
长链非编码RNA PVT1在胃癌组织中的表达及对胃癌细胞凋亡的影响
2019-09-05 01:39:30黄赛亚崔旭霞景琪越张笑添
山西医科大学学报 2019年8期
王 倩,黄赛亚,崔旭霞,景琪越,张笑添*
(1山西医科大学医学影像学系,太原 030001;2山西医科大学汾阳学院医学检验系;*通讯作者,E-mail:116048164@qq.com)
近几年来,胃癌的发病率和病死率较高,严重威胁人类的健康。长链非编码RNA是长度超过200 nt的核苷酸,没有开放阅读框,且在表观遗传学修饰、转录和细胞周期调控以及凋亡过程中起重要作用[1]。人类PVT1基因又名浆细胞瘤多样异位基因1(plasmacytoma variant translocation gene 1,PVT1),其基因位于染色体8q24区且位于染色体正义链上,跨越超过300 kb的基因组间隔,该区域的异常扩增往往提示肿瘤发病风险高,因此近几年来备受关注[2]。长链非编码RNA PVT1已经发现与肝癌、乳腺癌、淋巴瘤等密切相关并影响肿瘤细胞的增殖以及凋亡等过程[3-5]。基于此,本研究旨在明确lncRNA PVT1在胃癌组织中的表达以及对胃癌细胞凋亡的影响,为寻找治疗胃癌的靶点提供依据。
1 资料与方法
1.1 一般资料
胃癌组织来自于2017-08~2018-10山西省汾阳医院手术治疗的胃癌患者18例,收集其在手术切除的胃癌组织作为胃癌组标本,并收集其癌旁(约3 cm)正常组织作为对照组。18例胃癌组织中男10例,女8例,平均年龄67岁,TNM分期Ⅰ-Ⅱ期8例,Ⅲ-Ⅳ期10例;中高分化11例,低分化7例;淋巴结转移者9例,无淋巴结转移者9例。所有患者均经过病理活检确诊,术前均未进行放化疗,本研究经过伦理委员会批准,均签署知情同意书。
1.2 主要试剂
人胃癌细胞MGC-803,BGC-823,MKN-28购自武汉博士德有限公司,人胃黏膜细胞GES-1为本实验保存;DMEM培养基购于武汉博士德有限公司;胎牛血清购于杭州四季青生物有限公司;反转录试剂盒以及荧光定量PCR试剂盒购自宝生物有限公司;siRNA以及阴性对照由广州锐博生物科技有限公司合成;Lipofectamine 2000 购自Invitrogen公司;Annexin Ⅴ-FITC/PI凋亡试剂盒购自联科生物有限公司。caspase-3单克隆抗体购于CST公司,GAPDH多克隆抗体购于Bioworld公司。
1.3 细胞培养
人胃癌细胞MGC-803,BGC-823,MKN-28和胃黏膜细胞GES-1分别培养于含10%胎牛血清的DMEM培养基,并置于37 ℃、5%CO2的培养箱中。当细胞生长至80%汇合度时,用胰酶消化传代继续培养。
1.4 细胞转染
本实验分为三组,分别是PVT1-siRNA组、siRNA-NC组和空白对照组。转染前1 d将胃癌细胞置于六孔板中培养,细胞密度大约为3.0×105个/孔,第2天细胞融合度大约为60%时进行转染。分别用无血清DMEM培养基孵育siRNA(75 nmol)和lip2000(5 μl),将两者混匀室温静置10 min后,各自缓慢加于PVT1-siRNA组和siRNA-NC组的胃癌细胞MKN-28中,而空白组不作任何处理,各组在7 h后将无血清培养基更换为含10%胎牛血清的DMEM培养基中继续培养48-72 h进行荧光定量PCR和流式细胞术等实验。
1.5 荧光定量PCR检测胃癌组织和胃癌细胞中的lncRNA PVT1
使用Trizol试剂提取胃癌组织和癌旁组织、胃癌细胞MGC-803,BGC-823,MKN-28和胃黏膜细胞GES-1中的总RNA,将总RNA反转录为cDNA后,以GAPDH为内参进行荧光定量PCR实验。lncRNA PVT1的扩增条件是:95 ℃,5 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,40个循环,反应结束后进行熔解曲线分析,采用2-ΔΔCt法计算细胞中lncRNA PVT1的水平。并筛选lncRNA PVT1表达最高的胃癌细胞作为后续的研究对象。
1.6 流式细胞术检测细胞凋亡
本实验分为三组,分别是PVT1-siRNA组,siRNA-NC组和空白对照组。细胞转染48 h后,使用不含EDTA的胰酶消化,1 000 r/min低速离心并收集各组细胞,用预冷的PBS洗涤细胞两次,并按Annexin Ⅴ-FITC/PI双染凋亡检测试剂盒说明书进行操作,在30 min内使用流式细胞仪检测细胞凋亡情况。
1.7 Western blot实验检测caspase-3蛋白的表达
本实验分为三组,分别是PVT1-siRNA组,siRNA-NC组和空白对照组。细胞转染72 h后,RIPA裂解法提取胃癌细胞MKN-28中的总蛋白,使用BCA法检测蛋白质的浓度。取30 μg总蛋白进行12%SDS-PAGE凝胶电泳,湿转2 h至PVDF膜上,5%脱脂奶粉封闭2 h后一抗(caspase-3抗体1 ∶3 000,actin抗体1 ∶5 000)4 ℃孵育过夜,二抗室温孵育1 h,TBST洗膜3次后,使用ECL化学发光法呈色,各条带灰度值采用ImageJ软件进行扫描并记录。
1.8 统计学分析
2 结果
2.1 lncRNA PVT1在胃癌组织和胃癌细胞中的表达
荧光定量PCR检测结果显示,胃癌组织中lncRNA PVT1的水平显著高于癌旁组织(P<0.01,见图1A);同时与胃黏膜细胞GES-1相比,胃癌细胞MKN-28和BGC-823中的lncRNA PVT1的表达明显增高(P<0.01,见图1B),而其中胃癌细胞MKN-28中lncRNA PVT1的表达是最高的,故以胃癌细胞MKN-28作为后续的研究对象。

图1 lncRNA PVT1在胃癌组织和胃癌细胞中的表达情况Figure 1 The expression of lncRNA PVT1 in gastric cancer tissue and gastric cancer cells
2.2 胃癌组织中lncRNA PVT1的表达与临床病理参数之间的关系
胃癌组织中lncRNA PVT1的表达在TNM分期为Ⅲ-Ⅳ期的水平显著高于TNMⅠ-Ⅱ期的水平(P<0.05),淋巴结转移患者其水平显著高于无转移患者(P<0.05),中高分化患者其水平显著低于低分化患者的水平(P<0.05),但lncRNA PVT1的水平在不同年龄、不同性别间差异无统计学意义(P>0.05,见表1)。



病理参数nlncRNA PVT1t P性别 男101.517±0.251.0750.298 女81.631±0.18年龄 <60岁61.707±0.4991.1550.265 >60岁121.446±0.32TNM分期 Ⅰ-Ⅱ81.019±0.2663.3100.004 Ⅲ-Ⅳ101.443±0.272淋巴结转移 无91.021±0.3872.9830.008 有91.544±0.355分化程度 中高分化110.792±0.9343.3950.003 低分化72.150±0.605
2.3 沉默胃癌细胞MKN-28中的lncRNA PVT1的水平
为了鉴定siRNA的干扰效率,在转染48 h后收集细胞,荧光定量PCR检测胃癌细胞MKN-28中lncRNA PVT1的水平,与si-NC组相比,PVT1-siRNA能显著下调胃癌细胞MKN-28中lncRNA PVT1的水平(P<0.01),而空白对照组和si-NC组之间差异没有统计学意义(P>0.05,见图2)。

与si-NC组相比较,**P<0.01图2 荧光定量PCR检测胃癌细胞的转染效率Figure 2 The tranfection efficiency of gastric cancer cell detected by qRT-PCR
2.4 沉默lncRNA PVT1后对胃癌细胞MKN-28凋亡的影响
转染48 h后流式细胞仪检测结果显示si-PVT1组的细胞凋亡率明显高于si-NC组和空白对照组(P<0.01,见图3),而空白对照组和si-NC组则没有明显差异。该结果说明,沉默lncRNA PVT1的表达后可以诱导胃癌细胞发生凋亡。
2.5 lncRNA PVT1对胃癌细胞caspase-3蛋白水平的影响
Western blot结果显示,沉默胃癌细胞MKN-28中的lncRNA PVT1后,caspase-3蛋白的表达显著高于si-NC组和空白对照组(P<0.01),而空白对照组和si-NC组则没有明显差异(见图4),该结果表明lncRNA PVT1可能是通过抑制caspase-3蛋白的表达进而抑制胃癌细胞发生凋亡。

图3 沉默lncRNA PVT1对细胞凋亡的影响Figure 3 The effect of silencing lncRNA PVT1 on cell apoptosis in gastric cancer cell
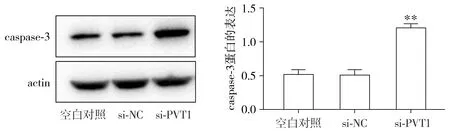
与si-NC组和空白对照组相比较,**P<0.01图4 Western blot检测caspase-3蛋白的表达Figure 4 The protein expression of caspase-3 by Western blot
3 讨论
近年来大量研究表明lncRNA在细胞生长和人类的疾病发展过程中发挥着重要的调控作用[6,7]。lncRNA PVT1在多种肿瘤中的表达增加[8,9],并影响着肿瘤的发生和发展。
已有研究表明,lncRNA PVT1与胃癌密切相关[10,11],为了研究lncRNA PVT1在胃癌发生发展中所起的作用,在本研究中我们检测了胃癌和癌旁组织,以及胃癌细胞和胃黏膜细胞中lncRNA PVT1的水平,结果发现胃癌组织中的lncRNA PVT1显著高于癌旁组织,并且lncRNA PVT1的表达与胃癌的TNM分期、淋巴结转移以及分化程度密切相关,TNM为Ⅲ-Ⅳ期的胃癌组织中lncRNA PVT1的表达显著高于TNM为Ⅰ-Ⅱ期;lncRNA PVT1在有淋巴结转移的胃癌组织中的表达显著高于无淋巴结转移的胃癌组织;分化程度低的胃癌组织中的lncRNA PVT1的表达显著高于中、高分化胃癌组织,其他学者[12]也得到了类似的结果,并且推断lncRNA PVT1可能募集PRC2蛋白复合物到p15、p16基因的启动子区,介导组蛋白H3K27位点甲基化进而抑制p15、p16的表达,促进了胃癌的生长,该结果进一步提示lncRNA PVT1可能参与了胃癌的发生和发展。另外,我们在前期的研究中也发现了lncRNA PVT1可以促进胃癌细胞迁移和增殖[13],进而促使了胃癌的发生发展,但是lncRNA PVT1是否参与胃癌细胞的凋亡过程尚不十分清楚。因此在本研究中,沉默胃癌细胞中的lncRNA PVT1的表达后,使用流式细胞术检测胃癌细胞的凋亡率,结果发现胃癌细胞MKN-28中的lncRNA PVT1的水平下调后,胃癌细胞MKN-28的凋亡率比对照组明显增高。已有研究证实,lncRNA PVT1可以通过不同的机制影响细胞的凋亡过程,沉默结肠癌细胞中的lncRNA PVT1后,可以激活TGF-β信号通路并促进结肠癌细胞凋亡[9]。还有研究证实lncRNA PVT1能抑制TGF-β信号通路蛋白SMAD4的表达,抑制PARP和caspase-3的断裂和激活,抑制细胞凋亡[14]。而在膀胱癌中,lncRNA PVT1则是通过抑制caspase-3的表达进而抑制细胞凋亡[15]。在非小细胞肺癌中,lncRNA PVT1则通过调控miR-497的表达而抑制肺癌细胞凋亡[16]。在鼻咽癌[17]中lncRNA PVT1则是通过抑制caspase-3、caspase-9以及PARP的表达进而抑制细胞凋亡,从而促进了肿瘤的进程。而其中caspase家族是凋亡途径中的关键成分,caspase因子的激活可触发凋亡途径,从而产生级联效应。因此在本研究中,我们检测了凋亡相关蛋白caspase-3的表达,结果显示沉默胃癌细胞中的lncRNA PVT1后,caspase-3蛋白的表达增高,说明敲低胃癌细胞MKN-28中的lncRNA PVT1后,导致凋亡途径蛋白caspase-3的激活,进而促进了胃癌细胞的凋亡。
综上所述,lncRNA PVT1可能作为癌基因促进胃癌的发生和发展,并且可以抑制胃癌细胞凋亡,因此有望成为胃癌诊断和治疗的靶点。
猜你喜欢
清华金融评论(2022年4期)2022-04-13 21:33:11
国际放射医学核医学杂志(2021年10期)2021-02-28 08:43:46
看世界·学术下半月(2020年7期)2020-09-10 07:22:44
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:06
中成药(2018年12期)2018-12-29 12:25:44
广东农业科学(2017年10期)2018-01-25 03:22:15
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年6期)2017-06-13 07:30:35
医学研究杂志(2015年4期)2015-06-10 06:42:43
甘肃农业科技(2015年12期)2015-04-22 02:57:24
