
土壤温度对设施葡萄枝条芽萌发和需热量的影响
2019-09-02 14:01罗国安张亚红孙利鑫张晓丽耿晓玲
江苏农业科学 2019年5期
罗国安 张亚红 孙利鑫 张晓丽 耿晓玲
摘要:以5年生红地球葡萄为材料,在设施葡萄揭盖蓄热后进行根域加热处理,并记录红地球葡萄枝条芽萌发情况,采用6种需热量模型统计芽的需热量值,并进行对比分析。结果发现,土壤温度提高,葡萄枝条芽萌发的时间和需热量均显著减少,且塑料大棚内的效果较日光温室更好;土壤温度相同时,日光温室内的红地球葡萄枝条芽的萌发时间较塑料大棚均较小,但需热量值则较大。结果表明,葡萄枝条芽的萌发和需热量与根域温度关系密切,将为设施葡萄的促早栽培提供参考。
关键词:设施葡萄;根域加热;萌芽;需热量
中图分类号: S663.104 文献标志码: A 文章编号:1002-1302(2019)05-0110-04
收稿日期:2017-10-24
基金项目:国家自然科学基金(编号:31360493)。
作者简介:罗国安(1992—),男,安徽宿松人,硕士研究生,研究方向为园艺设施与环境调控。E-mail:1558553656@qq.com。
通信作者:张亚红,博士,教授,主要从事农业气象与环境调控方面的研究工作。E-mail:zhyhcau@sina.com。
设施栽培是鲜食葡萄种植的主要方式,利用日光温室、塑料大棚等保护设施来调控环境条件,实现果品成熟期提前和品质提高。这种栽培方式自20世纪70年代在我国北方地区发展[1],已成为果农收入提高的重要途径。
同其他落叶果树一样,设施葡萄芽的萌发时间理论上由2个因子控制:一是休眠期的需冷量,只有滿足一定的低温积累才能打破芽的自然休眠,反之则会造成营养生长和开花结实的异常[2];二是萌芽期的需热量,设施葡萄在休眠结束之后,需要有一定的热量积累才能正常地萌芽展叶。葡萄的促成栽培主要是对休眠期的调控,即通过破休眠剂或集中预冷来提早结束休眠,以前的研究也多集中在休眠期的需冷量,如杨天仪等利用0~7.2 ℃模型[3]、高东升等利用犹他模型[4]、章镇等利用0~7.2 ℃模型[5]、王海波等利用3种不同模型[6]测定不同品种葡萄的需冷量。关于萌芽期的需热量较少,王西成等利用生长度时模型和有效积温模型估算江苏14个设施品种葡萄的需热量[7],奚晓君等则利用生长度时模型估算上海4个设施品种葡萄的需热量[8]。
以上研究均以需热量和需冷量值为基础,进而研究它们的关系,但未涉及土壤温度,而关于土壤温度与落叶果树生长发育的研究有:司海娣研究发现土温和气温的差异性导致日光温室中葡萄比塑料大棚中的萌芽早[9],王连荣等将设施早露蟠桃地上部接受正常自然休眠,根系接受不同温度处理,发现高温可以使花芽提前解除休眠[10],王世平等研究发现桃促成栽培早期土壤温度提高10 ℃,各物候期均早于未加温处理2~5 d[11]。为探讨土壤温度对设施葡萄枝条芽的萌发和需热量的影响,在枝条芽自然休眠结束后,设置不同的土壤温度,研究土壤温度对芽的萌发和需热量的影响。
有研究已对土壤温度对枝条芽萌发的生理及需热量的影响进行了初步探讨[12-15],在上述研究的基础上,本试验对根域温度与葡萄枝条芽的萌发和需热量的关系进行研究。
1 材料与方法
1.1 试验地概况及材料
宁夏永宁县小任果业有限公司位于宁夏银川市永宁县胜利乡,所采用的塑料大棚长96 m,跨度16 m,脊高4 m,钢架结构,覆盖和保温材料分别为PE膜和棉被,供试葡萄为红地球(Vitis vinifera),2007年种植,南北行向,株行距:0.5 m×1.3 m。日光温室长88 m,跨度9 m,脊高4 m,钢架结构,覆盖和保温材料分别为PE膜和棉被,供试葡萄为红地,2006年种植,东西行向,株行距:0.5 m×1.3 m。
1.2 试验时间及地点
试验于2014—2016年的11月至4月在宁夏永宁县小任果业有限公司葡萄基地进行。
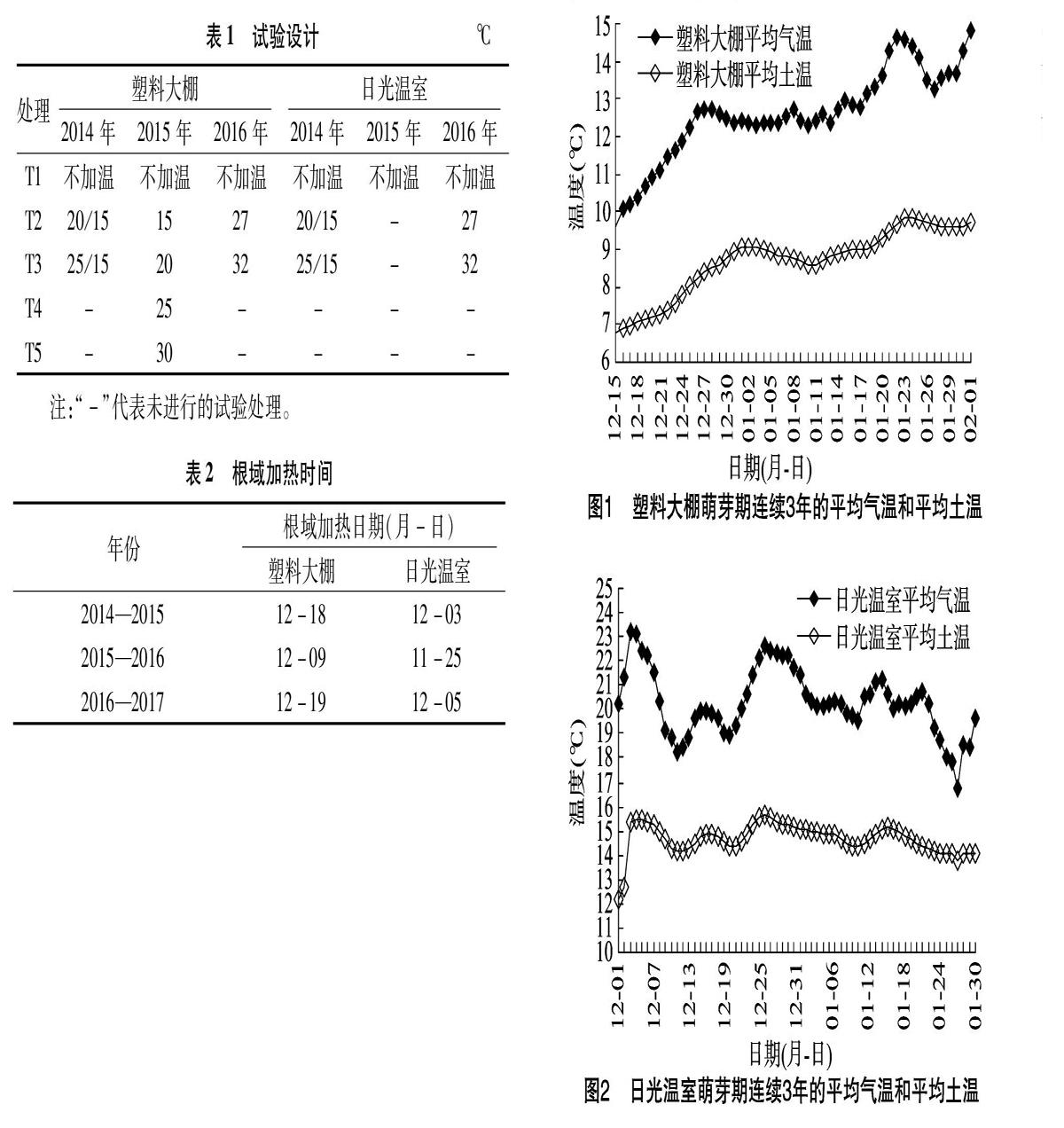
1.3 根域温度加热试验
连续3年对塑料大棚和日光温室内红地球葡萄扣棚反保温管理,休眠期间将棚内气温控制在0~7.2 ℃,以满足红地球葡萄需冷量。塑料大棚和日光温室分别在12月初进行升温管理(保温被白天揭开夜间覆盖),同时对土壤温度做以下处理:选择3个大小相同区域,其中2个区域分别将土壤温度利用发热电缆加热(表1),选择生长良好的10行葡萄(共250株),在距离葡萄主根40 cm处地表下30 cm铺设电热线(电热线固定在纳米材料板上)之后覆土,覆盖黑色地膜保温,外接控温仪控制温度(控温仪设置断电温度为理论温度+1 ℃)。另外区域作常温处理(CK)即地面覆盖黑色地膜,维持正常土温。安装加热设备后,在每个处理地表下(离葡萄根系30 cm)垂直方向10、20、30 cm处埋设温度探头,重复2次,试验采用完全随机区组设计,2行(50株)为一小区,重复3次,每个处理间设置保护行。
1.4 空气温度和土壤温度的监测
日光温室:采用美国Campbellsci公司的CR800数据采集器和相关传感器, 对CK行1.5 m处的气温和0.2 m 深度的土温进行测定。美国Campbellsci公司的CR10X-2M数据采集器和相关传感器,对处理行1.5 m处的气温和0.2 m深度的土壤温度进行测定,数据每15 min采集1次。
塑料大棚:采用美国Campbellsci公司的CR3000数据采集器和相关传感器,对处理行1.5 m处的气温和0.2 m处的土温进行测定。用温度记录仪(浙江杭州泽大仪器有限公司)对CK行 1.5 m 处的气温和0.2 m处的土温进行测定。
1.5 测定方法
1.5.1 休眠结束期的确定 塑料大棚在揭盖蓄热之后,立即给葡萄枝条芽涂抹生石灰水并进行根域加热处理,将根域加热的日期作为其休眠结束期(表2)。
1.5.2 萌芽率的统计 每个处理选取10株生长良好的葡萄并对其1年生枝条上的芽进行露绿期统计,葡萄开始露绿后每3 d对每个处理的葡萄芽的萌发率进行统计,直至萌发率≥50%,此时即为萌芽期。
萌发率=(露绿期的芽数目)/(总芽数)×100%。
露绿期:从新芽颜色能透过绒毛看到,到嫩芽最外面一片叶子的边缘可见。
1.5.3 需热量的计算 采用6种模型计算需热量值,计算方法如下:(1)温度最大值累计模型[16]:用温度最大值(计作ACT max ℃)表示,ACT max ℃=∑(t日最高温度) ℃;(2)平均温度累计模型[16]:用温度平均值(计作ACT med ℃)表示,ACT med ℃=∑(t日平均温度) ℃;(3)热量模型[17]:用日最高气温与最低气温之 差的累计值表示(计作Heat ℃),Heat ℃=∑(t日最高温度-t日最低温度) ℃;(4)生长度小时模型[18]:用每小时给定的温度(t小时)所相当的热量单位(记作GDH ℃)表示。t小时≤4.5 ℃时,GDH ℃=0 ℃,4.5 ℃< t小时<25.0 ℃时,GDH ℃=t-4.5 ℃,t小时≥25.0 ℃时,GDH ℃=20.5 ℃;(5)有效积温模型[19-20]:有效积温=∑(t日平均温度-t生物学零度),单位为 ℃;(6)最大积温模型[21]:最大积温=∑(t日最高温度-t生物学零度),单位为℃。
2 结果与分析
2.1 2种设施内的空气温度和土壤温度分析
研究发现,在需冷量满足后,气温和土温均呈现上升趋势,且气温比土温上升快,其中气温能够满足枝条芽萌发的温度条件,但是土温却一直维持在8 ℃左右,影响根系的萌动。塑料大棚3年气温平均最高值为15 ℃,最低值为10 ℃,平均值为13 ℃;塑料大棚3年土温平均最高值为11 ℃,最低值为7 ℃,平均值为9 ℃(图1)。
研究发现,在需冷量满足后,气温和土温均呈现稳定的趋势,且气温比土温更高,其中气温和土温均能够满足枝条芽萌发的温度条件。日光温室3年气温平均最高值为25 ℃,最低值为17 ℃,平均值为20 ℃;日光温室3年土温平均最高值为17 ℃,最低值为11 ℃,平均值为15 ℃(图2)。
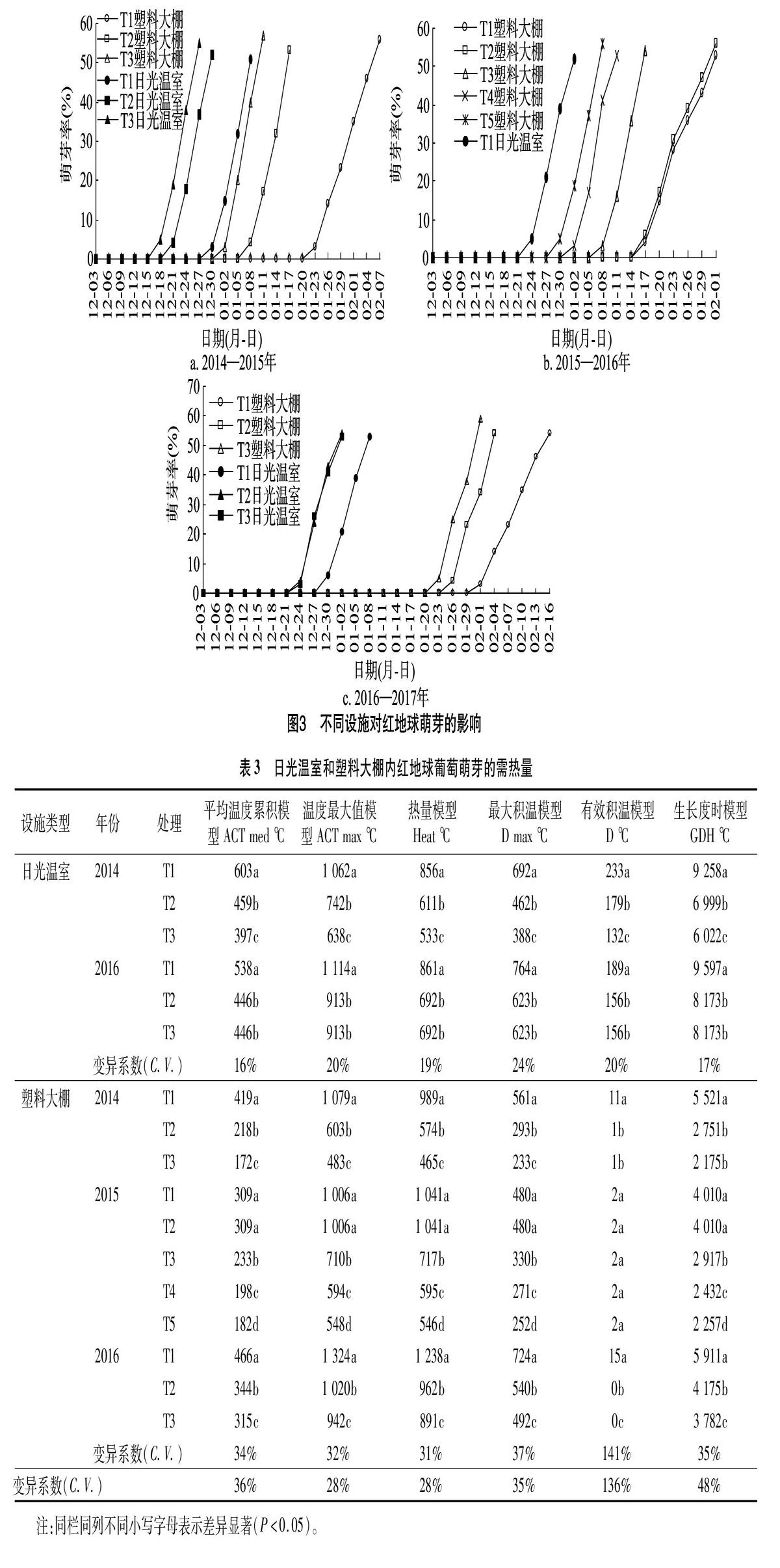
2.2 不同设施红地球枝条芽萌发的差异
研究发现,在2种设施内,不同土壤温度下红地球葡萄枝条芽的萌动时间均表现出差异性,一定的温度范围内,芽萌动时间随土壤温度的提高而提前。土壤温度相同或未加热时,日光温室内红地球枝条芽的萌动时间明显早于塑料大棚内的红地球。不加热情况下,日光温室内红地球葡萄的萌芽进度要比塑料大棚内的快,且2014年、2015年、2016年萌芽50%相差的平均天数为33 d;而当根域温度分别为20、25、27、32 ℃ 时,日光温室内红地球枝条芽萌发50%的日期较塑料大棚分别提前18、15、33、30 d(图3)。
在2种设施内,红地球葡萄的萌芽速度都随着土温的升高而提前,但2015年塑料大棚的CK、15 ℃和2016年日光温室的27、32 ℃没有差异,这表明土温对于红地球葡萄枝条芽萌发的促进是有上下限的;同一设施下,红地球葡萄枝条芽的萌发进度表现出年际差异性,且塑料大棚的年际差异性要比日光温室更大(图3)。
2.3 土壤温度对设施红地球枝条芽需热量的影响
用6种需热量模型计算红地球枝条芽的需热量,并进行多重比较和变异度分析。结果发现,同一设施内,不同根域温
度对于红地球葡萄枝条芽需热量的影响是显著性的,在一定温度范围内,根域温度越高,红地球葡萄枝条萌芽的需热量越小,相较于日光温室,塑料大棚内枝条萌芽需热量的值变异率大,表明土温对塑料大棚内葡萄芽需热量的影响更大;不同设施内,同一土壤温度或未加热条件下,用平均溫度累积模型、有效积温模型、最大积温模型、生长度模型统计的需热量值均表现为日光温室较大,用温度最大值、热量模型统计的需热量值则表现为2种设施内差异性小,这表明设施类型会影响需热量值的大小(表3)。
在2种设施内,用6个需热量模型统计的需热量变异系数的差异较大。在日光温室内,6个需热量模型的变异系数范围为16%~24%,最小为平均温度累积模型,最大为最大积温模型,表明这6种需热量模型均适于日光温室内葡萄需热量的统计;在塑料大棚内,6个需热量模型的变异系数范围为31%~141%,最小为热量模型,最大值为有效积温模型。平均变异系数低于30%的为温度最大值模型和热量模型,这表明温度最大值模型和热量模型适于塑料大棚内葡萄需热量统计。
3 结论与讨论
本研究发现在气温适宜条件下,土壤温度在一定范围内的升高能够明显促进葡萄枝条芽的萌发,显著减少葡萄枝条萌芽的需热量,且在塑料大棚内的效果更明显,这与张福庆等的研究结果[22-24]相符。根系是树体整体发育的基础和中心,通过吸收水分、矿质养分和合成内源激素等途径对叶片生长、碳素同化、花芽分化、果实发育等许多过程产生着影响[25]。张福庆等认为,土温影响葡萄根系的活动,进而影响枝条芽的萌发[22]。De Barba等发现,升高土壤温度会使云杉的萌芽时间提前[24]。孙鲁龙等研究发现,土壤有效积温与葡萄枝条芽的萌发有显著性相关[25]。
同需冷量模型一样,需热量模型也是物候学模型[26],不是以萌芽进程为基础,在不同环境条件下其准确性不同,而不同设施类型便是不同的环境条件,所以需热量模型在不同设施内的准确性具有差异性。本试验中2种设施内红地球葡萄芽的需热量值差异性较大便是这个原因。对于葡萄需热量的研究较少,奚晓军等用生长度时模型统计上海设施内葡萄的需热量,发现其值在9 113~10 722 GDH ℃[8],王海波等用生长度时模型和有效积温模型统计22个葡萄品种的需热量值,发现二者介于9 976~12 541 GDH ℃或253~353D ℃[12],本试验日光温室内红地球葡萄需热量的值接近他们的发现值,但塑料大棚内的与之相差较大。所以在引用需热量值与实际生产应用时,应注意不同设施环境的差异性。
生长度时模型和有效积温模型是最常用的2种模型。但与有效积温模型相比,生长度时模型考虑了低温的无效性、中温的有效性、高温的有限性,更符合自然条件。有效积温模型虽然广泛采用,但在萌芽期间,昼夜温差较大时,会有较大误差。尽管生长度时模型的温度效应划分较细,但在生态型有差异的地区仍会造成误差,但相较于其他模型,生长度时模型仍是最适合实际生产应用的模型。本试验中2种设施内生长度时模型差异性较大的原因是土壤温度和空气温度。
日光温室和塑料大棚的蓄热性相差较大,所以土壤温度和空气温度不同。在生产上,日光温室的葡萄要比塑料大棚的早1个月左右上市,主要就是温度的缘故。温度是葡萄萌芽期最重要的环境因子[27],土壤温度影响地下部的生长发育,空气温度影响地上部生长发育,所以适宜的土温同气温一样重要。本试验也表明,一定范围内土壤温度的升高能够显著性减少葡萄枝条芽的需热量,促进日光温室和塑料大棚内葡萄枝条芽的萌发。
参考文献:
[1]谭 钺,李 玲,李冬梅,等. 设施桃萌芽调控中冷量与热量的关系[J]. 应用与环境生物学报,2012,18(5):728-733.
[2]Alburquerque N,García-Montiel F,Carrillo A,et al. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements[J]. Environmental & Experimental Botany,2008,64(2):162-170.
[3]杨天仪,李世诚,蒋爱丽,等. 葡萄品种需冷量及打破休眠研究[J]. 果树学报,2001,18(6):321-324.
[4]高东升,束怀瑞,李宪利. 几种适宜设施栽培果树需冷量的研究[J]. 园艺学报,2001,28(4):283-289.
[5]章 镇,高志红,盛炳成,等. 葡萄不同品种需冷量研究初报[J]. 中国果树,2002(3):15-17.
[6]王海波,王孝娣,高东升,等. 不同需冷量桃树对周年光温变化的生长和生理响应研究[J]. 西北植物学报,2009,29(10):2058-2062.
[7]王西成,钱亚明,赵密珍,等. 设施葡萄萌芽调控中需冷量和需热量及其相互关系[J]. 植物生理学报,2014,50(3):309-314.
[8]奚晓军,蒋爱丽,田益华,等. 上海地区设施葡萄的需冷量及需热量研究[J]. 上海农业学报,2015(1):23-26.
[9]司海娣. 银川地区主要设施果树促早栽培小气候环境特征研究[D]. 银川:宁夏大学,2011.
[10]王连荣,陈海江,徐继忠,等. 地温对设施桃花芽若干生理指标的影响[J]. 河北农业大学学报,2004,27(1):58-60.
[11]王世平,费全风,秦卫国,等. 根域加温对促成栽培绯红葡萄生长发育的影响[J]. 果树学报,2003,20(3):182-185.
[12]王海波,王孝娣,王宝亮,等. 设施葡萄常用品种的需冷量、需热量及二者关系研究[J]. 果树学报,2011,28(1):37-41.
[13]尹 翠,孙利鑫,董 艳,等. 根区土壤加温对塑料大棚内红地球葡萄生长发育和品质的影响[J]. 浙江农林大学学报,2016,33(6):1092-1097.
[14]尹 翠,董 艳,孙利鑫,等. 根区土壤加温对塑料大棚内葡萄芽萌发及生理特性的影響[J]. 西南农业学报,2016,29(12):2944-2949.
[15]尹 翠,孙利鑫,曹 震,等. 根域加温对塑料大棚内葡萄芽萌发及生理特性的影响[J]. 江苏农业科学,2016,44(8):203-206.
[16]董 艳,尹 翠,孙利鑫,等. 土壤温度对设施红地球葡萄枝条萌芽影响及需热量估算方法评价[J]. 西南农业学报,2017,30(3):669-674.
[17]Galán C,García-Mozo H,Carianos P,et al. The role of temperature in the onset of the Olea europaea,L. pollen season in southwestern Spain[J]. International Journal of Biometeorology,2001,45(1):8-12.
[18]Javier Rodríguez-Rajo F,Frenguelli G,Jato V. The influence of air temperature on the starting date of Quercus pollination in the South of Europe[J]. Grana Palynologica,2003,42(3):145-152.
[19]Anderson J L,Seeley S D. Modelling strategy in pomology:development of the Utah models[C]. Ⅲ International Symposium on Computer Modelling in Fruit Research and Orchard Management.1992:297-306.
[20]Alcalá A R,Barranco D. Prediction of flowering time in olive for the Cordoba olive collection[J]. Hortscience,1992,27(11):1205-1207.
[21]沈元月,郭家选,祝 军,等. 早熟桃品种需冷量和需热量的研究初报[J]. 中国果树,1999(2):39-42.
[22]张福庆,李 巍,田卫东,等. 玫瑰香葡萄品种特性与汉沽产地分析[J]. 中外葡萄与葡萄酒,2007(5):39-42.
[23]De Barba D,Rossi S,Deslauriers A,et al. Effects of soil warming and nitrogen foliar applications on bud burst of black spruce[J]. Trees,2016,30(1):87-97.
[24]孙鲁龙,许丽丽,杜远鹏,等. 有效积温与葡萄萌芽进程的关系[J]. 植物生理学报,2016(8):1263-1270.
[25]束怀瑞. 果树栽培生理学[M]. 北京:农业出版社,1993:162-166.
[26]Hnninen H. Effects of climatic change on trees from cool and temperate regions:an ecophysiological approach to modelling of bud burst phenology[J]. Canadian Journal of Botany,1995,73(2):183-199.
[27]Jones G V,Davis R E. Climate in fluencies on grapevine phonology,grape composition and wine production and quality for Bordeaux,France[J]. American Journal of Enology & Viticulture,2000,51:249-261.张 宇,樊小雪,徐 刚,等. 不同氮肥与有机肥配施对蒜产量及品质的影响[J]. 江苏农业科学,2019,47(5):114-117.
