
皮氏菊头蝠(Rhinolophus pearsoni)的栖息生态特征
2019-09-04 10:25:02龚小燕黄太福张佑祥彭清忠刘志霄
生态学报 2019年15期
龚小燕,黄太福,吴 涛,张佑祥,彭清忠,刘志霄
吉首大学生物资源与环境科学学院, 吉首 416000
溶洞(Karst cave)是由于喀斯特作用而形成的地下空腔,其内部环境黑暗,小气候相对稳定,是许多动物类群(尤其是蝙蝠)栖息、繁育及避敌的适宜场所[1- 2]。
近20多年以来,栖息地选择已逐渐成为种群生态学研究的热点,而由于蝙蝠具有特殊的形态、生理、生态与行为习性[3],其栖息地选择的研究受到越来越多的重视[4- 7]。
皮氏菊头蝠(Rhinolophuspearsoni)又名绒毛菊头蝠,隶属于翼手目(Chiroptera)菊头蝠科(Rhinolophidae)菊头蝠属(Rhinolophus),广泛分布于我国南方及其邻近区域[8],是典型的洞栖食虫性蝙蝠,为洞穴生态系统的关键种类,具有重要的生态生物学研究价值[9]。目前,关于皮氏菊头蝠的研究主要涉及形态、回声定位、捕食行为、心电图、分类分布等方面[10- 15],而有关其在溶洞中的栖息生态特征还缺乏系统的研究报道。鉴于该蝠分布的广泛性、代表性及生态意义的普遍性,我们结合湘西州和张家界市溶洞众多的生态环境与资源特点[16- 17],对其栖息生态特征进行了较为系统的研究,以期为洞栖性蝙蝠栖息生态学的比较研究及物种保护实践提供基本资料。
1 研究区域简况与研究方法
1.1 湘西州与张家界市简况
湘西土家族苗族自治州(109°10′—110°22.5′E,27°44.5′—29°38′N,简称湘西州)位于武陵山脉东部,地处我国动物地理区划中的华中区、华南区和西南区的交汇地带,总面积约1.5万km2,受多条河流影响,境内喀斯特地貌发育良好,溶洞广布,属中亚热带季风性湿润气候[16],适宜于蝙蝠的栖息和繁衍。张家界市(109°40′—111°20′E,28°52′—29°48′N)位于湖南省西北部及武陵山区的腹地,地处云贵高原与洞庭湖沉降区的结合部,总面积约0.97万km2,地形复杂,沟谷众多,水系发达,岩溶地貌发育良好,属中亚热带山原型季风性湿润气候[17]。
1.2 研究方法
1.2.1物种鉴别
在前期调查中,项目组成员已经积累了丰富的蝙蝠野外辨识经验,可依据体型、毛色、栖息行为等特征辨认域内常见的蝙蝠种类。本文的研究对象(皮氏菊头蝠)体型中等偏大,毛色深棕或棕褐色[9,18],栖息时翼膜通常紧紧地包裹着身体,仅稍露背部及头部,与其他的蝠种易于区别。
1.2.2观测方法、溶洞类型划分与数据分析
近年,项目组在湘西州(吉首市、永顺县、古丈县、花垣县、龙山县和凤凰县)及张家界市境内进行生物资源本底调查的过程中,观察到不同种类的蝙蝠之间存在栖息行为及生态习性方面的明显差异。基于前期调查,我们自2016年1月起,选择了25个溶洞对皮氏菊头蝠的栖息生态特征进行了观测。进洞考察时,采用强光灯扫视肉眼可达之处,对观察到的蝙蝠的相关数据进行测量记录。通过尼康D800单反相机对蝙蝠的栖息生态特征进行拍摄或录相。通过高性能的智能手机内置的软件功能记录溶洞的GPS及海拔数据。对于洞道的长度、栖点(roostpoint,蝙蝠在洞道中的具体栖挂位点)高度、栖点离洞口的距离、栖点温度(roostsite temperature,蝙蝠栖挂点附近的岩壁温度)及蝙蝠的体温则分别采用UT391+激光测距仪(量程60 m,精确度±1.5 mm)和标智GM700手持式激光测温仪(测量范围-50—700℃,精确度±1.5℃)进行测量。测量体温时,将仪器置于离蝙蝠身体1—2 cm处,以减少对蝙蝠的干扰并保证数据的有效性与同一性。
依据洞道长度及复杂程度将溶洞划分为三类:Ⅰ类:洞道长度≤100 m,洞道结构简单,为单洞道;Ⅱ类:洞道长度介于100—500 m,洞道结构相对复杂,具分支;Ⅲ类:洞道长度≥500 m,洞道结构复杂,洞道分支多且分层,具有多个明显或隐蔽的洞口。
每个溶洞的地理位置、内部结构、开发利用程度及周边的人文景观差异较大,我们通过观察洞外植被、洞内景观受破坏的程度,结合对当地居民的问卷调查等综合信息将洞内蝙蝠受人为干扰的程度划分为三级:“强”(洞内钟乳石破坏较大,人为垃圾较多,常有人进洞游玩)、“中”(洞穴处于公路或村落附近,洞周植被破坏严重,偶有牲畜或人进洞避雨或玩耍)、“弱”(位置较为偏僻或洞口较为隐蔽或危险性较高,洞口植被相对较好,鲜有人进洞)。
依据蝙蝠栖挂点离地面的高度(栖点高度roostpoint height)及其隐蔽性进行栖点安全性评价:“安全性高”,栖点位置较高(大于3 m)或栖点位于较为隐蔽的地方,蝙蝠不太容易受到人及其他动物的伤害或不易被发现;“安全性中”,栖点离地面的高度在2—3 m之间,一般身高的人需借助工具才可伤害到蝙蝠;“安全性低”,栖点高度在2 m以下,蝙蝠容易受到人或其他动物的干扰或伤害。
调查期间,观察到自然光射入洞内的最大深度通常不超过40 m,因此,我们以40 m作为间距予以洞段划分,共分为13个洞段:0—40、40—80、80—120、120—160、160—200、200—240、240—280、280—320、320—360、360—400、400—440、440—480、480—520 m;通常人伸手及踮脚的一般高度在2 m左右,故将2 m作为栖点高度划分的依据,共分8个区段:0—2、2—4、4—6、6—8、8—10、10—12、12—14、14—16 m。
采用Excel软件功能对栖息行为、栖点、体温等数据进行整理与初步统计,使用SPSS 22.0中的独立样本t检验与单因素方差分析法(ANOVA)对皮氏菊头蝠的体温与栖点温度进行显著性检验、相关性分析与作图。
2 结果与分析
2.1 种群数量及分布情况
在25个溶洞中,总共记录到皮氏菊头蝠450只次(Individual-by-times, IBT),分布海拔介于248—911 m,约86%的只次栖于Ⅲ类溶洞,而栖于Ⅰ类和Ⅱ类溶洞中的只次分别为5%和9%(表1)。在湘西州和张家界市的溶洞中全年均可发现该蝠的栖息,常见其独栖或集数只至20只左右的小群栖息。
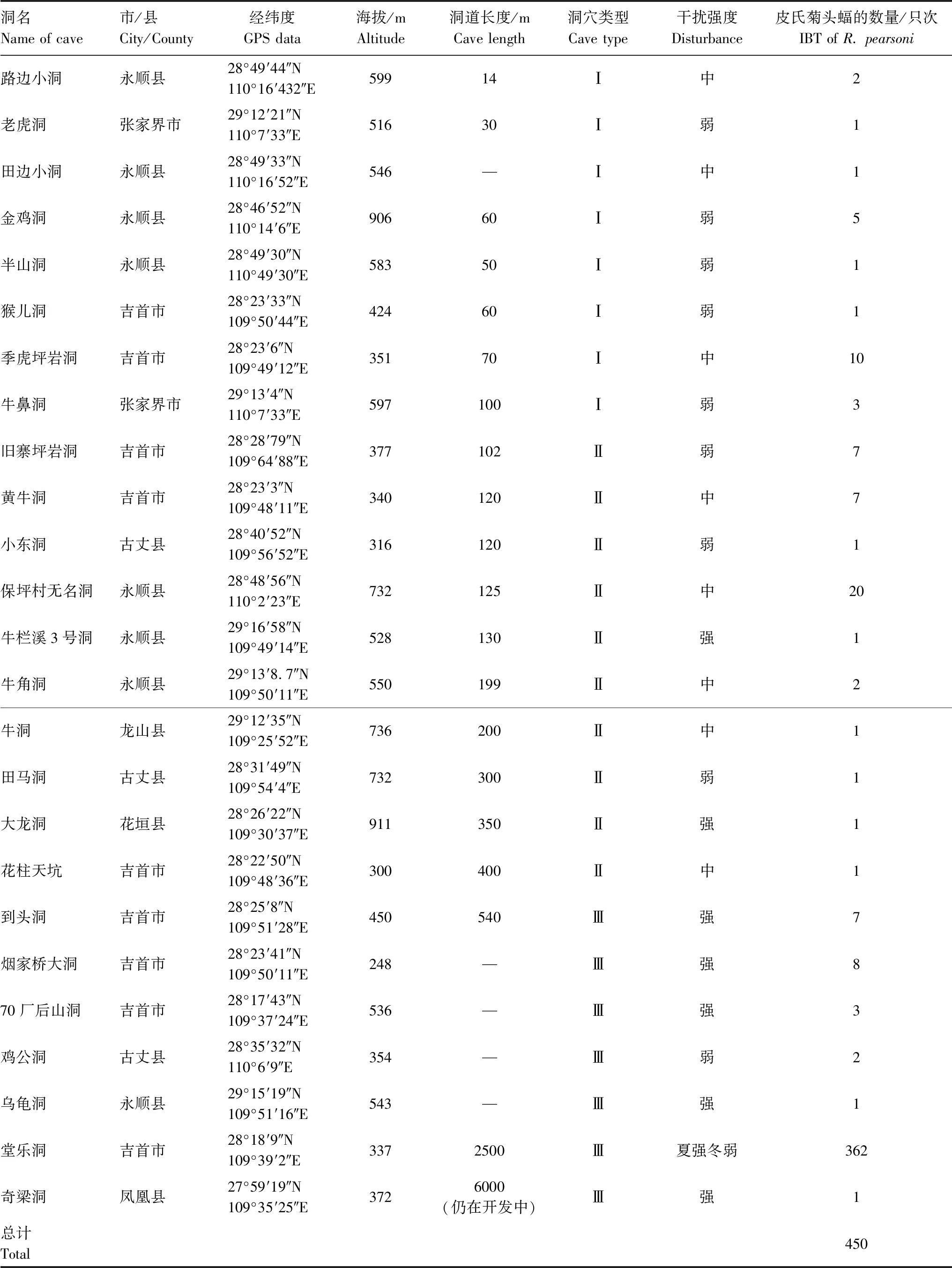
表1 所调查溶洞的基本情况
IBT:只次 Individual-by-times;Ⅰ:Ⅰ类洞穴(洞道长度≤100 m,洞道结构简单,为单洞道)Cave type Ⅰ(cave length≤100 m, singlar and simple cave tunnel);Ⅱ:Ⅱ类洞穴(洞道长度介于100—500 m,洞道结构相对复杂,具分支)Cave type Ⅱ(cave length 100—500 m, relatively complex and branching cave tunnel);Ⅲ:Ⅲ类洞穴(洞道长度≥500 m,洞道结构复杂,洞道分支多且分层,具有多个明显或隐蔽的洞口)Cave type Ⅲ(cave length ≥500 m, very complexand multi-branching-storied cave tunnel)
2.2 栖息位置及姿势
皮氏菊头蝠栖点离洞口的平均距离为(233.0±125.84) m(n=450),主要集中在440 m之内(占99.3%),其中在200—280 m范围内的个体数较多,约占1/3(33%),而离洞口440 m以上的洞段,其分布数量很少(仅为0.7%)。栖点的平均高度为(6.67±12.54) m(n=450),个体的栖点高度通常在2—10 m之间(84%),其中在6—8 m范围内的个体数较多(34%),而栖点离地面的高度超过10 m之后,所观察到的个体数很少(6.7%),栖高在2 m以下的个体数也仅占9.3%(图1)。
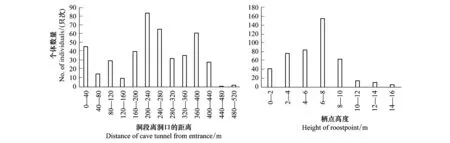
图1 皮氏菊头蝠栖点离洞口的距离及栖点高度的分布(只次/individual-by-times, IBT)Fig.1 Distribution of roostpoint distance from cave entrance and height from cave ground for Rinolophus pearsoni
该蝠种主要栖挂于洞道的侧壁面(64%),其余则栖挂于洞道顶壁的岩板面、凹窝内及钟乳石的下段或尖端(36%)(表2),通常采用“双足爪倒挂身体”(简称“双足倒挂”)的姿势栖息(63.8%),但“单足爪倒挂身体”(简称“单足倒挂”)栖息的个体也较常见(36.2%),其中使用左足倒挂(20.2%)的频率稍高于右足(16%)。约64%的个体的栖点“安全性高”,而“安全性低”和“安全性中”的栖点分别为17.6%和18.4%。

表2 皮氏菊头蝠(Rinolophus pearsoni)的栖点位置分布
注:①:洞道的顶壁岩板面 Cave ceiling;②:洞道的侧壁面 Side walls of cave;③:狭缝间 Crevices;④:凹窝内 Pits;⑤:钟乳石的下段或尖端 stalactite tips;IBT:只次 Individual-by-times;—:无 No content
2.3 体温与栖点温度
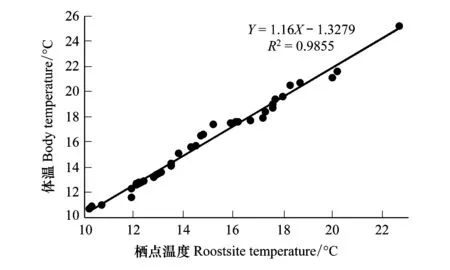
图2 皮氏菊头蝠体温与栖点温度之间的线性关系 Fig.2 Linear correlation between body and roostsite temperature(℃) of Rinolophus pearsoni
通过对皮氏菊头蝠4个部位(翼膜、背毛、耳部和鼻部)的温度进行ANOVA单因素方差分析,结果显示:它们之间不存在显著性差异(P>0.05,F=0.204,P=0.893),变化也缺乏规律性,其体温介于10.7—25.2℃之间,平均值为(15.9±3.5)℃(n=38),而栖点温度的变化范围是10.2—22.7℃,平均值为(14.9±3.0)℃(n=38)。通过独立样本t检验对该蝠栖点温度与体温进行分析,结果显示:体温和栖点温度之间不存在显著性差异(P>0.05,F=1.468,P=0.230),但体温(Y)总是稍高于栖点温度(X)(平均值之差为1℃),并且两者之间呈线性正相关,其关系式为:Y=1.16X-1.3279(X∈[10.7, 25.2],R2=0.9855,图2)。
3 讨论
3.1 皮氏菊头蝠的栖点选择
皮氏菊头蝠作为典型的洞栖性蝙蝠,通常在溶洞中蛰伏及繁育[19]。在湘西州及张家界市境内,其栖息的溶洞海拔可能介于200—1000 m之间,这也是域内绝大多数山体的海拔高度范围。溶洞通常位于山体内,因此一般也笼统地被称为“山洞”或“岩洞”。该蝠喜栖于洞道的侧壁,离地高度通常在2 m以上,绝大多数个体栖息于洞道长而结构复杂的溶洞。洞道宽阔、分支多而分层,可供蝙蝠选择的栖挂位点及微生境丰富多样,既有利于降低种内及种间的空间竞争,也有利于其隐藏、避敌、繁育及冬眠,特别是在受到人类及其他动物干扰时,蝙蝠可随时变换位置而无需离开该洞,以减少能耗。
大菊头蝠(Rhinolophusluctus)和西南鼠耳蝠(Myotisaltarium)的栖点距洞口的平均距离分别为(38.00±26.93) m(n=20)和(118.30±71.32)m(n=361),栖点高度的平均值分别为(2.24±1.73) m(n=20)和(4.06±2.14) m(n=361)。将皮氏菊头蝠与大菊头蝠[20]相比,前者的栖点通常离洞口的距离((233.03±125.84) m,n=450)较远而离地面的位置较高((6.67±12.54) m,n=450),较常栖挂于钟乳石的尖端或下段,而后者较常见于洞道的凹窝处。与西南鼠耳蝠[21]相比,皮氏菊头蝠的栖点在洞道中的位置也较深,离地面的距离也较高。总体上,这3种蝙蝠都主要选择在离洞口较近的洞段分散或独栖冬眠,栖点大多位于洞道的侧壁,大菊头蝠[20]最靠近洞口,栖点离地面的位置也最低,西南鼠耳蝠也多在洞口段冬眠,栖点位置较为多样,而皮氏菊头蝠多选择在洞道较深及位置较高处冬眠,钟乳石尖端或下段是其常见的栖挂点。此外,溶洞中的气象因子(温、湿度和CO2含量)存在时空异质性[22],而3种蝙蝠的微环境温度(即栖点温度)也存在差异,其中大菊头蝠((10.62±2.05)℃,n=14)和西南鼠耳蝠((10.35±4.14)℃,n=113)的栖点温度较为接近,皮氏菊头蝠((14.9±3.0)℃,n=38)的相对较高,表明3种蝙蝠喜居的洞段存在温度上的差异,皮氏菊头蝠所栖洞段的温度较高,洞段也较深。可见,这3种蝙蝠在栖点选择方面存在分化现象,显然对于它们与其他种类的蝙蝠(如中华菊头蝠Rhinolophussinicu、大蹄蝠Hipposiderosarmiger等)的空间及时间生态位分化的系统深入研究有助于揭示洞栖性蝙蝠的共存机制[23]。
3.2 皮氏菊头蝠的栖挂姿势
皮氏菊头蝠与大菊头蝠的栖挂姿势相似,在自然深睡的状态下翼膜紧紧地包裹着身体,仅露出少部分的头部及背部(图3a、b),但大菊头蝠个体较大,外观上更为狭长笔直,易于识别,并且皮氏菊头蝠“单足倒挂”的姿势(36.2%)较大菊头蝠多(14%)。虽然皮氏菊头蝠也可集小群育幼或冬眠(图3c、d),但并不像中华菊头蝠(R.sinicus)那样集群冬眠时个体之间紧靠在一起,而在自2007年以来的洞穴调查过程中,我们从未见到大菊头蝠2只或多只在一起冬眠或栖息的情况(母仔除外),通常一个溶洞内仅有一只大菊头蝠,即使不只一只,它们之间的距离也很远。西南鼠耳蝠的栖息姿势及栖点则较为多样,除了悬空倒挂外,还常见其以腹部贴壁倒挂、呈匍匐状卧伏于横缝间或挤进呈垂直状的竖缝间等隐蔽处冬眠[21]。不同蝙蝠之间栖挂姿势及集群模式方面的差异,可能需要从体制构造、行为遗传、进化生态,尤其是骨骼系统、肌肉系统等功能形态学或生态功能学,以及翼手类行为或进化生态学角度予以理解,这需要进一步的研究。

图3 皮氏菊头蝠的栖挂姿势及冬眠群Fig.3 Roosting posture and hibernating group of Rinolophus pearsonia:双足倒挂在洞侧壁上的姿势 Upside-down roosting posture by two-feet hanging under the side wall of karst cave;b:单足倒挂在钟乳石下段的姿势 Upside-down roosting posture by one-foot hanging under the tip section of stalactite;c:堂乐洞石壁长廊洞段的冬眠群 Hibernating group in Shibichanglang section of Tangle cave;d:永顺县芙蓉镇保坪村无名洞的冬眠群 Hibernating group in Wumingdong cave, Baoping village, Furong town, Yongshun county
3.3 皮氏菊头蝠冬眠期的体温及栖点温度
冬眠是动物应对冬季寒冷及食物缺乏的一种行为及生理机制,期间存在着间期性或周期性短暂觉醒现象[3]。冬眠期自然醒来或受到干扰后,蝙蝠通常会飞往洞道的深处,寻找隐蔽性更好或更高的栖点继续冬眠,因此,复杂的洞道系统有利于其随时调整栖位,找到安全合适的栖点,节省能量,但无论栖点位于何处,处于自然状态下的蝙蝠的体温都稍高于其环境温度约1℃。可是,在受到人为干扰时,其体温会逐渐升高,当体温升高到18—20℃以上时,多数蝙蝠会醒来飞走。
测量及分析发现,皮氏菊头蝠的平均体温及体温变化范围均高于大菊头蝠[20]和西南鼠耳蝠[21],而大菊头蝠与西南鼠耳蝠的平均体温非常接近,其主要原因可能是,后两者的栖点通常靠近洞口,而前者的栖点离洞口更远,受洞外气温的影响更小。
3.4 皮氏菊头蝠的种群数量变化及种群保护
冬眠期间,皮氏菊头蝠的数量稍多,在同一处可集成数只至20左右的小群冬眠,但更常见的是独栖冬眠,而夏秋时节在溶洞中日眠的数量明显减少,仅偶见其于偏僻洞穴中集小群育幼或独栖,这一方面可能是皮氏菊头蝠存在迁移现象,可随季节或月份气候及食物条件的变化而迁往其他区域或迁至同一区域的其他洞穴栖息,另一方面也可能受到人为干扰因素的影响,繁育期间更多地选择人迹罕至的僻静洞穴育幼而避免人类的干扰。
洞栖性蝙蝠在洞穴中冬眠、日眠及育幼,依赖洞穴环境生存及繁衍[24- 25],因此保护好天然溶洞是洞栖性蝙蝠种群保护的根本所在,也是自然生态保护的重要方面,应该引起地方政府及有关部门的足够重视,总体而言,因地制宜,采取切实可行的措施减少人类对天然溶洞的干扰与破坏是保护洞栖性蝙蝠的有效途径。
致谢:彭乐、王敏、郑小芳、向颖、肖雨晴等同学参加了部分野外工作,特此致谢。
猜你喜欢
湖南水利水电(2021年6期)2022-01-18 06:07:30
珠江水运(2021年19期)2021-11-08 11:14:26
建材发展导向(2021年16期)2021-10-12 05:39:58
石家庄铁路职业技术学院学报(2018年2期)2018-07-12 08:36:32
小溪流(画刊)(2016年12期)2017-02-04 18:34:54
水利科技与经济(2016年8期)2016-04-22 03:41:26
微型小说选刊(2015年5期)2015-06-05 09:15:29
作文周刊·小学一年级版(2015年43期)2015-05-30 10:48:04
小星星·阅读100分(低年级)(2015年1期)2015-04-07 07:36:04
小猕猴智力画刊(2015年3期)2015-04-02 11:56:43
