩散方程的物理含义为吸附解析关系只与扩散机理相关,细胞壁对离子的吸附解析速率与未被吸附解析量的一次方成比例[34-35]。采用一级扩散方程对细胞壁吸附Cd的时间动力学曲线进行拟合,单Cd处理及Cd+Zn处理的R2均为0.99(表6)。由一级扩散方程参数可知,单Cd处理的饱合吸附量(Qm)比Cd+Zn处理高13.42%,说明单Cd处理的细胞壁组分具有更高的Cd吸附能力。
根细胞壁Cd解吸时间动力学曲线如图5,根细胞壁对Cd的解吸随着处理时间的延长而不断增加。在1~120 min,两个处理的细胞壁解吸量迅速增加,单Cd处理的细胞壁解吸量增加幅度大于Cd+Zn处理。在360 min左右,两个处理的Cd解吸进入平台期,细胞壁对Cd的解吸增速极其缓慢。采用一级扩散方程对细胞壁解吸Cd的时间动力学曲线进行拟合,单Cd处理及Cd+Zn处理的R2分别为0.94和0.96(表7)。单Cd处理的Qm与Cd+Zn处理差异不大,说明单Cd处理和Cd+Zn处理的细胞壁组分Cd解吸能力差异较小。

图4 不同处理根细胞壁Cd吸附动力曲线Figure 4 Cd adsorption kinetic curve of cell wall from root of different treatments

表6 吸附动力学一级扩散方程拟合参数Table 6 The adsorption fitting parameters of first-order diffusion kinetic equation

图5 不同处理根细胞壁Cd解吸动力学曲线Figure 5 Cd desorption kinetic curve of cell wall from root of different treatments

表7 解吸动力学一级扩散方程拟合参数Table 7 The desorption fitting parameters of first-order diffusion kinetic equation
3 讨论
实验结果表明,在供试范围内,Zn、Cd共同处理对水稻幼苗地上部Cd积累具有显著的促进作用,同时抑制水稻幼苗地上部的生长,这可能是Cd和Zn相互作用的结果。
3.1 Zn对Cd胁迫下水稻生长和吸收Cd的影响
Cd对水稻的生长及营养代谢具有明显的抑制作用[36],但低剂量的Cd2+会刺激植物的根系生长[37-38]。多数研究表明,Zn在植物应对非生物胁迫中起到积极的作用,Zn可以通过稳定、保护细胞膜,缓解Cd造成的氧化胁迫,从而减缓Cd对水稻生长的抑制[6]。实验结果表明单Cd处理与Cd+Zn处理的根部生物量均高于CK,而地上部生物量单Cd处理与CK持平,Cd+Zn处理略小于CK。由于在Cd+Zn处理中,根部Zn含量约为地上部的两倍,而根未表现出明显的毒害效应,所以推测过高的Zn含量可能是导致Cd+Zn处理地上部生物量下降的主要原因。
Cd与Zn属同主族元素,化学性质相近,多数研究认为,Cd、Zn共用多数吸收、转运通道[39-40],故在植物的吸收上,两者表现为拮抗作用[1-3]。而一些研究结果却与之相反。有研究表明[41],当同时施用Zn和Cd时,10 μmol·L-1Cd+50 μmol·L-1Zn的组合施用增加了菜豆地上部及根部的Cd和Zn含量。实验结果表明,与CK相比,单Cd处理根部与地上部Zn含量均略有下降,表明在Zn低浓度条件下,Cd与Zn表现为拮抗作用,而Cd+Zn处理根部与地上部Cd含量与积累量均高于CK,表明在Zn浓度≥30 μmol·L-1的高浓度条件下,Zn促进了Cd的吸收与转运。
3.2 高浓度Zn促进水稻吸收Cd的机制
根表铁膜被认为可以吸附和共沉淀重金属离子,降低其生物有效性从而减少根系对有害离子的吸收[42-43]。同样,有研究指出,不同水稻品种的Cd吸收差异与各个品种根表铁膜的形成量密切相关[44-45]。实验结果表明,相较于单Cd处理,Cd+Zn处理显著提高了根表铁膜中的Fe含量及Mn含量(与CK持平),更有利于铁膜及锰膜的形成[46],从而进一步促进了Cd在铁膜中的络合与共沉淀,表现为Cd+Zn处理中根表铁膜Cd含量显著高于单Cd处理。值得注意的是,相较于CK,单Cd处理显著降低了根表铁膜中Fe及Mn含量,表明在供试范围内,Cd抑制水稻根表铁膜的形成,而Zn则可以缓解这一过程。另外,我们发现,虽然Cd+Zn处理铁膜中Cd含量高于单Cd处理,但相较于单Cd处理,其根部Cd含量也较高,暗示Cd+Zn根部Cd主要积累于根表铁膜中,共质体中Cd含量较小。
细胞壁是金属离子跨膜进入原生质体的最后屏障。植物细胞壁的主要成分包括纤维素和基质多糖,而基质多糖则包括果胶、HC1、HC2[33,47],其中含有大量带负电荷的基团如羧基、醛基、氨基以及磷酸基等,通过吸附、络合、沉淀等作用阻隔重金属离子向细胞内转运[48]。Cd+Zn处理中,果胶与HC1含量均低于单Cd处理,而果胶与HC1被认为是影响Cd吸附的重要成分[49-50]。在吸附解吸实验中,Cd+Zn处理Cd吸附总量相较于单Cd处理更低而解吸总量更高,表明Cd+Zn处理的细胞壁对Cd的拦截效应更弱,由此推测Cd+Zn处理导致细胞壁组分变化,降低果胶与HC1比例,进而减少细胞壁对Cd的拦截是导致水稻地上部Cd吸收增加的原因之一。
4 结论
根表铁膜是水稻吸收重金属进入水稻根系的第一道屏障。综上所述,Zn缓解Cd对水稻根表铁膜形成的抑制效应,促进Cd在根表铁膜的沉积,降低根对Cd的吸收,缓解Cd对水稻根系生长的影响。Cd一旦进入根系,Zn可以通过降低根系细胞壁中的果胶与HCl含量,减少细胞壁对Cd的吸附容量,提高细胞壁的解吸容量,促进Cd向地上部的运输,由此推测,高浓度Zn处理并不能抑制Cd在地上部的积累及其对地上部的毒害。对于Zn如何改变细胞壁成分(果胶和HC1)的合成与分解代谢的相关研究,将能更好地解释高浓度Zn促进水稻吸收转运Cd的机理。
猜你喜欢
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19 08:39:24
金属加工(热加工)(2020年12期)2020-02-06 05:59:26
中成药(2018年6期)2018-07-11 03:01:12
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
腹腔镜外科杂志(2016年11期)2016-06-01 12:10:08
中国粮油学报(2016年1期)2016-02-06 02:17:06
磁共振成像(2015年9期)2015-12-26 07:20:31
实用手外科杂志(2015年2期)2015-08-28 09:50:58
天然产物研究与开发(2014年8期)2014-04-27 14:16:35
