
防城河口湾鱼类群落结构及其与环境因子关系研究
2019-10-14 03:17何思璇何斌源
热带海洋学报 2019年5期
何思璇, 何斌源
防城河口湾鱼类群落结构及其与环境因子关系研究
何思璇1, 何斌源2
1.中央民族大学生命与环境科学学院, 北京 100081;2.广西壮族自治区海洋研究院, 南宁 广西 530022
文章根据2016年5月、8月、11月和2017年2月在防城河口湾海域进行的渔业资源和环境调查数据, 研究了河口湾鱼类群落结构及其空间和季节变化, 分析了鱼类群落结构与主要环境因子的关系。结果表明: 全年调查共渔获鱼类95种, 隶属于2纲12目37科69属, 适温性以暖水性鱼类为主, 生态类型以底层和近底层鱼类为主。其中, 春季60种, 夏季59种, 秋季54种, 冬季46种, 夏—秋季间种类相似性指数最高。全年优势种为褐菖鲉和长鳍篮子鱼, 主要种有黄鳍棘鲷、条马鲾、李氏䲗和真赤鲷等10种; 夏、秋季的最大优势种均为长鳍篮子鱼, 春、冬季的最大优势种分别为褐菖鲉和条马鲾。全湾鱼类年均资源密度为45836尾×km–2和 435.21kg×km–2, 资源密度具有明显的空间差异和季节变化。全年各站位种数变化范围在2~21种, 多样性指数在0.468~4.031, 丰富度指数在0.120~1.926, 均匀度指数在0.181~0.969。聚类分析、单因子相似性分析(ANOSIM)及相似性百分比(SIMPER) 分析表明该海域鱼类群落存在一定程度的时空异质性。相关性分析表明影响防城河口湾鱼类群落结构的主要环境因子为水温、盐度和硝酸盐。
鱼类; 种类组成; 群落结构; 环境因子; 防城河口湾
河口海湾处于陆地与海洋之间的生态交错带, 是地球上生物多样性最丰富、生态服务价值最高的自然生态系统之一, 在保护生物多样性及栖息地、防洪减灾、净化污染、抵御海侵和调节气候等方面具有重要的生态作用, 同时是人类经济发展的重要空间资源(陆健健, 2003)。对于鱼类而言, 河口海湾是其重要的产卵场、索饵场、越冬场和洄游通道, 直接影响其生理活动及种群数量变化(徐兆礼, 2008; 李显森等, 2013; 孙鹏飞等, 2014; 翟璐等, 2015); 同时, 鱼类群落结构与功能可反映海洋生态系统状态, 可作为评价海洋生态系统健康的指标。防城河口湾(108°16′58.8″—108°28′58.8″E, 21°32′31.2″— 21°43′1.2″N)是广西的重要海湾, 位于防城港市江山半岛与企沙半岛之间, 海域面积126.95 km2。通过遥感影像解译发现, 自1990年至今, 围填海使湾口宽度减少了20%, 湾内湿地面积减少了29.7% (即3769.40hm2), 24.4% (即203.84hm2)的原生红树林已消失。流域沿岸向防城河排放大量污水, 加重了防城河口湾水体和沉积物中重金属污染程度(黎清华等, 2014)。在北部湾, 大面积的、较系统的鱼类调查研究多集中在较深海域, 河口海湾的鱼类调查研究则不系统不全面、水平落后, 尤其是防城河口湾, 相关调查频次低、站位少、重复性差, 难以跟踪渔业资源趋势性变化。因此, 本文在该河口湾开展了4个季度的鱼类群落及相关水质指标调查研究海洋渔业资源结构及其与环境因子的相互关系。
1 材料与方法
1.1 调查站位设置与采样
在防城河口湾海域共设置21个调查站位, 对应编号Y1~Y21, 站位分布见图1。分别于2016年5月、8月、11月和2017年2月的当地大潮期, 租用当地渔船开展单船型底拖网采样作业, 底拖网网长10m, 网口宽度3m, 网高1m, 囊网网目尺寸10mm, 曳纲长度30m; 每站拖曳15min左右, 平均拖速5.556km×h–1。所得渔获冷藏带回实验室进行分类、计数和称量。结合各鱼种可捕系数、网具参数、拖网时间及速度计算资源密度(尾×km–2和kg×km–2), 其中, 中上层鱼类可捕系数取0.3, 近底层、底层鱼类取0.5。与鱼类样品采集同步现场测定水深、水色、透明度、水温、pH值、盐度、溶解氧饱和度, 并采集水样带回实验室分析生化需氧量(BOD)、化学需氧量(COD)、亚硝酸盐、硝酸盐、氨氮、无机氮、无机磷、硅酸盐、总氮、总磷、悬浮物、叶绿素等环境因子, 共19项。现场测定、样品采集和保存、室内分析和资料整理均按照《海洋监测规范第4部分:海水分析GB 17378—2007》(中华人民共和国国家质量监督检验检疫总局等, 2008)和《海洋渔业资源调查规范SC/T 9403—2012》(中华人民共和国农业部, 2013)要求进行。
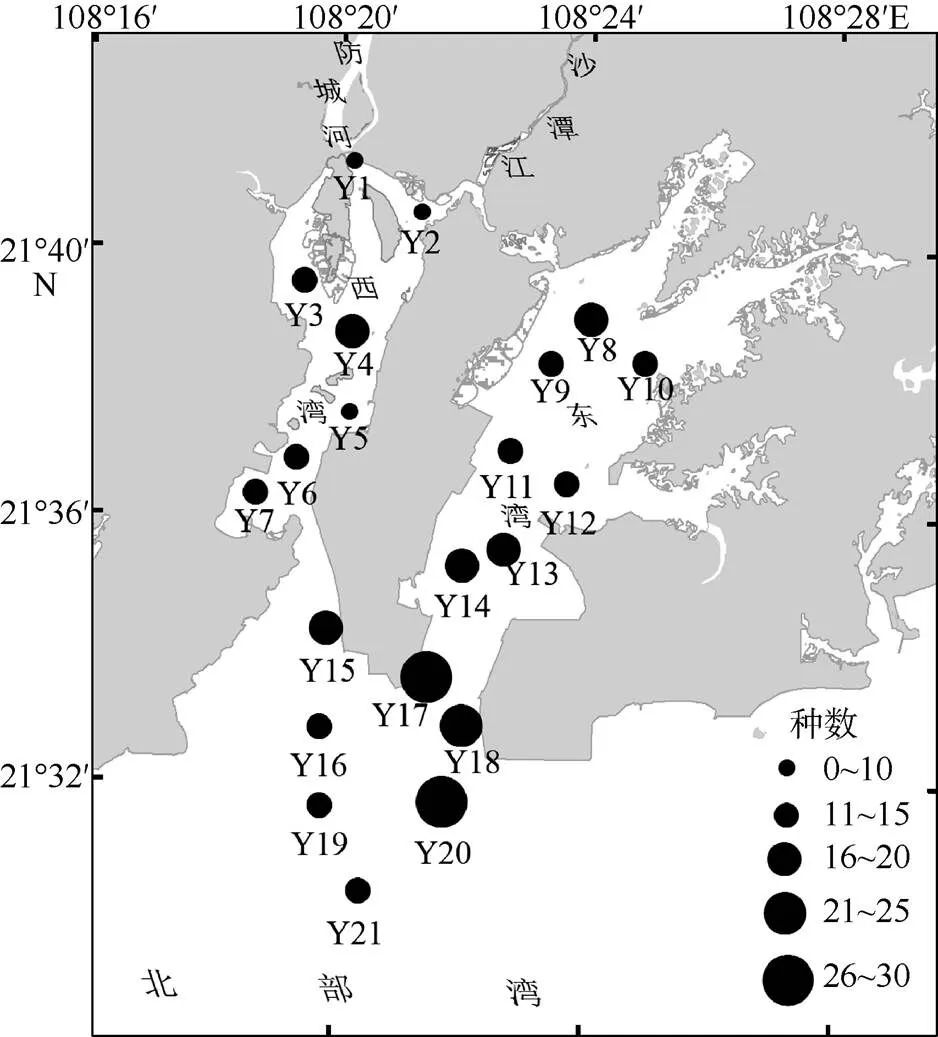
图1 调查站位及鱼类种数空间分布
1.2 数据处理
1.2.1 各种指数计算
Shannon-Weaver多样性指数():

Margalef丰富度指数():

Pielou均匀度指数():

式中:为某站位的所有个体数;为该站位鉴定出的物种数目;p为某站位中第种的个体数占该站位总个数的比例。
物种相似性指数S(单位: %):
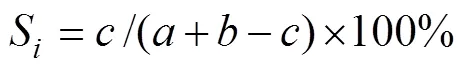
式中:、分别为某两个季节或海区各自的物种数;为两个季节或海区共有的物种数。
相对重要值IRI(单位: %):
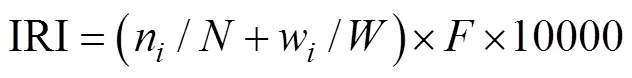
式中:为某季或全年的所有个体数,n为某季或全年第种的个体数;为某季或全年的总生物量,w为某季或全年第种的生物量;为出现频率。按以下分级标准: IRI≥500为优势种, 100≤IRI<500为主要种, 10≤IRI<100为一般种, IRI<10为少见种。
1.2.2 数据分析
采用IBM SPSS Statistics 23.0软件进行鱼类群落的密度、生物量和群落指数的方差分析, 及各群落数量指标与环境因子的相关分析。在PRIMER v6软件中, 把4个季度各站位不同种群的密度数据作4次方根转换, 选择贡献率在5%以上的种类计算Bray-Curtis相似性矩阵, 采用等级聚类分析来研究群落结构, 采用单因子相似性分析(ANOSIM)检验不同组群落结构的差异显著性, 采用相似性百分比 (SIMPER)分析造成各组内群落结构相似的典型种(Clarke, 1993; Clarke et al, 2001)。
2 结果与分析
2.1 群落种类组成
全年共采集到鱼类95种, 隶属于2纲12目37科69属, 软骨鱼纲仅获1种(黄魟), 硬骨鱼纲94种。以鲈形目种数最多, 共60种, 其次为鲽形目和鲉形目, 分别有13种和7种。区系组成以暖水性种为主, 有72种, 占75.8%; 暖温性种23种, 占24.2%, 具有亚热带性质。按栖息水层统计, 所获鱼类以底层鱼类为主, 有56种, 占总种数的58.9%; 近底层鱼类26种, 占27.4%; 中上层鱼类13种, 占13.7%, 总体上群落属近岸生态类型。
春季捕获鱼类10目60种, 夏季8目59种, 秋季8目54种, 冬季7目46种。4个季度都出现的鱼类有21种, 包括鲈形目11种、鲽形目5种、鲉形目4种和鲀形目1种。季节间种类相似性指数大小顺序为: 夏—秋, 56.9; 秋—冬, 51.5; 春—秋, 50.0; 春—夏, 45.1; 夏—冬, 43.8; 春—冬, 39.5。
全年在各站位采集到的鱼类种数差异较大, 在13~37种之间, Y2站位种数最少, Y17种数最多(图1)。总体上可看出一个种数高值区, 包括Y14、Y17、Y18和Y20, 沿着湾口至东湾的深水航道排成一条线; 还有一个亚高值区, 包括站位Y3和Y4, 这里是咸淡水混合区。
将21个站位分为西湾(Y1至Y7)、东湾(Y8至Y14)和湾口(Y15至Y21) 3个分区统计, 可知西湾鱼类有55种, 东湾有60种, 湾口有65种。东湾与西湾共有种39种, 相似性指数51.3; 东湾与湾口共有种42种, 相似性指数50.6; 西湾与湾口共有种40种, 相似性指数50.0。3个分区的共有种有36种, 仅占总种数95种的37.9%。可见区域之间的种类组成有一定程度差异。
2.2 群落优势种
不同季节的鱼类群落优势种结构有所差异(表1)。春季IRI≥100 的种群有8种, 其中IRI≥500 的优势种有4 种: 褐菖鲉、真赤鲷、李氏䲗和二长棘犁齿鲷; 100≤IRI<500 的主要种有4 种: 多鳞鱚、高体斑鲆、黄鳍棘鲷和条马鲾。夏季只有长鳍篮子鱼和黄鳍棘鲷是优势种, 主要种有11 种; 秋季优势种为长鳍篮子鱼、黄鳍棘鲷和皮氏叫姑鱼, 主要种有7种; 冬季优势种有5 种: 条马鲾、日本银鲈、多鳞鱚、黄鳍棘鲷和短吻鲾, 主要种有4 种。周年优势种有褐菖鲉和长鳍篮子鱼, 主要种有黄鳍棘鲷、条马鲾、真赤鲷、李氏䲗和多鳞鱚等10种, 一般种有18种, 少见种有65种。夏季和秋季的最大优势种均为长鳍篮子鱼, 春、冬季的最大优势种分别为褐菖鲉和条马鲾。各个季度全部优势种及主要种的相对重要值之和的占比分别为: 春93.3%、夏86.2%、秋83.1%、冬88.7%; 春季的优势种最集中, 仅褐菖鲉一种的重要值就达6184.83, 占57.4%; 秋季优势种最不明显, 第一优势种的重要值仅为847.94。

表1 防城河口湾鱼类优势种及其相对重要值IRI (单位: %)
注:“/”表示未捕获。
全年30个优势种和主要种之中有11个暖温性种, 包括多鳞鱚、褐菖鲉、黑棘鲷、皮氏叫姑鱼、鹿斑仰口鲾、李氏䲗、及达副叶鲹、铅点多纪鲀、鲬、真赤鲷和中国花鲈。全年23个暖温性种的相对重要值累积占比分别为: 春86.5%、夏53.6%、秋49.4%、冬21.8%。暖温性种的种数虽然远低于暖水性种, 但全年暖温性种的重要值占比达67.2%, 暖水性种占比为32.8%。
2.3 群落密度和生物量
防城河口湾鱼类群落年均密度为45836尾×km–2。表1居前的12个周年优势种和主要种的密度累计占总量的77.8%。各季节均出现高密度区, 春、夏季高密度区集中在湾口, 秋季高密度区在东湾和湾口连接区, 冬季高密度区则在西湾北部区域(图2)。最大密度出现在春季的Y20站位, 达830222尾×km–2;最小密度出现在秋季的Y7站位, 仅1235尾×km–2。鱼类密度有明显的季节变化, 分别为: 春106880尾×km–2、夏34474尾×km–2、秋14390尾×km–2、冬27599尾×km–2; 4个季节之间差异显著(3,80=3.416,<0.05)。3个分区之间的密度差异极显著(2,81= 5.737,<0.01), 大小顺序为: 湾口(98669尾×m–2)>西湾(24692尾×m–2)>东湾(14146尾×m–2)。站位间密度差异不显著(20,63=1.047,>0.05), 主要是由于组内(即季节)密度差异较大, 均方数值大, 导致值较低。

图2 防城河口湾4个季节鱼类群落密度分布
调查海域的鱼类年均生物量为435.21kg×km–2。表1中重要值≥100的12个周年优势种和主要种的生物量累计占总量的64.4%。各季节鱼类的高生物量区分布规律与密度基本相似(图3), 最高生物量出现在春季的Y20站位, 达3527.15kg×km–2; 最小生物量出现在秋季的Y7站位, 仅6.50kg×km–2。不同海域分区之间的生物量差异显著(2,81=3.963,<0.05), 依次为: 湾口(1042.41kg×m–2)>西湾(437.21 kg×m–2)>东湾(401.34kg×m–2)。站位间生物量差异不显著(20,63=1.281,>0.05)。各季节的鱼类生物量大小顺序分别为: 春(587.37kg×km–2)>夏(486.44kg×km–2)>冬(377.43kg×km–2)>秋(289.58 kg×km–2), 季节之间差异不显著(3,80=0.981,>0.05)。
2.4 群落多样性
防城河口湾鱼类群落的站位种数平均为8.8种, 最多达21种, 最少仅2种; 5~7种者较多, 占41.7%; 17种以上者占10.7% (图4)。4个季节的鱼类种数多少依次为: 夏(10.3)>春(10.0)>冬(7.7)>秋(7.0), 季节之间差异显著(3,80=2.971,<0.05)。不同海域分区之间的种数差异显著(2,81=3.133,<0.05), 分别为: 湾口(10.6种)>西湾(8.3种)>东湾(7.4种)。站位间种数差异也显著(20,63=1.898,<0.05)。

图3 防城河口湾4个季节鱼类群落生物量分布

图4 防城河口湾4个季节鱼类群落的种数(a)、种类多样性指数(b)、丰富度指数(c)和均匀度指数(d)
多样性指数范围在0.468~4.031, 平均为2.376, 2~3者占了46.4%, 大于3者占总数21.4%, 有较多区域群落较稳定(图4)。3个海域分区的均值分别为: 西湾(2.473)>东湾(2.357)>湾口(2.296), 差异不显著(2,81=0.440,>0.05)。站位间值差异不显著(20,63=1.100,>0.05)。夏季值高于其他季节, 但季节之间差异不显著(3,80=1.990,>0.05)。
丰富度指数值范围在0.120~1.926, 均值为0.853, 大于1.000者占27.4% (图4)。各季节的值高低依次为: 夏(1.150)>春(1.013)>冬(0.822)>秋(0.756), 季节之间差异显著(3,80=2.895,<0.05)。分区间差异不显著(2,81=1.581,>0.05), 但站位间差异显著(20,63=1.824,<0.05)。
均匀度指数值范围在0.181~0.969之间, 平均为0.821, 0.800以上者占65.5% (图4)。季节间的值差异显著(3,80=3.491,<0.05)。3分区的均值分别为: 西湾(0.868)>东湾(0.859)>湾口(0.736), 差异极显著(2,81=5.615,<0.01)。站位间差异不显著(20,63=1.187,>0.05)。
2.5 群落结构多元分析
图5是根据Bray-Curtis相似性矩阵所做的聚类分析结果。春季群落主要分为4个组, C1组由西湾北部站位组成, C2组由东湾北部站位, C3组主要由湾口区站位组成, C4组由东湾、西湾的中等盐度站位组成, 另有3个站位独立于上述4个组。冬季鱼类群落分为3个组, D1组由湾口深水站位组成, D2组由中等水深的站位组成, D3组由全部西湾站位和东湾北部站位组成, 另有2个站位独立于上述3个组。夏、秋季鱼类群落均分为2个组, 且均为其中一组有2个站位而另一组有19个站位的格局, 夏秋群落结构异质性总体上比春冬两季的低。单因子相似性分析(ANOSIM)分析表明上述4个季度不同组之间群落结构种类组成差异极显著(<0.001)。

图5 4个季节防城河口湾鱼类群落等级聚类图 C1~C4表示春季的4个显著差异的站位组, X1、X2表示夏季的2个显著差异的站位组, Q1、Q2表示秋季的2个显著差异的站位组, D1~D3表示冬季的3个显著差异的站位组。图中虚线表示某个组包括的站位范围
相似性百分比(SIMPER)分析表明: 春季C1~ C4组内平均相似性分别为39.5%、48.2%、69.3%和40.6%。C1组的主要典型种包括中国花鲈、黑棘鲷、日本银鲈、条马鲾、犬牙缰鰕虎鱼和粗高鳍鲉, 对组内相似性的累积贡献率达82.4%。C2组的主要典型种为褐菖鲉、真赤鲷和皮氏叫姑鱼, 贡献率达85.7%。C3组的主要典型种包括褐菖鲉、真赤鲷、李氏䲗、高体斑鲆和多鳞鱚, 贡献率至81.7%。C4组的主要典型种有真赤鲷、粗高鳍鲉、褐菖鲉、东方宽箬鳎和高体斑鲆, 贡献率达87.7%。
夏季2组的组内平均相似性分别为35.8%和30.2%。X1组典型种为黑棘鲷和黄鳍棘鲷, 贡献率达100%; X2组主要典型有鹿斑仰口鲾、斑头舌鳎和黄鳍棘鲷等, 累积贡献率至80.5%。
秋季Q1组的组内平均相似性为20.2%, Q2组的为28.9%。Q1组典型种是鹿斑仰口鲾和红尾银鲈, 贡献率达100%; Q2组主要典型种包括鹿斑仰口鲾、红尾银鲈、卵鳎和长鳍篮子鱼等, 贡献率为90.1%。
冬季D1、D2、D3组的组内平均相似性依次为62.5%、43.2%和42.8%。D1组主要典型种有李氏䲗、高体斑鲆和卵鳎等, 累积贡献率至91.2%; D2组主要典型种有日本钩嘴鳎、斑头舌鳎和大头狗母鱼等, 贡献率达100%; D3组主要典型种有条马鲾、多鳞鱚、日本银鲈等, 贡献率达91.8%。
2.6 群落数量指标与环境因子的相关性
考虑到季节间群落指标数据差异较大而环境因子较相近, 本文将数据分4个季节各自单独地进行相关分析。结果表明群落指标与环境因子的关系具有复杂性和综合性, 19项环境因子与6项群落指标存在1次至9次的显著或极显著相关关系, 但无任何一项环境因子在4个季节都与同一个群落指标存在显著相关关系(表2)。水温和盐度是与群落指标显著相关项次最多的因子, 均为9项次; 硝酸盐有7项次, pH、BOD和总氮均有6项次; 叶绿素仅1项次。与环境因子相关性显著次数较多的群落数量指标有、密度和种数, 分别有21项次、18项次和15项次。密度和生物量对环境因子的反应较为同步, 在春季均与盐度、水温、硝酸盐显著相关, 在夏季均与pH、溶解氧饱和度、BOD、水温显著相关, 在冬季均与水温、盐度、pH值呈负相关关系。秋季的相关性情况不同于其他3个季节,、密度及生物量不与环境因子存在显著相关关系, 全部群落指标仅有10项次显著相关, 其他季节次数分别为: 春季28项次、夏季24项次和冬季23项次。
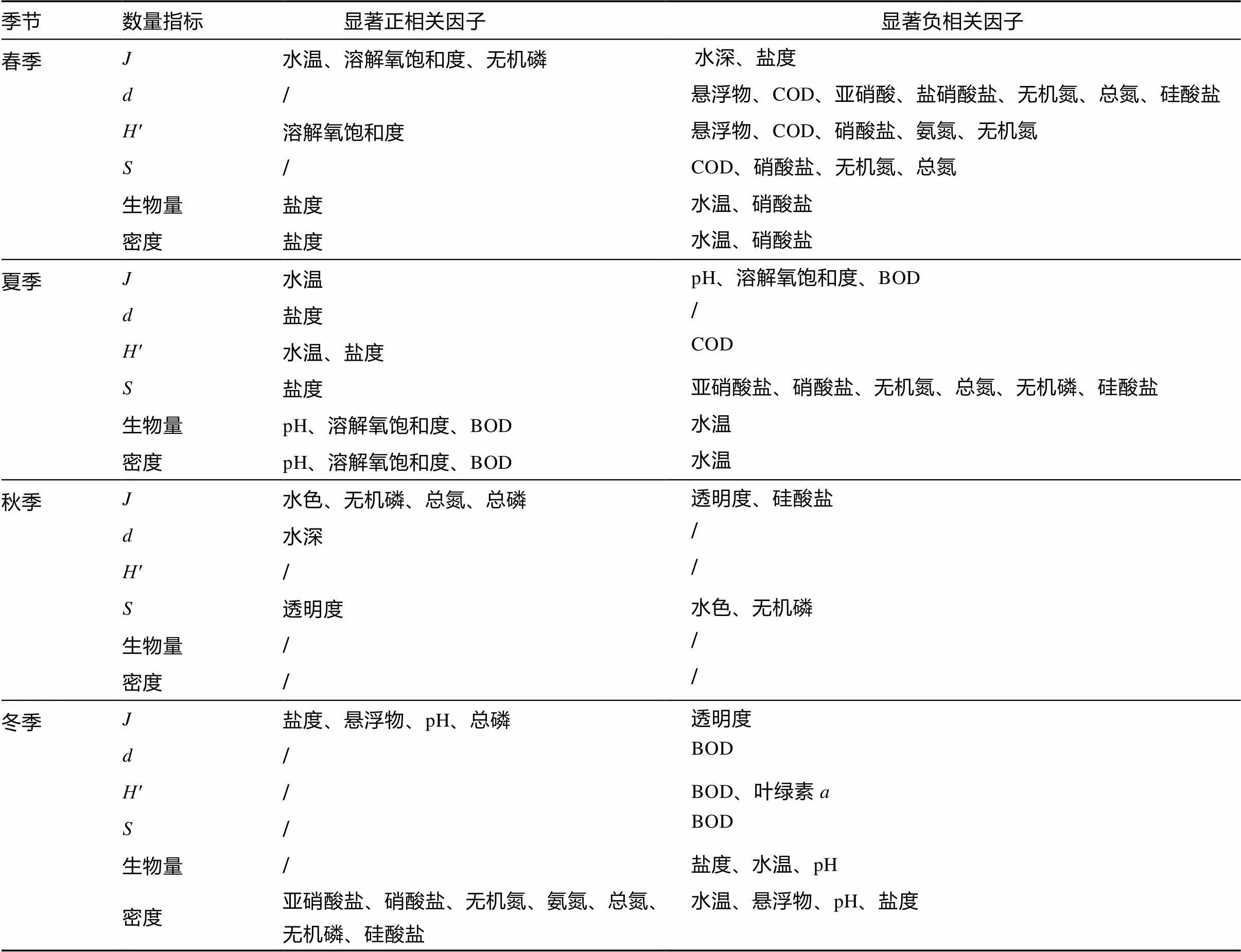
表2 防城河口湾各季节的鱼类群落数量指标与环境因子的相关性
注: “/”指无显著相关的环境因子
3 讨论
3.1 鱼类物种区系性质
北部湾广西海域的鱼类区系具有明显的亚热带性质, 鱼类以暖水性类种为主, 占比81.8%~86.4% (李显森等, 1987; 罗春业等, 1999; 赖廷和等, 2016)。近岸的河口、红树林、海草中的鱼类适温性都呈现相似规律, 暖水性种占绝对优势, 占比71.9%~92.6% (中国海湾志编纂委员会, 1993; 广西海洋开发保护管理委员会, 1996; 何斌源, 1999; 何斌源等, 2001; 何秀玲等, 2003; 王倩等, 2006; 黄德练等, 2013)。本文全年在防城河口湾记录到鱼类95种, 暖水性种占75.8%; 暖温性种占24.2%, 表现出明显的亚热带区系特征, 属于印度—西太平洋鱼类区系的一部分, 这是由该河口湾位于北回归线的边缘南部这一地理区位决定的。但防城河口湾的暖温性鱼类比例高于北部湾及沿岸海湾, 却与福建东山湾(约23°48′N)的研究结果(张静等, 2013)相近, 可能因为防城河口湾地形地貌复杂且红树林繁盛, 为鱼类提供了丰富多样的小生境, 对海洋性暖温种和河口性暖温种均适宜。
3.2 鱼类群落优势种季节及空间分布特征
优势种结构是生物群落特征的重要信息, 优势种主导资源密度的季度分布和空间分布。王雪辉等(2011)指出北部湾10m以深海域(约17°—21°N)鱼类群落以发光鲷、黄斑鲾、日本竹䇲鱼、二长棘犁齿鲷和大头白姑鱼为主要优势种。李渊等(2016)报道春、秋季北部湾的湾口海域(约16°—18°N)鱼类优势种有日本竹荚鱼、发光鲷和红背圆鲹等3种。防城河口湾鱼类周年优势种主要有褐菖鲉、真赤鲷、李氏䲗和二长棘犁齿鲷等, 与上述海区鱼类优势种结构存在不同程度的差异, 这是由其纬度较高、位于近岸和河口等条件决定的。
防城河口湾鱼类资源密度和平均体重的季节变化呈现从春至冬均先升后降, 其趋势由优势种结构主导, 与鱼类生长、洄游规律及海洋捕捞扰动有关。鱼类资源密度在春、夏季均表现为湾口高于内湾(图2), 空间分布格局较为相似, 但群落性质已发生了根本性变化。春季暖温性种占总密度的比例高达78.1%, 到了水温更高的夏秋季则下降至30.0%以下; 春季优势种褐菖鲾和真赤鲷均为暖温性种, 适应春季相对低温、低盐的沿岸水团, 这时洄游至近岸海域产卵繁殖, 渔获绝大多数为幼鱼。冬季褐菖鲉密度不到春季的1%且仅分布在湾口, 甚至未捕到真赤鲷。夏、秋季最大优势种长鳍篮子鱼是暖水性种, 其集中繁殖时期在春末夏初, 夏季平均体重已至11.84g, 夏季密度是其他季节的20倍以上。已有研究认为北部湾鱼类优势种多在北部湾东北部繁育, 夏秋之际洄游至湾西部或深水海域生活(孙典荣, 2008; 傅昕龙等, 2019), 本文结果与之相似。但防城河口湾鱼类资源密度的季节变化与渔业管理状态的耦合程度也较高, 由于南海伏季休渔, 同时春季多为幼鱼不成渔汛, 捕捞活动主要集中在夏末至冬初, 对秋季鱼类资源量影响较大。
3.3 鱼类资源量历史变化
20世纪中后期至21世纪初, 北部湾鱼类资源变化经历了快速下降和缓慢回升两个阶段, 以1999年首次南海伏季休渔为分水岭。1962—1999年渔业资源密度一直呈下降趋势, 1992—1993年资源密度仅为1962 年的43.4%, 1998年资源密度仅为1962 年的16.7%, 同时质量差、寿命短、个体小和营养层次低的种类的替代质量高、寿命长、个体大和营养层次高的种类成为优势种; 自1999年起南海区开始实行伏季休渔, 资源有所恢复; 2000—2001年资源密度上升到1962 年的46.6%, 与1992 年的水平相当, 但难以恢复到1962 年的水平(赖廷和等, 2016)。张文超等(2017)报道2014年夏冬两季海南岛洋浦近岸(约19°44′N)鱼类资源密度为7.30×103尾×km–2和111.17kg×km–2。目前防城河口湾夏冬两季鱼类资源密度平均为31.04×103尾×km–2和 431.94kg×km–2, 对于近岸海域而言属于较高水平。据作者于2010年8月在湾口区设6个站位调查, 湾口区鱼类资源密度为4.65×104尾×km–2和241.20kg×km–2, 平均体重5.19g。本文2016年8月湾口区的鱼类资源密度为7.91×104尾×km–2和944.01g×km–2, 平均体重11.92g, 可知资源密度分别提高了70.2%和291.4%, 平均体重增加了129.9%, 6年间资源得到一定程度的恢复。
3.4 鱼类群落结构与环境因子的关系
影响海洋鱼类群落结构和数量的自然因素复杂而综合, 包括水深、盐度、温度、溶解氧、悬浮物、pH值、地形、水团甚至全球气候变化等自然因素(Colvocoresses et al, 1984; 邱永松, 1988; Greenwood et al, 2007; 王雪辉等, 2010; Aschan et al, 2013; 王小林等, 2013; 王雪辉等, 2013; 单秀娟等, 2014; 管伟等, 2017; 求锦津等, 2018); 同时环境污染、围填海、捕捞等人为因素引起水质底质恶化, 栖息地、产卵场和繁育场被破坏甚至丧失, 导致鱼类群落结构发生明显演替, 呈现小型化和低值化等趋势(胡聪, 2014; 崔磊等, 2017; Lechêne et al, 2018; 叶孙忠等, 2018), 反过来实施湿地修复工程有助于鱼类群落重建(Lechêne et al, 2018)。
影响防城河口湾鱼类群落数量和结构的环境因子较多, 同时水温和盐度是最主要的影响因子。水温与资源密度多呈显著负相关关系, 由于防城河口湾位于北热带边缘, 实测5月水温已到25℃, 8月高至33℃, 鱼类趋向于聚集在相对低温的水域, 如夏季长鳍篮子鱼集中在水温低、水深较大的湾口区Y15站位, 密度达275048尾×km–2, 占该站的90.6%。盐度显著地影响春、冬季鱼类群落结构及数量分布, 但在两个季节所起作用却相反; 由于湾内水域盐度低且水浅, 拖网渔获以出于觅食或洄游需要的外来鱼类为主, 冬季河流流量少, 涨潮时高盐度水团可上溯至更上游的水域, 外来鱼类更多地游到西湾顶部, 冬季优势种条马鲾聚集在西湾顶部的Y1至Y5站位, 密度占比在38.9%~61.6%。春季的优势种褐菖鲉和真赤鲷分布盐度范围很广, 但93.7%的褐菖鲉和92.4%的真赤鲷个体集中在盐度高的湾口区。各种鱼类自身的渗透压调节能力决定了各自的种群分布格局。
秋季鱼类站位平均种数少、数量低、多样性指数不高, 资源密度与19种生境因子均无显著相关关系。虽无法定量但这明显地与渔业活动管控程度同频共振, 由于伏季休渔管控, 捕捞主要集中在夏末至冬初, 导致秋季鱼类群落结构及数量发生较大变化。同时氮磷营养盐、BOD、COD等指标一般起着消极作用。目前防城河口湾正在实施“蓝色海湾”整治项目, 水道疏浚、拆除堤围、生态化海堤建设等工程将急剧改变河口湾水动力、水质和底质状况, 必将影响海洋生物尤其是鱼类群落结构, 应密切跟踪监测并及时评估工程效果, 加强水质管控、生态系统管理与保护, 切实恢复提升资源数量和质量。
崔磊, 吕颂辉, 董悦镭, 等, 2017. 围填海工程对淇澳岛附近水域环境因子与生物群落的影响[J]. 热带海洋学报, 36(2): 96–105. CUI LEI, LÜ SONGHUI, DONG YUELEI, et al, 2017. Influence on the biological community and environmental factors around Qi’ao Island caused by reclamation project[J]. Journal of Tropical Oceanography, 36(2): 96–105 (in Chinese with English abstract).
单秀娟, 陈云龙, 戴芳群, 等, 2014. 黄海中南部不同断面鱼类群落结构及其多样性[J]. 生态学报, 34(2): 377–389. SHAN XIUJUAN, CHEN YUNLONG, DAI FANGQUN, et al, 2014. Variations in fish community structure and diversity in the sections of the central and southern Yellow Sea[J]. Acta Ecologica Sinica, 34(2): 377–389 (in Chinese with English abstract).
傅昕龙, 徐兆礼, 阙江龙, 等, 2019. 北部湾西北部近海鱼类资源的时空分布特征研究[J]. 水产科学, 38(1): 10–18. FU XINLONG, XU ZHAOLI, QUE JIANGLONG, et al, 2019. Temporal-spatial distribution characteristics of fish stocks in north-west coastal waters of Beibu Gulf[J]. Fisheries Science, 38(1): 10–18 (in Chinese with English abstract).
管伟, 徐兆礼, 陈佳杰, 2017. 福建南日岛南部水域鱼类群落结构及多样性[J]. 生态学报, 37(9): 3172–3181. GUAN WEI, XU ZHAOLI, CHEN JIAJIE, 2017. Structure and diversity of fish communities in the waters south of Nanri Island[J]. Acta Ecologica Sinica, 37(9): 3172–3181 (in Chinese with English abstract).
广西海洋开发保护管理委员会, 1996. 广西海岛资源综合调查报告[M]. 南宁: 广西科学技术出版社. Guangxi Oceanic Development and Protection Administration, 1996. Investigation report on the Guangxi sea island resources[M]. Nanning: Guangxi Science and Technology Press (in Chinese).
何斌源, 1999. 广西两港湾红树林鱼类生态的比较研究[J]. 海洋通报, 18(1): 28–35. HE BINYUAN, 1999. Comparative study on the ecology of mangrove fishes between two bays of Guangxi[J]. Marine Science Bulletin, 18(1): 28–35 (in Chinese with English abstract).
何斌源, 范航清, 莫竹承, 2001. 广西英罗港红树林区鱼类多样性研究[J]. 热带海洋学报, 20(4): 74–79. HE BINYUAN, FAN HANGQING, MO ZHUCHENG, 2001. Study on species diversity of fishes in mangrove area of Yingluo Bay, Guangxi province[J]. Journal of Tropical Oceanography, 20(4): 74–79 (in Chinese with English abstract).
何秀玲, 叶宁, 宣立强, 2003. 雷州半岛红树林海区的鱼类种类调查[J]. 湛江海洋大学学报, 23(3): 3–10. HE XIULING, YE NING, XUAN LIQIANG, 2003. Investigation of fishes in mangrove areas of Leizhou Peninsula[J]. Journal of Zhanjiang Ocean University, 23(3): 3–10 (in Chinese with English abstract).
胡聪, 2014. 围填海开发活动对海洋资源影响评价方法研究[D]. 青岛: 中国海洋大学. HU CONG, 2014. Evaluation methods of reclamation impact on marine resources[D]. Qingdao: Ocean University of China (in Chinese with English abstract).
黄德练, 吴志强, 黄亮亮, 等, 2013. 钦州港红树林鱼类群落时间变化格局及其与潮差等环境因子关系[J]. 桂林理工大学学报, 33(3): 454–460. HUANG DELIAN, WU ZHIQIANG, HUANG LIANGLIANG, et al, 2013. Temporal variation of fish community and correlation with environmental variables in Qinzhou harbor mangroves[J]. Journal of Guilin University of Technology, 33(3): 454–460 (in Chinese with English abstract).
赖廷和, 何斌源, 2016. 广西北部湾海洋硬骨鱼类图鉴[M]. 北京: 科学出版社. LAI TINGHE, HE BINYUAN, 2016. Marine osteichthyes fishes in Guangxi Beibu Gulf of China[M]. Beijing: Science Press (in Chinese).
黎清华, 万世明, 何军, 等, 2014. 近两百年来人类活动对北部湾潮间带环境的影响[J]. 海洋地质与第四纪地质, 34(1): 57–64. LI QINGHUA, WAN SHIMING, HE JUN, et al, 2014. Human impact on the intertidal environment in Beibu Gulf over the last 200 years[J]. Marine Geology & Quaternary Geology, 34(1): 57–64 (in Chinese with English abstract).
李显森, 梁志辉, 蒋明星, 1987. 北部湾北部我国沿岸海区鱼类区系的初步调查[J]. 广西科学院学报, 3(2): 95–116. LI XIANSEN, LIANG ZHIHUI, JIANG MINGXING, 1987. A preliminary study on the fish fauna in the northern Beibu Gulf along our coastal area[J]. Journal of Guangxi Academy, 3(2): 95–116 (in Chinese with English abstract).
李显森, 于振海, 孙珊, 等, 2013. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠[J]. 应用生态学报, 24(8): 2353–2359. LI XIANSEN, YU ZHENHAI, SUN SHAN, et al, 2013. Ecological niche breadth and niche overlap of dominant species of fish assemblage in Yangtze River estuary and its adjacent waters[J]. Chinese Journal of Applied Ecology, 24(8): 2353–2359 (in Chinese with English abstract).
李渊, 张静, 张然, 等, 2016. 南沙群岛西南部海域和北部湾口海域底层游泳动物多样性[J]. 中国水产科学, 23(1): 177–187. LI YUAN, ZHANG JING, ZHANG RAN, et al, 2016. Diversity of demersal nekton in the southwestern sea of the Nansha Islands and the mouth of Beibu Bay[J]. Journal of Fishery Sciences of China, 23(1): 177–187 (in Chinese with English abstract).
陆健健, 2003. 河口生态学[M]. 北京: 海洋出版社. LU JIANJIAN, 2003. Estuary ecology[M]. Beijing: Chinese Ocean Press (in Chinese).
罗春业, 李英, 朱瑜, 等, 1999. 广西北部湾鱼类区系的再研究[J]. 广西师范大学学报(自然科学版), 17(2): 85–89. LUO CHUNYE, LI YING, ZHU YU, et al, 1999. A further faunistic study of fishes from the Beibu Bay[J]. Journal of Guangxi Normal University, 17(2): 85–89 (in Chinese with English abstract).
求锦津, 王咏雪, 李铁军, 等, 2018. 舟山长白海域主要游泳动物生态位及其分化研究[J]. 生态学报, 38(18): 6759–6767. QIU JINJIN, WANG YONGXUE, LI TIEJUN, et al, 2018. Study on the niche and differentiation of major nekton species in the Zhoushan Changbai Sea area[J]. Acta Ecologica Sinica, 38(18): 6759–6767 (in Chinese with English abstract).
邱永松, 1988. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 12(4): 303–313. QIU YONGSONG, 1988. The regional changes of fish community on the northern continental shelf of South China Sea[J]. Journal of Fisheries of China, 12(4): 303–313 (in Chinese with English abstract).
孙典荣, 2008. 北部湾渔业资源与渔业可持续发展研究[D]. 青岛: 中国海洋大学. SUN DIANRONG, 2008. A study on fishery resources and sustainable fishery development in the Beibu Bay[D]. Qingdao: Ocean University of China (in Chinese).
孙鹏飞, 单秀娟, 吴强, 等, 2014. 莱州湾及黄河口水域鱼类群落结构的季节变化[J]. 生态学报, 34(2): 367–376. SUN PENGFEI, SHAN XIUJUAN, WU QIANG, et al, 2014. Variations in fish community structure in the Laizhou Bay and the Yellow River Estuary[J]. Acta Ecologica Sinica, 34(2): 367–376 (in Chinese with English abstract).
王倩, 杨光, 吴孝兵, 等, 2006. 广西合浦儒艮国家级自然保护区及邻近水域鱼类种数及保护对策[J]. 应用生态学报, 17(9): 1715–1720. WANG QIAN, YANG GUANG, WU XIAOBING, et al, 2006. Fish resources and their conservation strategies in Hepu Dugong State Nature Reserve and its adjacent waters[J]. Chinese Journal of Applied Ecology, 17(9): 1715–1720 (in Chinese with English abstract).
王小林, 徐宾铎, 纪毓鹏, 等, 2013. 海州湾及邻近海域冬季鱼类群落结构及其与环境因子的关系[J]. 应用生态学报, 24(6): 1707–1714. WANG XIAOLIN, XU BINDUO, JI YUPENG, et al, 2013. Fish community structure and its relationships with environmental factors in Haizhou Bay and adjacent waters of East China in winter[J]. Chinese Journal of Applied Ecology, 24(6): 1707–1714 (in Chinese with English abstract).
王雪辉, 邱永松, 杜飞雁, 等, 2010. 北部湾鱼类群落格局及其与环境因子的关系[J]. 水产学报, 34(10): 1579–1586. WANG XUEHUI, QIU YONGSONG, DU FEIYAN, et al, 2010. Fish community pattern and its relation to environmental factors in the Beibu Gulf[J]. Journal of Fisheries of China, 34(10): 1579–1586 (in Chinese with English abstract).
王雪辉, 邱永松, 杜飞雁, 等, 2011. 北部湾鱼类多样性及优势种的时空变化[J]. 中国水产科学, 18(2): 427–436. WANG XUEHUI, QIU YONGSONG, DU FEIYAN, et al, 2011. Spatio-temporal variability of fish diversity and dominant species in the Beibu Gulf[J]. Journal of Fishery Sciences of China, 18(2): 427–436 (in Chinese with English abstract).
王雪辉, 林昭进, 杜飞雁, 等, 2013. 南海西北部陆架区鱼类的种类组成与群落格局[J]. 生态学报, 33(7): 2225–2235. WANG XUEHUI, LIN ZHAOJIN, DU FEIYAN, et al, 2013. Fish species composition and community pattern in the continental shelf of northwestern South China Sea[J]. Acta Ecologica Sinica, 33(7): 2225–2235 (in Chinese with English abstract).
徐兆礼, 2008. 瓯江口海域夏秋季鱼类多样性[J]. 生态学报, 28(12): 5948–5956. XU ZHAOLI, 2008. Analysis of fish diversity in the waters off the Oujiang estuary in summer and autumn[J]. Acta Ecologica Sinica, 28(12): 5948–5956 (in Chinese with English abstract).
叶孙忠, 罗冬莲, 蔡建堤, 等, 2018. 东山湾渔业生物群落结构及生物量分布特征[J]. 渔业研究, 40(5): 358–365. YE SUNZHONG, LUO DONGLIAN, CAI JIANDI, et al, 2018. Characteristics of community structure and biomass distribution of fishery resources in Dongshan Bay[J]. Journal of Fisheries Research, 40(5): 358–365 (in Chinese with English abstract).
翟璐, 徐宾铎, 纪毓鹏, 等, 2015. 黄河口及其邻近水域夏季鱼类群落空间格局及其与环境因子的关系[J]. 应用生态学报, 26(9): 2852–2858. ZHAI LU, XU BINDUO, JI YUPENG, et al, 2015. Spatial pattern of fish assemblage and the relationship with environmental factors in Yellow River Estuary and its adjacent waters in summer[J]. Chinese Journal of Applied Ecology, 26(9): 2852–2858 (in Chinese with English abstract).
张静, 陈永俊, 宋普庆, 等, 2013. 福建东山湾游泳动物群落物种组成及其多样性[J]. 海洋渔业, 35(1): 15–23. ZHANG JING, CHEN YONGJUN, SONG PUQING, et al, 2013. Nekton composition and diversity of Dongshan Bay, Fujian[J]. Marine Fisheries, 35(1): 15–23 (in Chinese with English abstract).
张文超, 叶振江, 田永军, 等, 2017. 北部湾洋浦海域鱼类群落结构[J]. 生态学杂志, 36(7): 1894–1904. ZHANG WENCHAO, YE ZHENJIANG, TIAN YONGJUN, et al, 2017. Fish community structure in Yangpu waters of Beibu Gulf[J]. Chinese Journal of Ecology, 36(7): 1894–1904 (in Chinese with English abstract).
中国海湾志编纂委员会, 1993. 中国海湾志第十二分册(广西海湾)[M]. 北京: 海洋出版社. China Gulf Chronicle Compilation Committee, 1993. Editorial board of China bay survey (Vol. 12) Guangxi Bay[M]. Beijing: China Ocean Press (in Chinese).
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB 17378.4–2007 海洋监测规范第4部分: 海水分析[S]. 北京: 中国标准出版社. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of China, 2008. GB 17378.4–2007 The specification for marine monitoring— Part 4: seawater analysis[S]. Beijing: Standards Press of China (in Chinese).
中华人民共和国农业部, 2013. SC/T 9403–2012 海洋渔业资源调查规范[S]. 北京: 中国农业出版社. Ministry of Agriculture of the PRC, 2013. SC/T 9403–2012 Technical specification for marine fishery resources survey[S]. Beijing: China Agriculture Press (in Chinese).
ASCHAN M, FOSSHEIM M, GREENACRE M, et al, 2013. Change in fish community structure in the Barents Sea[J]. PLoS One, 8(4): e62748.
CLARKE K R, 1993. Non-parametric multivariate analyses of changes in community structure[J]. Australian Journal of Ecology, 18(1): 117–143.
CLARKE K R, WARWICK R M, 2001. Change in marine communities: an approach to statistical analysis and interpretation[M]. Plymouth: Plymouth Marine Laboratory.
COLVOCORESSES J A, MUSICK J A, 1984. Species associations and community composition of middle Atlantic Bight continental shelf demersal fishes[J]. Fishery Bulletin, 82(2): 295–313.
GREENWOOD M F D, MATHESON R E JR, MCMICHAEL R H JR, et al, 2007. Community structure of shoreline nekton in the estuarine portion of the Alafia River, Florida: differences along a salinity gradient and inflow-related changes[J]. Estuarine, Coastal and Shelf Science, 74(1–2): 223–238.
LECHÊNE A, BOËT P, LAFFAILLE P, et al, 2018. Nekton communities of tidally restored marshes: a whole- estuary approach[J]. Estuarine, Coastal and Shelf Science, 207: 368–382.
Study on fish community structure and its relationship with environmental factors in Fangchenghe Estuary of Guangxi, China
HE Sixuan1, HE Binyuan2
1. College of Life and Environmental Sciences, Minzu University of China, Beijing 100081, China; 2. Guangxi Academy of Oceanography, Nanning 530022, China
Based on the survey in May, August and November 2016 and February 2017, the spatial and seasonal dynamics of the fish community structure and its relationship with the environmental factors in Fangchenghe Estuary of Guangxi, China were studied. A total of 95 species of fishes were collected in four seasons, belonging to two classes, 12 orders, 37 families, and 69 genera. Among them, 72 species were warm water fishes and 23 species were warm temperate fishes, accounting for 75.8% and 24.2% of the total, respectively. As for their ecological types, the bottom and near-bottom fishes were the dominant species, which accounted for 86.3% of the total species. In all, 60 species in spring, 59 species in summer, 54 species in autumn, and 46 species in winter were recorded. The similarity index between summer and autumn was the highest among season pairs. According to the index of relative importance (IRI), the whole-year dominant species wereand, whose IRIs were higher than 500; 10 main species included,,,, and so on, whose IRIs ranged from 100 to 500.acted as the biggest dominant species in both summer and autumn, so didin spring andin winter. The annual average resource density and biomass of the study area were 45836 individual×km–2and 435.21 kg×km–2, respectively. The resource abundance varied significantly among pairs of seasons or regions. The scale of species abundance per sample ranged from 2 to 21 species; the species diversity index, from 0.468 to 4.031; the richness index,from 0.120 to 1.926; and the evenness index, from 0.181 to 0.969. Cluster analysis, analysis of similarity (ANOSIM) and similarity of percentage (SIMPER) showed that there existed some spatial-temporal heterogeneity in the fish community structure. Correlation analysis showed that the water temperature, salinity and nitrate content were the major environmental factors affecting the fish community structure.
fish; species composition; community structure; environmental factor; Fangchenghe Estuary
date: 2019-01-01;
date: 2019-04-28.
National Key Research and Development Program of China (2017YFC0506100); Science and Technology Project of State Oceanic Administration (YLFCJ20164006-F); Science and Technology Project of Guangxi Oceanic Administration (GXHYJ100); Guangxi Science and Technology Project (AA17129002)
HE Binyuan. E-mail: hebinyuan2008@126.com
10.11978/2019001
http://www.jto.ac.cn
2019-01-01;
2019-04-28。
殷波编辑
国家重点研发计划重点专项(2017YFC0506100); 国家海洋局专项(YLFCJ20164006-F); 广西海洋局研究专项(GXHYJ100); 广西科技计划项目(桂科AA17129002)
何思璇(1997—), 女, 广西省东兴市人。主要从事生态学研究。E-mail: hesixuan2018@126.com
何斌源。E-mail: hebinyuan2008@126.com
Editor: YIN Bo
P735.542
A
1009-5470(2019)05-0086-12
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
今日农业(2021年8期)2021-11-28
食品安全导刊(2021年20期)2021-08-30
水上消防(2021年3期)2021-08-21
儿童时代·幸福宝宝(2020年9期)2020-09-08
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
探索科学(2017年5期)2017-06-21
食品工业科技(2014年13期)2014-03-11
