
黄土高原马铃薯不同连作年限土壤理化性质及微生物特性
2019-09-16 02:19:38孙小花胡新元陆立银谢奎忠
干旱地区农业研究 2019年4期
孙小花, 胡新元,陆立银,谢奎忠,张 武
(1.甘肃省农业科学院马铃薯研究所,甘肃 兰州 730070; 2.甘肃省农业科学院,甘肃 兰州 730070)
随着马铃薯产品价格的不断提升和消费逐步向高附加值产品的转变,一些企业及农民合作组织为满足旺盛的订单需求造成马铃薯多年连作,致使产量和效益下降,土壤质量退化,土壤病害连发等问题频发[1-2],许多优质种薯生产基地马铃薯连作现象也极为普遍。因此,缓解和克服由于连作引起的土壤退化和营养失衡等问题已成为马铃薯生产过程中首要需解决的问题。目前,关于连作问题已经在设施蔬菜[3-4]、瓜果[5]、花生[6]、豆类[7-8]和中药材[9-10]等作物上进行了大量研究。马铃薯连作土壤障碍的形成机制及其克服与消除技术国内外已有报道,主要从连作土壤致病菌的产生与病虫害作用机制[11]、植物化感与自毒作用[12]、连作对马铃薯光合作用的影响[13-14]、干物质积累[15]、内源激素代谢[16],以及土壤基因组学[17]等不同角度试图对上述问题进行理解和阐释。而关于马铃薯不同连作年限土壤理化性质及微生物特性的研究报道较为少见,且连作障碍产生的主要原因就是土壤理化性状的改变以及土壤微生物组成的失衡。
因此,本研究以马铃薯不同连作年限的土壤为主要研究对象,依托田间定位试验,对马铃薯不同连作年限土壤容重、土壤结构分维的变化,土壤总有机碳的积累,马铃薯主要生育期土壤酶活性的动态,土壤微生物数量、菌群组成变化等方面进行了探讨,试图从时间尺度上来探讨土壤物理、化学和生物学性质对马铃薯连作的响应,评估可能的连作土壤障碍因子,明确马铃薯连作障碍机理,为解决连作土壤障碍因子的消减技术提供理论依据。
1 材料与方法
1.1 研究区概况
试验布设在甘肃省定西市安定区宁远镇红土村,位于N35°26.714′,E104°50.886′,海拔1 991 m,试验地为平坦川地,属中温带偏旱区,平均海拔2 000 m,日照时数2 476.6 h,年均气温6.4℃,≥10℃积温2 239.1℃,无霜期140 d。多年平均降水量390.9 mm,年蒸发量1 531 mm,干燥度2.53,为典型的黄土高原半干旱雨养农业,属一年一熟区。土壤为典型的黄绵土,不同连作年限土壤基础养分见表1。

表1 不同连作年限土壤基础养分
注:pH值根据水∶土=2.5得到。
Note: the pH value in the
Table was measured with water∶soil ratio of 2.5.
1.2 试验设计
试验地2009年种植小麦,2010年开始进行定位试验,2018年为定位试验第9年。试验设5个处理,处理1:撂荒(CK1),2005年豌豆,2006年起撂荒;处理2:轮作(CK2),小麦/豌豆/马铃薯轮作;处理3:连作2 a马铃薯(2015-2017年连续种3 a马铃薯);处理4:连作4 a马铃薯(2013-2017年连续种5 a马铃薯);处理5:连作6 a马铃薯(2011-2017年连续种7 a马铃薯)。每个处理3次重复,每小区面积23.814 m2(宽4.2 m× 长5.67 m)。马铃薯每个小区种植7行,株距37 cm,行距60 cm。每年3月下旬播种小麦,4月下旬播种马铃薯。小麦每hm2播种量为187.50 kg,施纯N 及P2O5各105.00 kg、农家肥3万kg。马铃薯在4月下旬播种,播种密度为67 500株·hm-2,施肥参照当地标准,每hm2施P2O5108.00 kg、纯N 135.00 kg、农家肥3万kg,其中1/4的氮肥在现蕾期结合培土追施,其余作为基肥施入。播种和施肥均为机械化一次性完成,田间管理同当地大田。
1.3 取样
在马铃薯主要生育期选择地势、肥力等生态因子基本一致的有代表性的样地在每个样方内按五点取样, 根据试验要求分别取(1)播前0~20、20~40 cm土层,测土壤容重;(2)播前、收后0~40 cm土层原状土,测土壤结构分维;(3)播前0~20、 20~40、 40~60、 60~80、 80~100 cm土层土样,自然风干后研磨过0.15 mm筛,测土壤总有机碳(TOC);(4)播前、苗期、盛花期、块茎膨大期、成熟期分别在垄上马铃薯行间靠近植株根系处取样,保鲜冷藏,测土壤酶活性和土壤微生物数量。所有土样均去掉可见的植物根系和残体。
1.4 测定项目与方法
1.4.1 土壤化学因子[18]土壤pH值用电位法;全N用凯氏法;全P用碱熔-钼锑抗比色法;全K用NaOH熔融-火焰光度法;碱解N用扩散法;速效P用Olsen法;速效K用1 mol·L-1NH4Ac浸提-火焰光度法。
1.4.2 土壤容重 采用环刀法[18]。
1.4.3 土壤结构分维 土壤粒径分级采用中国科学院南京土壤研究所提出的我国土粒分级标准[19],即砂粒粒径为0.05~1 mm,粉粒粒径为0.005~0.05 mm,粘粒粒径<0.005 mm。取0~40 cm土层混合样,过>4、4、2、1、0.5、0.25、0.15、0.125、0.055 mm筛。
1.4.4 土壤总有机碳 重铬酸钾氧化法[18]。采用“S”型取样法,在马铃薯播前取0~20、20~40、40~60、60~80、80~100 cm土层混合样。自然风干后研磨过0.15 mm筛。称取通过0.15 mm筛孔的风干土样0.1~1 g,滴加0.800 mol·L-1K2Cr2O7标准溶液5 mL,再加入5 mL浓H2SO4。将试管放于170℃~180℃的油浴锅中加热5 min,后将试管中溶液倒入250 mL三角瓶中,定容至100 mL,加入邻菲罗啉指示剂3滴,用标准的0.2 mol·L-1FeSO4滴定,记下FeSO4滴定体积(V),计算土壤有机碳含量。
1.4.5 土壤酶活性 土壤脲酶采用比色法,过氧化氢酶采用 KMnO4滴定法,(转化酶)蔗糖酶采用3,5-二硝基水杨酸比色法测定[20]。
1.4.6 土壤微生物数量 采用平板计数法[21]。细菌采用牛肉膏蛋白胨培养基, 真菌采用马丁氏培养基, 放线菌采用高氏1号培养基。
1.5 数据统计与分析
应用SPSS 16.0和Excel 2010等软件对试验所得数据进行统计、相关分析、显著性检验,并绘制图表。
2 结果与分析
2.1 马铃薯不同连作年限土壤物理性状
2.1.1 土壤容重 土壤容重是考量土壤综合性质的一个重要参考标准。通过对不同处理土壤0~40cm土层容重的研究表明(见表2):5种不同处理土壤容重在1.19~1.35 g·cm-3范围内,其中0~20 cm土层:土壤容重从小到大依次排序为连作6 a<撂荒(CK1)<连作2 a<轮作(CK2)<连作4 a;而20~40 cm土层土壤容重从小到大依次为连作6 a<轮作(CK2)<连作4 a<连作2 a<撂荒(CK1)。综上所述,0~20 cm土层连作4 a土壤容重最大,为1.33 g·cm-3, 20~40 cm土层撂荒(CK1)处理土壤容重最大,为1.35 g·cm-3;不论哪个层次连作6 a处理土壤容重均最小,仅为1.19~1.22 g·cm-3。连作6 a土壤容重最小,说明土壤疏松多孔,结构性良好;反之,撂荒(CK1)容重大,土壤紧实板硬,缺乏团粒结构,妨碍根系伸展,不利于马铃薯生长发育。
综上分析,对马铃薯播前土壤而言,不同土地利用方式对土壤容重影响较大,撂荒处理土壤板结严重;随着连作年限的增加,人为扰动频繁,土壤容重反而减小。

表2 不同处理土壤容重分析
注:同行数据后标以不同字母者表示在P<0.05水平上差异显著。
Note: Different letters in the same row indicate significant difference atP<0.05 level.
2.1.2 土壤结构分维 任何一种土壤都是由粒径不同的各种土粒组成的,从图1可以看出,各处理>4 mm粒径的土壤最多,占总样的16.14%~32.49%,从小到大依次排序为轮作(CK2)<连作4 a<连作6 a<连作2 a<撂荒(CK1);0.15 mm粒径的土壤占百分比最小,为2.23%~3.61%,从小到大依次排序为撂荒(CK1)<连作2 a<连作6 a<连作4 a<轮作(CK2)。
土壤中各粒级的含量并非平均分配,而是以某一级或两级颗粒的含量和影响为主,从而显示出不同的颗粒性质。由图2可以看出,同一种处理下土壤各粒径含量差异很大。撂荒(CK1)处理>4 mm~0.055 mm粒径的土壤占15.8%~32.49%、轮作(CK2)处理>4 mm~0.055 mm粒径的土壤占3.61%~19.88%、连作2 a>4 mm~0.055 mm粒径的土壤占2.37%~31.40%、连作4 a>4 mm~0.055 mm粒径的土壤占3.41%~20.99%、连作6 a>4 mm~0.055 mm粒径的土壤占3.03%~21.88%。
2.2 马铃薯不同连作年限土壤化学性质
2.2.1 土壤总有机碳 耕层土壤总有机碳含量变化直接影响作物的生长状况,是土壤生产潜力和土壤质量的一个重要体现。由图3可知,马铃薯连作农田土壤总有机碳的动态变化趋势呈明显下降趋势,撂荒处理土壤总有机碳含量最大,为9.53 g·kg-1,连作6 a土壤总有机碳含量最低,为8.17 g·kg-1,连作6 a土壤总有机碳含量比撂荒降低了14.27%,可见连作年限对土壤总有机碳的影响很大,撂荒更有利于土壤TOC(总有机碳)的积累。
为了研究不同处理土壤垂直剖面中总有机碳的变化特征,本研究在马铃薯播前分别取0~20、20~40、40~60、60~80、80~100 cm土层混合样,结果如图4、图5所示, 由于前茬作物残体的凋落、根茬的残留及土壤微生物的分解,使得农田土壤碳库表现出动态变化。各处理土壤总有机碳积累量为5.79~8.52 g·kg-1;0~20 cm土壤TOC积累量显著高于其它处理,说明撂荒有利于土壤TOC的积累。
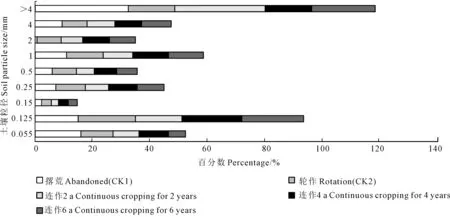
图1 不同粒径土壤所占百分数Fig.1 Percentage of soil with different particle sizes

图2 同一处理不同粒径土壤所占比例Fig.2 Proportion of soil with different size in the same treatment

图3 播前不同处理耕层土壤总有机碳含量Fig.3 Total soil organic carbon contents before sowing
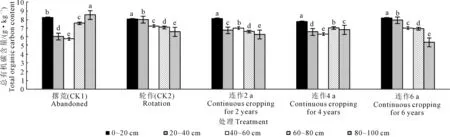
注:不同字母表示在P<0.05水平上差异显著,下同。 Note: Different letters are significant differences at P<0.05 level, the same below.图4 马铃薯苗期不同处理各层次土壤总有机碳含量Fig.4 Total soil organic carbon content of potato at seedling stage with different treatments
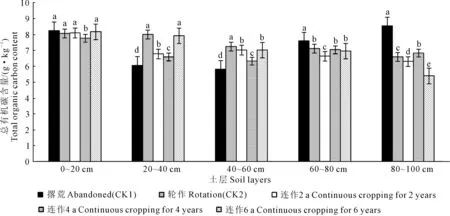
图5 马铃薯苗期不同土层各处理土壤总有机碳含量Fig.5 Total soil organic carbon content in different soil layers at seedling stage
2.2.2 土壤酶活性
转化酶:马铃薯主要生育期内土壤转化酶呈“S”型动态变化趋势,出现先高后低又升高的变化过程(图6)。峰值出现在苗期,为23.24~29.50 mg·g-1·d-1;块茎膨大期最低,为23.48~26.45 mg·g-1·d-1。酶活性的提高可加快有机质的分解,从而提高土壤养分含量。随着马铃薯生育期的推进,酶活性较理化性质的变化更加敏感,对土壤肥力的演化具有重要影响。
脲酶:马铃薯主要生育期内土壤脲酶的动态变化呈“M”型趋势,出现双峰变化过程(图7)。第一个峰值出现在苗期,为1.55~1.91 mg·g-1·d-1;第二个峰值块茎膨大期最低,为1.31~1.79 mg·g-1·d-1。
过氧化氢酶:马铃薯主要生育期内土壤过氧化氢酶的动态变化呈“S”型趋势,出现先高后低的变化过程(图8)。峰值出现在播前,为3.87~4.17 mL·g-1·d-1,苗期最低,为3.57~4.22 mL·g-1·d-1。
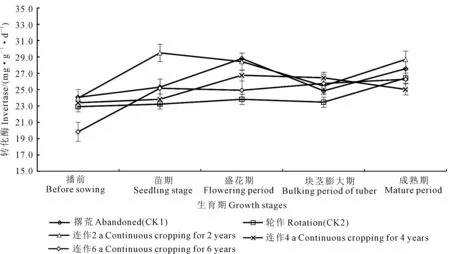
图6 马铃薯主要生育期土壤转化酶动态变化Fig.6 Dynamic changes of soil invertase in potato field at main growth stages
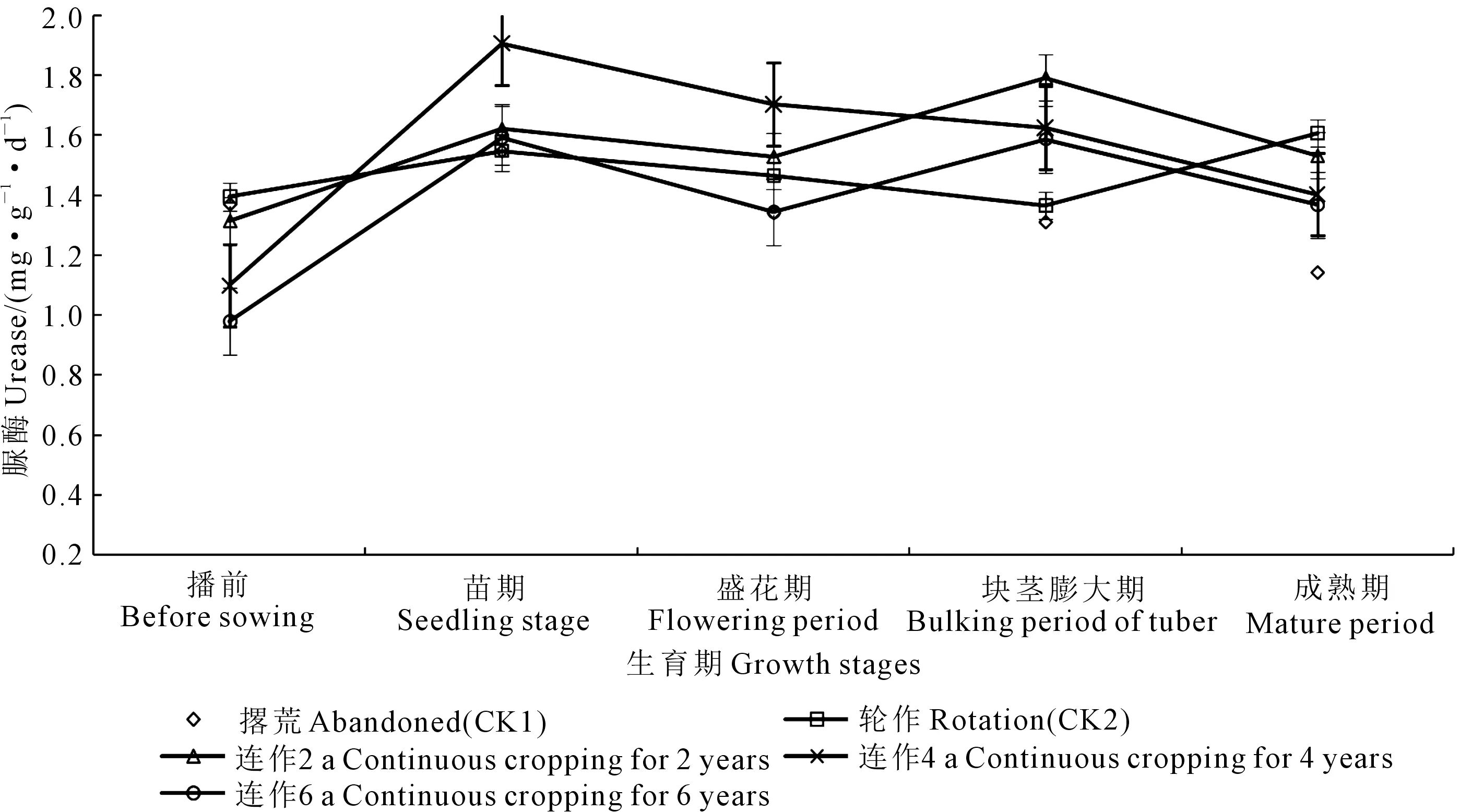
图7 马铃薯主要生育期土壤脲酶动态变化Fig.7 Dynamic changes of soil urease in the main growth period of potato

图8 马铃薯主要生育期土壤过氧化氢酶动态变化Fig.8 Dynamic changes of soil catalase in potato field at main growth stages
在土壤养分(氮、磷、钾、有机质等)和土壤酶的共同作用下,土壤中进行着复杂的生化反应和物质循环。由表3可知,转化酶、脲酶、过氧化氢酶3种土壤酶之间均呈极显著正相关;3种酶与容重呈极显著负相关,与其他肥力因子均呈极显著正相关,表明3种酶在促进土壤养分转化,改良土壤理化性状方面发挥着重要作用。同一种酶不仅对特定的土壤肥力因子有显著相关性,而且对多种土壤养分因子均有极显著的相关性,说明3种土壤酶不仅在促进单一土壤养分因子转化中发挥着作用,而且均参与了其他土壤养分因子的转化过程,共同影响着土壤的理化性状。
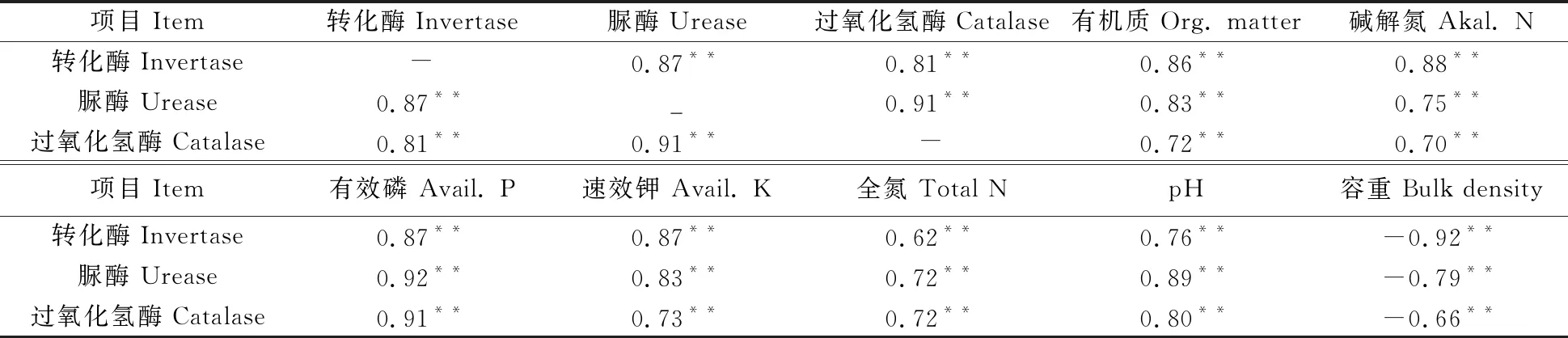
表3 土壤酶活性与主要养分含量的相关性分析
注:*、**分别表示在5%和1%水平相关性显著。
Note:*, ** mean significant correlation at 5% and 1% levels respectively.
为进一步探讨土壤酶活性与土壤肥力的关系,对不同连作年限的土壤酶活性与土壤肥力因子进行主成分分析,以筛选出产生影响的主要因子群。由表4可知,前两个主成分的累计方差贡献率为89.29%(>85%) ,能反映土壤肥力系统的变异信息。通过对供试土壤主成分的规格化特征向量分析,第一主成分土壤酶(转化酶、脲酶、过氧化氢酶)与土壤主要养分(有机质、碱解氮、有效磷、速效钾、全氮、pH)在全部因子中占主导地位,是土壤肥力的重要组成部分,方差贡献率达到81.15%,在全部因子中占主导地位,是土壤肥力的最重要方面,故第一主成分代表了土壤各项指标的大小,是反映“土壤肥力水平”的综合指标。因此,第一主成分可以近似地表示土壤的综合肥力。
2.2.3 土壤微生物数量 土壤微生物是土壤养分的活性部分,用于评价土壤质量的生物学性状。土壤微生物群落(主要有细菌、放线菌和真菌三大类)主要参与土壤生物化学过程、有机物的分解转化。本研究对不同连作年限的土壤分为两个层次,即0~10、10~20 cm。由表5可知,细菌:0~10 cm土层连作2 a的细菌数量显著高于撂荒、连作4 a和连作6 a;真菌:0~10 cm土层轮作显著高于撂荒、连作2 a、连作4 a和连作6 a;放线菌:0~10 cm土层撂荒处理显著高于轮作、连作2 a、连作4 a和连作6 a。微生物数量随着土壤深度的增加而减少。
各处理中的土壤中微生物数量,细菌>放线菌>真菌,且细菌数量占95%以上,占绝对优势,说明土
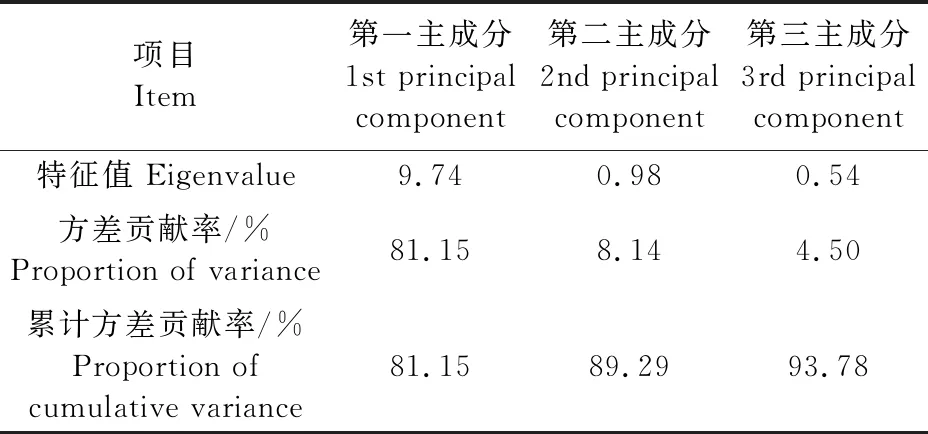
表4 供试土壤主成分特征值
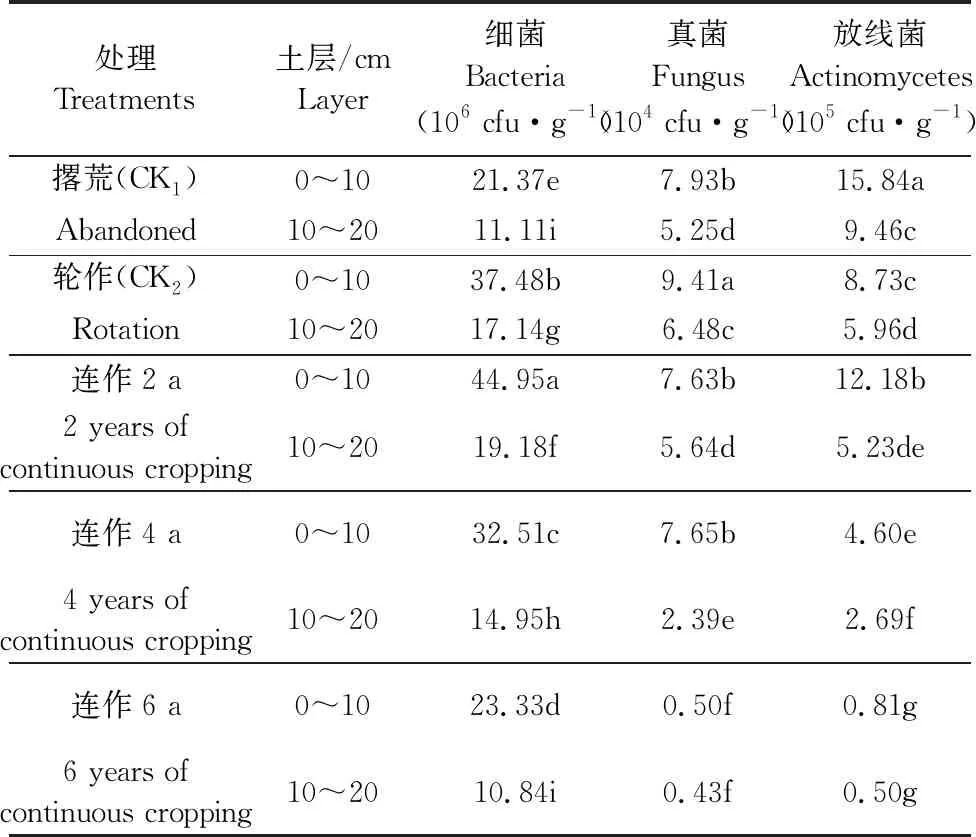
表5 不同处理土壤微生物数量方差分析
注:同列数据后标以不同字母者表示在P<0.05水平上差异显著。
Note: Different letters in the same column indicate that the difference is significant atP<0.05 level.
壤中细菌是优势菌,真菌和放线菌也占有一定比重。在0~10 cm土层中,细菌数量的大小顺序为:连作2 a>轮作>连作4 a>连作6 a>撂荒;真菌数量以轮作0~10 cm最高,连作6 a 10~20 cm最低;放线菌数量为撂荒>连作2 a>轮作>连作4 a>连作6 a。由于微生物的数量直接影响土壤有机质的含量,因此预示着微生物群落结构也随之改变,进而影响作物的生长状况。
3 讨 论
1)长期连作通过改变土壤中不同粒级团聚体比例而降低有机碳的稳定性[22]。本研究以粒径重量分布取代粒径数量分布来描述土壤分形特征,探究黄土高原旱作区马铃薯连作区土壤团粒结构分维特征。结果表明,连作6 a土壤容重最小,且连作可以改变土壤中不同粒级团聚体的比例,各处理中>4 mm粒径的土壤含量最高。徐雪风[23]等对盆栽马铃薯不同连作年限土壤生物和非生物环境因子的研究发现:土壤容重随着连作年限的增加而逐年上升,在连作7 a时比对照高5.39%。本研究所得结论有所不同,究其原因,本研究是在田间自然条件下进行的,土壤容重受人为扰动频率不大,只有在耕种、采样时有翻动,而盆栽试验土壤人为翻动更多,这也为马铃薯连作土壤物理性状的研究提供了新思路、新方法。
2)有机碳是维持土壤可持续生产力的关键因素,且有机碳库与土壤质量和作物生产力有密切关系。本研究表明,马铃薯长期连作显著降低土壤有机碳含量,连作6 a土壤总有机碳含量比撂荒降低了14.27%,即随着连作年限延长,土壤有机碳含量呈逐渐降低趋势,相似结果也被早前学者报道[24]。然而,黄金花等[25]对秸秆还田下长期连作棉田土壤有机碳活性组分的变化特征研究表明,棉花长期连作导致 0~40 cm 深度内土壤有机碳的活性指数显著增加,但降解指数则显著降低,与本研究结果相反。本研究定位试验虽有农家肥的投入,但对土壤总有机碳的影响不大,可能是由于农家肥对土壤活性有机碳(易氧化有机碳、颗粒有机碳等)的变化影响较大。农家肥的投入对土壤活性有机碳的动态影响还需要进一步研究。
3)土壤酶参与土壤有机质的分解、合成以及氮、磷、钾等物质的循环,其活性强度随土壤理化性质、有机物含量等因素的变化而变化[26-27]。本研究中,马铃薯主要生育期内土壤转化酶、过氧化氢酶变化动态呈“S”型曲线,脲酶呈“M”型变化趋势;3种酶活性与土壤肥力因子均呈极显著正相关,与土壤容重呈极显著负相关。农作物由于其自身营养特性的原因对于土壤养分并非均衡性吸收,有学者据此认为长此以往所带来的土壤养分亏缺、比例失调或不平衡会导致连作障碍发生[28],土壤酶在促进土壤中碳源、有机物质等的转化中并不是孤立的,而是紧密联系且互相影响的。这些研究结论的不一致体现了土壤酶活性、土壤养分含量与微生物之间关系的复杂性。
4)本研究表明,马铃薯长期连作显著降低土壤微生物数量,土壤微生物群落结构随连作年限明显改变。秦越[29]等对不同连作年限马铃薯播种前和成熟期根际土壤微生物多样性的研究表明,连作导致马铃薯根际土壤细菌多样性水平降低,这与吴凤芝等[3]对黄瓜连作、李春格等[30]对大豆连作的研究结果相一致。马铃薯连作显著影响土壤微生物群落结构,相似的结果也在其他作物上得到证明[31-32]。而导致连作土壤微生物群落结构出现这种定向性变化的深层次机理尚不明确,可能与根系分泌物有关。
4 结 论
本研究不同处理土壤容重差异显著,连作6 a土壤容重最小,连作改变了土壤中不同粒级的分配比例,>4 mm粒径的土壤含量最高。马铃薯长期连作显著降低土壤总有机碳含量,连作6 a土壤总有机碳含量较撂荒降低14.27%。马铃薯主要生育期内土壤转化酶、过氧化氢酶、脲酶活性与土壤肥力因子均呈极显著正相关,与土壤容重呈极显著负相关。马铃薯长期连作显著降低土壤微生物数量,群落结构随连作年限明显改变。因此,连作马铃薯农田土壤理化性质和土壤微生物群落结构之间的关系将是未来解决连作土壤障碍因子消减技术的关键点。
猜你喜欢
建材发展导向(2022年10期)2022-07-28 03:04:22
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
现代畜牧科技(2021年4期)2021-07-21 06:13:08
当代陕西(2020年24期)2020-02-01 07:06:40
当代陕西(2020年24期)2020-02-01 07:06:36
当代陕西(2020年24期)2020-02-01 07:06:36
中成药(2017年6期)2017-06-13 07:30:35
陕西画报(2016年1期)2016-12-01 05:35:30
创新作文(小学版)(2016年31期)2016-03-11 19:08:09
动物营养学报(2015年3期)2016-01-07 11:06:10
