
西双版纳地区鞘花的繁殖物候特征及影响因素
2019-09-10 07:22:44李慢如张玲
广西植物 2019年9期
李慢如 张玲
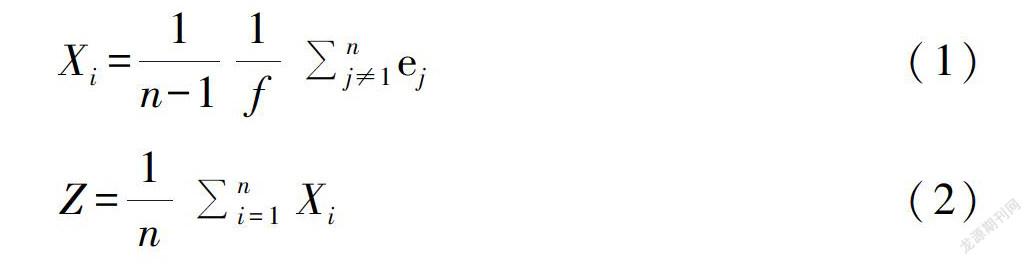


摘要: 桑寄生植物作為关键种和关键性食物资源在生态系统中扮演着重要角色,其鞘花的繁殖物候特征不仅会影响自身的繁殖适合度而且还会影响依赖于其获取食物资源的动物。为了解鞘花的繁殖物候特征及其影响因素,探究其与寄主植物和种子散布者之间的相互作用关系。该研究以西双版纳地区分布的鞘花为对象,通过定期观测鞘花和其寄主植物木荷的繁殖物候,测量它们的生物学特性和温湿度等环境因子,并分析鞘花在个体水平和种群水平上的繁殖物候特征以及寄主植物和温湿度对其繁殖物候的影响。结果表明:(1)鞘花的开花物候属于集中大量开花模式,整个种群的花期和果期的持续时间分别约为20 d和72 d,花期和果期的同步性指数都较高,6月中旬鞘花果实被取食的数量和速率最大,之后逐渐降低。(2)鞘花的始花期与木荷的始花期相关性较高,花期和果期与木荷的繁殖物候基本重叠。(3)每月开花和果熟的个体数量与同期和前一个月的平均温度和相对湿度的相关关系均不显著。总之,桑寄生植物的繁殖物候特征可能受到较多因素的影响,若想全面了解半寄生植物的繁殖物候特征,就要综合考虑生物和非生物等多种因子的共同作用。
关键词: 开花物候, 同步性, 种子散布, 寄主植物, 桑寄生
中图分类号: Q948.9文献标识码: A文章编号: 1000-3142(2019)09-1252-09
Abstract: Mistletoes play an important role in the ecosystem as key stone species and critical food resources for many birds and mammals. As a main kind of mistletoe species in Xishuangbanna region, the reproductive pheno-logical characteristics of Macrosolen cochinchinensis can not only affect their reproductive fitness, but also affect the animals that depend on them for food resources. To understand the reproductive phenological characteristics and impact factors of this mistletoe species, and also to explore the interaction between host plants and its seed dispersers of this mistletoe, we studied the reproductive phenology of M. cochinchinensis in Xishuangbanna region. We observed the reproductive pheno-logy of M. cochinchinensis and its host species Schima superba at regular intervals, and measured their biological traits and environmental factors like temperature and humidity. Meanwhile, we analyzed the reproductive phenological characteristics of Macrosolen cochinchinensis at individual and population levels, and probe into the effects of its host plants and environmental factors (temperature and humidity etc.) on their reproductive phenology at the same time. The main results were as follows: (1) The flowering phenology of M. cochinchinensis belong to mass flowering patterns. The flowering and fruit duration of the whole population were about 20 d and 72 d respectively. The synchrony index of flowering and fruiting duration was relatively higher. The number of fruits foraged and rate of fruit foraged reached the maximum peak in the middle of June, and then gradua decreased. (2) The first flowering date of M. cochinchinensis and its host species Schima superba was highly correlated, and the flowering and fruiting period of Macrosolen cochinchinensis and Schima superba were mainly overlapped in time scale. (3) Individuals numbers of flowering and fruit mature plants per month of Macrosolen cochinchinensis was not significantly correlated with the average temperature and relative humidity of the same and previous months. In summary, the reproductive phenological characteristics of mistletoe species may be influenced by many factors, and it is essential to consider comprehensively combination of multiple factors such as many biotic and abiotic factors to understand the reproductive phenological characteristics of those hemiparasite mistletoes in the ecosystem.
Key words: flowering phenology, synchrony, seed dispersal, host plants, mistletoe
植物具有季节性和周期性的繁殖物候事件是其生活史的重要部分(Rathcke & Lacey, 1985; Steltzer & Post, 2009),不仅可以影响自身的传粉和种子散布等,而且还会影响依赖这些植物获取资源的动物(Wright & Meagher, 2003)。植物的繁殖物候主要分为开花物候和结果物候。开花物候通常较多地关注植物具有繁殖能力即盛开(花药散发)阶段;结果物候包括果实出现、生长、成熟和种子散布(Hilty, 1980; Rathcke & Lacey, 1985; Buide et al., 2002; 刘志民和蒋德明, 2007)。同时还可以用如开始(onset)、时长 (duration)和同步性(synchrony)等多个物候参数的组合来进行定量描述(Augspurger, 1983; Rathcke & Lacey, 1985; Clark & Thompson, 2011)。植物的繁殖时间除了受系统发育因子的影响外 (Wright & Calderon, 1995; Davies et al.,2013),还会受到多种生物因子(如同域分布的其它植物、传粉者和种子散布者)和非生物因子(如温度、光周期和降水等)的影响(Wright & Meagher, 2003; Elzinga et al., 2007; Craine et al., 2012)。此外,植物自身的生物学特性也能影响植物的开花物候和繁殖适合度,如个体较大的植物比个体较小的植物更早开花且花期更长(Ollerton & Lack, 1998),同步性较低(Otarola et al., 2013)。
桑寄生植物(mistletoes)是一类自身可以进行光合作用的半寄生性常绿灌木的总称,在生态系统中由于其特殊的生活型和生活史过程,与寄主植物、传粉者和种子散布者广泛发生作用(Watson, 2001; Aukema, 2003)。它们通过专性固着器官即吸器(haustorium)附着在寄主植物的枝条,从其寄主植物的木质部吸收水分和养分以满足自身生活史发展的需求(Mathiasen et al., 2008)。有研究表明当寄主植物与寄生植物的繁殖物候存在部分重叠时,与未被感染的同种植物相比,被感染的寄主植物将吸引更多的传粉者和种子散布者(Candia et al., 2014)。同时,种子散布对于桑寄生植物的生存至关重要(Aizen, 2003),大多数桑寄生植物的种子都依赖于鸟类进行散布(Room, 1972),鸟类取食之后通过排泄和回吐的方式将黏性极强的种子蹭到寄主植物的枝条上,适应性较强的种子就开始萌发和定殖(Davidar, 1983a)。在桑寄生植物的繁殖物候阶段,林内寄生感染区的鸟类丰度和多度也会明显提升(Bennetts et al., 1996; Watson, 2001; Napier et al., 2014),因此它们之间的相互作用关系也被认为是一种弥散型的协同进化(Watson, 2001; Aukema & Del Rio, 2002)。
到目前为止,在城市和半荒漠草地灌丛生态系统中均有桑寄生繁殖物候特征及影响因素的相关研究(Teixeira-Costa et al., 2017; Yule & Bronstein, 2018)。但是,在热带雨林生态系统中这类研究还未见开展,其繁殖物候是否同样受寄主植物的物候与种类、其自身的生物学特性以及气候因子的影响还并不清楚。因此,本研究以西双版纳地区分布的一种桑寄生科(Loranthaceae)植物——鞘花(Macrosolen cochinchinensis)为对象,具体回答以下两个科学问题:(1)鞘花具有怎样的繁殖物候特征;(2)鞘花的繁殖物候特征是否受到寄主植物的物候和生物学特性、自身的生物学特性和温湿度等生物因子和非生物因子的影响。希望通过本项研究加深对寄主-桑寄生-种子散布者系统中桑寄生-寄主偏害共生关系和桑寄生-鸟类互利共生关系的理解,同时在全球气候变化背景下为森林管理中合理防治半寄生植物提供基础资料和理论依据。
1材料與方法
1.1 物候观测
该研究在云南省西双版纳国家级自然保护区-勐仑片区(101°07′—101°09′ E,21°58′—21°59′ N)的热带雨林里进行。该地区属热带北缘,气候类型为热带季风气候,年平均气温约22 ℃,年平均降水量1 500 mm(朱华等, 2015)。根据降水量的多少又可将一年分为三季:雨季(5月中旬前后至10月下旬)、雾凉季(10月下旬至次年3月中下旬)、旱季(3月中下旬至5月中旬前后)(张克映, 1963)。选取感染在8棵寄主植物木荷(Schima superba)上生长良好且具有繁殖能力的鞘花24株,该种植物主要分布于中国西南和东南地区(汪强等, 1996),多见于原生林、次生林的高大乔木上和人为干扰程度较大的生境中,种子散布者多为啄花鸟科(Dicaeidae)等体型较小的鸟类(肖来云和普正和, 1994),同时使用双筒望远镜辅助物候观测。
从2017年开始对位于中国科学院西双版纳热带植物园内的桑寄生植物物候观测得知,鞘花的花果期大都集中在每年的3月至8月份,为了更完整地观测记录鞘花的繁殖物候,本研究的物候观测时间为2018年2月至同年9月,约每周观测一次,记录鞘花和木荷的开花物候(始花期和末花期)和结果物候(果熟初期和果末期)共4个物候参数,其中果熟初期仅针对鞘花,木荷的结果物候则从果实出现开始记录,并将以上物候事件出现的日期转换成儒略日(Julian day)即物候事件出现的日期为距离该年1月1日的天数 (陈发军等, 2017)以方便计算。物候观测标准定义为单个植株上开始有第一朵花出现时记为个体的始花期,由于每次观测时间的间隔不完全均等,若在当次观测中发现植物已经开花,则再结合花色判断该个体植株的始花期;最后一次记录到植株枝条上有花朵存在记为末花期。为进一步了解鸟类取食鞘花果实的时间动态,用孔径为5 mm的纱网和四根长为1.5 m的PCV管制作面积为1.5 m × 1.5 m的果皮收集框并用铁架支撑,放置在4株鞘花的正下方,这些个体与其它鞘花个体在空间上没有重叠以保证每一个果皮收集框的果皮均来自于相同的个体,每周收集一次并统计被鸟类取食的果皮数量。取食速率为每次收集到的果皮数量除以两次收集时间间隔的天数,结果用平均值±标准误表示。
1.2 物候参数的计算
始花期和果熟初期按照上述物候观测的标准记录;花期长短与果期长短则为始花/果熟初期至末花期/果末期的时间间隔;个体植物和种群的异步性程度则使用Augspurger (1983)提出的同步性指数计算公式进行衡量。
式中,Xi用于表示种群中任意一个给定的个体与除它之外的其它所有同种个体的物候同步性程度(公式1),Z用于表示种群中任意一个给定的个体与其它所有同种个体的物候同步性指数求和的平均值(公式2),Xi与Z的变异范围均为0~1,“0”表示种群内个体之间的花期无重叠,“1”则表示完全重叠。
1.3 生物学特性及温湿度的测定
生物学特性测定指标包括木荷的胸径、冠幅以及鞘花的寄生高度和冠幅等。使用围尺在1.3 m处测量木荷的胸径,使用测距仪测量木荷的树高和鞘花的寄生高度,并估测木荷和鞘花的冠幅。使用ibuttonDS1923温湿度记录仪(上海沃第森电子科技有限公司),每90 min采集一次样地内的温湿度数据。
1.4 数据分析
使用Shapiro-wilk方法对该文各变量进行正态性检验,由于有些变量经过转化后仍不正态,因此鞘花繁殖物候的4个物候参数之间,以及它们与自身的生物学特性(寄生冠幅、寄生高度)、寄主生物学特性(寄主冠幅、寄主高度)和温湿度统一使用Spearman相关分析法计算相关系数并检验相关性的显著性;当寄生/寄主的生物学特性与物候参数之间的相关性,每月鞘花开花/果熟个体数与当月/前一个月的温湿度之间的相关性达到显著后,再使用回归分析方法进行检验。数据分析和作图使用R3.3.1、SigmaPlot12.5和Adobe Illustrator CS6软件完成。
2结果与分析
2.1 鞘花的繁殖物候特征
2.1.1 开花物候研究区的鞘花约在3月末、4月初开花,4月上中旬达到开花高峰期,整个种群在4月下旬进入末花期,花期持续时间最短为10 d,最长为31 d,平均花期约20 d,属于单峰式集中大量开花模式,且开花物候曲线呈现右倾式。较早开花的个体相应地花期也越长(r=-0.864,P<0.001)。整个种群平均始花期错开约6 d,其中最大的可相差20 d(图1),种群花期的同步性指数为0.79。
2.1.2 果期物候4月中下旬鞘花上开始出现果实,5月中下旬果实开始由绿色变成橙色,之后逐渐成熟。整个种群在8月初进入果末期,果期持续时间最短为63 d,最长为97 d,平均果期约为72 d,果期物候模式属于平顶式,即所有个体均能在相当长的一段时间内有熟果的存在。与开花物候特征类似,果实越早成熟的个体相应地果期也越长(r=-0.777,P<0.001),果实越晚成熟的个体,同步性指数越大(r=0.815,P<0.001)。整个种群平均果熟初期的错开时间约7 d,其中最大的可相差27 d(图1),与花期相比,果期的同步性更高(Z=0.88),但是个体水平上花期与果期的同步性指數相关性不显著(r=0.115,P=0.699)。
另外,随着果实的发育成熟,5月末收集到第一批被鸟类取食后丢弃的果实外果皮,果皮取食的数量在6月中旬达到最高(96.3±18.6),随后呈下降趋势;果实的取食速率与果实的取食数量具有相同的趋势,整个种群于7月中下旬进入果末期(图2)。
2.2 寄主物候对鞘花繁殖物候的影响
该研究观测的寄主植物木荷只有少数个体在3月末开始开花,大多数个体约在4月中旬进入始花期,5月初达到开花高峰期,整个种群在5月下旬进入末花期,花期持续时间最短为21 d,最长为50 d,平均花期约30 d。5月中旬木荷开始坐果,整个种群在8月初进入果末期,平均果期为85 d。鞘花在木荷开花前一周进入始花期,它们的始花期与木荷的始花期相关性达到了极显著的水平(P<0.001)。鞘花的平均花期比木荷约少10 d,果熟期与木荷的果期在时间上基本重叠(图3)。
2.3 寄主与寄生生物学特性对鞘花繁殖物候的影响
2.3.1 寄主生物学特性的影响木荷的冠幅与其树高的相关性较高(r=0.556,P<0.05),但冠幅与鞘花的物候参数没有显著的相关关系。鞘花寄生的高度基本在木荷树高的一半以上,且木荷植物越高,鞘花的寄生高度也越高(r=0.627,P<0.05)。木荷的高度与鞘花的始花期和花期长短有显著的相关性,表现在木荷的高度越高,鞘花的始花期越早,花期越短(表1)。回归分析也表明木荷的高度对于鞘花的花期长短有显著的影响(R2=0.281, P<0.05)。
2.3.2 寄生生物学特性的影响表现在寄生的高度越高,相应地其冠幅也越小,果实成熟的时间越早,花期和果期都相应缩短,但是这些相关关系均未达到显著水平(表1)。
2.4 温湿度对鞘花繁殖物候的影响
物候观测期间,研究区域的月平均温度为21.4 ℃,而随着雨季的到来,月平均相对湿度为51.86%~97.19%,一直保持较为稳定的上升趋势(图4)。在雨季来临之前,鞘花的花期基本结束,果熟期则与雨季同步。每月开花/果熟的鞘花个体数量与同期和前一个月的平均温度、相对湿度的相关性均不显著。
3讨论
3.1 鞘花的繁殖物候特征
植物的繁殖物候会受到各种生物和非生物因子的影响,对于半寄生植物来说,它们从寄主植物吸取水分和养分等营养物质,又与传粉者和种子散布者相互作用。因此,其繁殖物候及其影响因素也更为复杂和特殊(Aukema, 2003; Yule & Bronstein, 2018)。鞘花与大部分桑寄生科植物一样,其开花物候类型属于集中大量开花且具有较高的花期同步性指数,这在其它桑寄生科植物中也得到了验证(Davidar, 1983b)。较高的花期同步性指数被认为具有吸引更多传粉者的优势(Primack, 1980; Augspurger, 1981; Buide et al., 2002),但是由于环境因子和基因型的变异,种群内植物的开花物候不可能达到完全同步(Elzinga et al., 2007)。
植物从开花到果实成熟是一个循序渐进的过程,因此其果期物候相应地会受到植物始花期和花期长短的影响。开花较早的鞘花个体其果熟期相应也会越早,从果实较早成熟的个体到果实较晚成熟的个体,果期的同步性增加,这在其他由脊椎动物散布的植物中也得到了验证(Eriksson & Ehrlen, 1991)。以往的研究表明果期的同步性会受到花期同步性的制约即与花期同步性相比果期同步性更低(Gorchov, 1990),但在本研究中果期的同步性却高于花期的同步性,且花期的同步性与果期的同步性还有较弱的正相关关系(r=0.115,P=0.699)。
冠幅较大的鞘花,果实的产量也越高,一般情况下果实产量越高,移除的数量也会越高。鞘花在7月末进入果末期,此时只有冠幅较大的鞘花植株上还有果实。因此,取食数量和取食速率都明显降低,这在一项关于槲寄生科的Phoradendron californicum植物上有所体现(Larson, 1996)。由于鸟类特殊的取食行为和桑寄生植物的种子能在短时间内萌发的特性(Davidar, 1983b; 肖来云和普正和, 1989),果实被取食之后的果皮数量就代表了桑寄生植物的种子被最大可能性散布出去的数量(Yule & Bronstein, 2018)。因此,果量较大的鞘花种子被散布的可能性也相对较高。
3.2 寄主物候对鞘花繁殖物候的影响
当寄主植物所积累的资源满足其自身的繁殖需求时,会促使其上感染的桑寄生植物的开花时间与其开花时间相近(Overton, 1997)。本研究中木荷始花期与鞘花始花期的相关性达到了显著性水平,且它们的花果期在时间上高度重叠,但是一项关于槲寄生科沙漠槲寄生(Phoradendron californicum)的研究却表明其与其感染的五种寄主植物物候事件的发生顺序并不一致(Yule & Bronstein, 2018)。另外,鞘花虽然先于木荷开花,但是由于木荷是雌雄同花,虫媒传粉植物(张宏达和任善湘,1988),不依赖于鸟类进行种子散布,表明木荷的繁殖物候对鞘花吸引传粉者和种子散布者作用可能并不显著。因此,仅根据鞘花与木荷之间繁殖物候的时间重叠,并不能得出木荷的繁殖物候是决定鞘花繁殖物候的唯一因素。
3.3 寄主与寄生生物学特性对其繁殖物候的影响
鞘花的花期长短受到木荷高度的影响,这可能是由于木荷植物的高度影响了鞘花的寄生高度,而间接影响了鞘花的花期长短。同时鞘花的寄生高度越高,始花期越晚,分析可能是由于鞘花开花时,雨季尚未来临,而鞘花所需要的水分和养分是通过寄主的木质部从寄主的根部向冠层运输的。在对草本植物的研究中也表明个体越高的物种的始花期通常晚于个体较矮的物种(Du & Qi, 2010)。
植物的繁殖物候受到植物大小的影响(Torimaru & Tomaru, 2006)。鞘花的冠幅越大,始花期越早,相应地花期也越长。对豆科Lotus corniculatus 植物的物候觀测也表明始花期与植物大小和花期长短高度相关即个体较大的植物开花更早并有更长的花期(Ollerton & Lack, 1998)。在野生肉豆蔻中,相较于个体较小的植株,个体较大的植株有较长的花期和较低的同步性(Otarola et al., 2013)。由此可知,植物大小会影响植物的开花物候,而植物大小和物候又会反过来影响植物的繁殖适合度(Ollerton & Lack, 1998)。
3.4 温湿度对鞘花繁殖物候的影响
热带地区植物的开花和结果物候多集中在湿季,开花物种的数量与每月的降水量是显著正相关的(Silva et al., 2011)。因此,木荷与鞘花的繁殖物候期的高度重叠可能是由于气候因子介导形成的。本研究中鞘花的繁殖物候也集中在雨季,果期长短与相对湿度有较高的相关关系,对非洲的桑寄生植物的研究也发现果期在湿季时达到高峰期(Watson, 2001),但是每月开花果熟个体数量与平均温度和相对湿度的相关性却不显著。同时,由于雨季来临降水较多,在果皮收集期间,收集到未成熟果实的数量与果皮被取食数量呈现相反的趋势。
然而,本研究也存在一定的局限性。一是在热带雨林中,林下能见度较低,物候观测较为困难,因此选取的鞘花个体数量相对较少;二是鞘花的寄生高度相对较高,而寄主植物在生长过程中可能会出现遮挡鞘花个体部分枝条的情况,从而会导致对于物候事件的发生时间未能较为准确地判断;三是鞘花个体聚集分布,单株木荷植物上感染的鞘花个体数存在较大差异,且由于寄主植物的个体数较少,因此本研究没有考虑单株木荷植物上感染的鞘花植物个体之间的繁殖物候是否存在差异。
根据对西双版纳地区桑寄生-寄主网络的调查发现,鞘花的寄主种类多达46种(王煊妮和张玲, 2017),由于寄生植物的繁殖物候会因感染到不同种类的寄主植物而不同(Yule & Bronstein, 2018),因此在未来的研究中不仅要关注多种寄主植物种类上单一寄生植物种类的繁殖物候,还要考虑物候事件的时间长度和强度(Freitas & Bolmgren, 2008),在不同的生境和气候条件下,进行长时间的物候观测(Godschalk, 1983),研究更多桑寄生植物种类的花期和果期物候。同时随着全球气候变暖,极端气候事件增加(Deb et al., 2018),植物的物候特征已经受到影响,表现在始花期普遍提前,种群内个体间的花期同步性程度降低等方面(Zohner et al., 2018)。然而,植物物候对气候变化的响应速度在不同地区和植物种类间存在较大差异(Zhao et al., 2013; Renner & Zohner, 2018),对于这类依赖鸟类散布种子的半寄生植物来说,除了延长物候观测时间以判断该植物对于气候变化的响应程度之外,还需要关注气候变暖对于其生物学特征(如株高、开花数目等)、传粉者和种子散布者的丰度等相互作用动物的影响(王颖洁等, 2017),从而全面地揭示其繁殖物候特征及其影响因素以及对气候变化的响应过程和机制。
参考文献:
AIZEN MA, 2003. Influences of animal pollination and seed dispersal on winter flowering in a temperate mistletoe [J]. Ecology, 84(10): 2613-2627.
AUGSPURGER CK, 1981. Reproductive synchrony of a tropical shrub-experimental studies on effects of pollinators and seed predators on Hybanthus prunifolius (Violaceae) [J]. Ecology, 62(3): 775-788.
AUGSPURGER CK, 1983. Phenology, flowering synchrony, and fruit set of six neotropical shrubs [J]. Biotropica, 15(4): 257-267.
AUKEMA JE, 2003. Vectors, viscin, and Viscaceae: Mistletoes as parasites, mutualists, and resources [J]. Front Ecol Environ, 1(4): 212-219.
AUKEMA JE, DEL RIO CM, 2002. Variation in mistletoe seed deposition: Effects of intra-and interspecific host characteristics [J]. Ecography, 25(2): 139-144.
BENNETTS RE, WHITE GC, HAWKSWORTH FG, et al., 1996. The influence of dwarf mistletoe on bird communities in Colorado ponderosa pine forests [J]. Ecol Appl, 6(3): 899-909.
BUIDE ML, DIAZ-PEROMINGO JA, GUITIAN J, 2002. Flo-wering phenology and female reproductive success in Silene acutifolia Link ex Rohrb [J]. Plant Ecol, 163(1): 93-103.
CANDIA AB, MEDEL R, FONTURBEL FE, 2014. Indirect positive effects of a parasitic plant on host pollination and seed dispersal [J]. Oikos, 123(11): 1371-1376.
CHANG HD, REN SX, 1988. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 49: 213-224. [張宏达, 任善湘, 1998. 中国植物志 [M]. 北京:科学出版社, 49:213-224.]
CHEN FJ, YANG YH, LIU JR, et al., 2017. Variation characteristics and the impact factors of flowering phenology of Lagerstroemia indica in urban gardens in South Sichuan [J]. Chin J Ecol, 36(7): 1841-1846. [陈发军, 杨银华, 柳继荛, 等, 2017. 川南城市园林中紫薇开花物候变化特征及其影响因素 [J]. 生态学杂志, 36(7):1841-1846.]
CLARK RM, THOMPSON R, 2011. Estimation and comparison of flowering curves [J]. Plant Ecol Divers, 4(2-3): 189-200.
CRAINE JM, WOLKOVICH EM, TOWNE EG, et al., 2012. Flowering phenology as a functional trait in a tallgrass prairie [J]. New Phytol, 193(3): 673-682.
DAVIDAR P, 1983a. Birds and neotropical mistletoes-effects on seedling recruitment [J]. Oecologia, 60(2): 271-273.
DAVIDAR P, 1983b. Similarity between flowers and fruits in some flowerpecker pollinated mistletoes [J]. Biotropica, 15(1): 32-37.
DAVIES TJ, WOLKOVICH EM, KRAFT NJB, et al., 2013. Phylogenetic conservatism in plant phenology [J]. J Ecol, 101(6): 1520-1530.
DEB JC, PHINN S, BUTT N, et al., 2018. Climate change impacts on tropical forests: Identifying risks for tropical Asia [J]. J Trop For Sci, 30(2):182-194.
DU G, QI W, 2010. Trade-offs between flowering time, plant height, and seed size within and across 11 communities of a QingHai-Tibetan flora [J]. Plant Ecol, 209(2): 321-333.
ELZINGA JA, ATLAN A, BIERE A, et al., 2007. Time after time: Flowering phenology and biotic interactions [J]. Trend Ecol Evol, 22(8): 432-439.
ERIKSSON O, EHRLEN J, 1991. Phenological variation in fruit characteristics in vertebrate-dispersed plants [J]. Oecologia, 86(4): 463-470.
FREITAS L, BOLMGREN K, 2008. Synchrony is more than overlap: Measuring phenological synchronization considering time length and intensity [J]. Rev Brasil Bot, 31(4): 721-724.
GODSCHALK SKB, 1983. The reproductive phenology of three mistletoe species in the Loskop Dam Nature Reserve, South Africa [J]. S Afr J Bot, 2(1): 9-14.
GORCHOV DL, 1990. Pattern, adaptation, and constraint in fruiting synchrony within vertebrate-dispersed woody plants [J]. Oikos, 58(2): 169-180.
HILTY SL, 1980. Flowering and fruiting periodicity in a premontane rain forest in pacific Colombia [J]. Biotropica, 12(4): 292-306.
LARSON DL, 1996. Seed dispersal by specialist versus generalist foragers: The plant’s perspective [J]. Oikos, 76(1): 113-120.
LIU ZM, JIANG DM, 2007. Progress in the study of plant reproductive phenology [J]. Acta Ecol Sin, 27(3): 1233-1241. [劉志民, 蒋德明, 2007. 植物生殖物候研究进展 [J]. 生态学报, 27(3):1233-1241.]
MATHIASEN RL, NICKRENT DL, SHAW DC, et al., 2008. Mistletoes: Pathology, systematics, ecology, and ma-nagement [J]. Plant Dis, 92(7): 988-1006.
NAPIER KR, MATHER SH, MCWHORTER TJ, et al., 2014. Do bird species richness and community structure vary with mistletoe flowering and fruiting in Western Australia? [J]. Emu-Austr Ornithol, 114(1): 13-22.
OLLERTON J, LACK A, 1998. Relationships between flowering phenology, plant size and reproductive success in Lotus corniculatus (Fabaceae) [J]. Plant Ecol, 139(1): 35-47.
OTAROLA MF, SAZIMA M, SOLFERINI VN, 2013. Tree size and its relationship with flowering phenology and reproductive output in wild nutmeg trees [J]. Ecol Evol, 3(10): 3536-3544.
OVERTON JM, 1997. Host specialization and partial reproductive isolation in desert mistletoe (Phoradendron californicum) [J]. SW Nat, 42(2): 201-209.
PRIMACK RB, 1980. Variation in the phenology of natural-populations of montane shrubs in New-Zealand [J]. J Ecol, 68(3): 849-862.
RATHCKE B, LACEY EP, 1985. Phenological patterns of terrestrial plants [J]. Ann Rev Ecol Syst,16: 179-214.
RENNER SS, ZOHNER CM, 2018. Climate change and phenological mismatch in trophic interactions among plants, insects and vertebrates [J]. Ann Rev Ecol Evol Syst, 49: 165-182.
ROOM PM, 1972. Fauna of mistletoe Tapinanthus-bangwensis (Engl & K. Krause) growing on cocoa in Ghana: Relationships between fauna and mistletoe [J]. J Anim Ecol, 41(3): 611-621.
SILVA IA, DA SILVA DM, DE CARVALHO GH, et al., 2011. Reproductive phenology of Brazilian savannas and riparian forests: Environmental and phylogenetic issues [J]. Ann For Sci, 68(7): 1207-1215.
STELTZER H, POST E, 2009. Seasons and life cycles [J]. Science, 324(5929): 886-887.
TEIXEIRA-COSTA L, COELHO FM, CECCANTINI GCT, 2017. Comparative phenology of mistletoes shows effect of different host species and temporal niche partitioning [J]. Botany, 95(3): 271-282.
TORIMARU T, TOMARU N, 2006. Relationships between flowering phenology, plant size, and female reproductive output in a dioecious shrub, Ilex leucoclada (Aquifoliaceae) [J]. Can J Bot, 84(12): 1860-1869.
WANG Q, LI LQ, LI MR, 1996. Studies on the chemical constituents of Qiaohua jisheng (Macrosolen cochinchinensis) [J]. Chin Trad Herb Drug, 21(9): 518-521. [汪强, 李良瓊, 李美蓉, 1996. 鞘花寄生化学成分的研究 [J]. 中草药, 21(9):518-521.]
WANG XN, ZHANG L, 2017. Species diversity and distribution of mistletoes and hosts in four different habitats in Xishuangbanna, Southwest China [J]. J Yunnan Univ (Nat Sci Ed), 39(4): 701-711. [王煊妮, 张玲, 2017. 西双版纳4种生境下的桑寄生与寄主植物多样性及分布特点 [J]. 云南大学学报(自然科学版), 39(4):701-711.]
WANG YJ, LU NN, YANG JY, et al., 2017. Effects of climate warming on flowering plants, pollinators and their interactions [J]. Chin J Ecol, 36(7): 2010-2018. [王颖洁, 路宁娜, 杨洁羽, 等, 2017. 气候变暖对植物、传粉者及其相互作用的影响 [J]. 生态学杂志, 36(7):2010-2018.]
WATSON DM, 2001. Mistletoe—A keystone resource in forests and woodlands worldwide [J]. Ann Rev Ecol Syst, 32: 219-249.
WRIGHT JW, MEAGHER TR, 2003. Pollination and seed predation drive flowering phenology in Silene latifolia (Caryophyllaceae) [J]. Ecology, 84(8): 2062-2073.
WRIGHT SJ, CALDERON O, 1995. Phylogenetic patterns among tropical flowering phenologies [J]. J Ecol, 83(6): 937-948.
XIAO LY, PU ZH, 1989. The propagation of Loranthaceae from Xishuangbanna [J]. Acta Bot Yunnan, 11(2): 175-180. [肖來云, 普正和, 1989. 西双版纳桑寄生植物的繁殖 [J]. 云南植物研究, 11(2):175-180.]
XIAO LY, PU ZH, 1994. Study on the relation between the spread of Loranthaceae and birds in Xishuangbanna, Yunnan [J]. Acta Ecol Sin, 14(2): 128-135. [肖来云, 普正和, 1994. 云南西双版纳桑寄生植物传播与鸟的关系研究 [J]. 生态学报, 14(2):128-135.]
YULE KM, BRONSTEIN JL, 2018. Reproductive ecology of a parasitic plant differs by host species: Vector interactions and the maintenance of host races [J]. Oecologia, 186(2): 471-482.
ZHANG KY, 1963. A preliminary analysis of climatic characte-ristics and forming factors in Southern Yunnan [J]. Acta Meteorol Sin, 33(2): 218-230. [张克映, 1963. 滇南气候的特徵及其形成因子的初步分析 [J]. 气象学报, 33(2):218-230.]
ZHAO J, ZHANG Y, SONG F, et al., 2013. Phenological response of tropical plants to regional climate change in Xishuangbanna, South-western China [J]. J Trop Ecol, 29(2): 161-172.
ZHU H, WANG H, LI BG, et al., 2015. Studies on the forest vegetation of Xishuangbanna [J]. Plant Sci J, 33(5): 641-726. [朱华, 王洪, 李保贵, 等, 2015. 西双版纳森林植被研究 [J]. 植物科学学报, 33(5):641-726.]
ZOHNER CM, MO L, RENNER SS, 2018. Clobal warming reduces leaf-out and flowering synchrony among individuals [J]. eLife, 7: 1-15.
猜你喜欢
廊坊师范学院学报(自然科学版)(2021年2期)2021-09-10 23:36:42
人与自然(2019年6期)2019-07-24 13:43:02
电子测试(2018年6期)2018-05-09 07:31:50
中成药(2018年2期)2018-05-09 07:19:32
现代园艺·综合版(2017年1期)2017-07-14 16:37:59
江苏农业科学(2017年7期)2017-05-23 23:10:25
农业与技术(2016年19期)2016-12-12 02:23:50
上海管理科学(2015年6期)2015-07-31 18:08:35
电测与仪表(2015年15期)2015-04-12 00:43:50
首都食品与医药(2014年17期)2014-11-05 02:43:16
