
东方田鼠家族群对成员个体觅食行为的影响
2019-08-27 07:01:16吴帅玲陶双伦唐显江左方明李俊年
生态学报 2019年13期
吴帅玲,陶双伦,*,章 力,马 静,唐显江,汪 玲,左方明,李俊年
1 吉首大学生物资源与环境科学学院, 吉首 416000 2 莱布尼茨神经生物学研究所, 马格德堡 39118
自然界,动物集群觅食的现象非常普遍[1- 5]。生态学家普遍认为,集群觅食能给群内个体的生存带来诸多好处,例如,互相观察和模仿学习,能增进个体生存和适应环境的能力[4, 6- 7];轮流警惕能提高风险规避效率,缓冲捕食风险压力[8- 9]。新近的研究[10- 11]发现,个体在分享群体觅食带来的上述好处的同时,会因群内成员个体间的相互影响而潜在地增大其觅食代价,例如,信息发现和解读方面的差异性:经验丰富的个体比经验较少的个体能更为敏锐地发现和解读环境信息,同时释放出有价值的信息[12- 14];信息传递的时空差异性:与风险发现者相距较近的个体相比,相距较远的个体因获悉警报信息的时间相对滞后,会延误逃避风险的最佳时机[15- 17];争夺有限的食物资源:个体间常常因争夺有限的食物资源而发生肢体接触式的争斗,或释放虚假警报如频繁抬头、四处张望,甚至采用恐吓、威慑动作驱赶靠近者[18- 20]。
相较于临时性集群及非亲缘关系组成的群体,以亲缘关系为纽带而结成的社会性群体——家族群,成员数一般相对较小,群体结构在一定时期内维持相对稳定,个体于社群所处的等级地位相对明确;个体间熟悉和默契程度较高,易于解读和辨识同伴个体的行为动机及发出的信号信息,相对减少了个体间因相互干扰对觅食造成的不利影响[21- 22]。然而,家族群觅食尽管能给成员个体带来上述诸多好处,但个体间因群体生活而存在的一些不可避免的干扰性因素则能潜在地影响其觅食适合度[10- 11, 23- 26]。例如,群内成员个体每次发出的反捕食警觉信号如警惕性叫声和动作,无论这些警觉行为动作是否能有效地预测风险,均有可能短暂中断或中断其他成员个体的觅食活动,潜在地降低其摄入率[24- 26];而该反捕食警觉信号的准确性和对群内个体的生存价值,又取决于成员个体评价环境时所具有的经验和认知水平[13- 14, 16- 17]。因为它不仅左右着风险发现者对风险信号的判断和决策,还影响着其他成员个体对风险发现者所发出的信号的解读与决策。因此,探讨家族群成员个体间的相互作用对个体觅食行为的影响,有助于揭示家族群成员个体在通过交流与学习等方式分享群体生活所带来的觅食方式及风险规避上的利益的同时,是否存在个体对风险信息处理的差异以及为争夺有限食物资源所产生的干扰性竞争而引起的摄入率降低的代价。我们感兴趣的是,觅食活动中,家族群成员个体发出的各种警觉信号会对群内其他个体的觅食行为产生何种影响?个体的行为反应对自身觅食活动中的各警觉动作时间分配是否会产生影响,进而引起警觉动作时间分配上的变异?个体在警觉时间分配上的变异是否会引起摄入率的动态变化?
为了探明上述问题,以栖息于湖南省洞庭湖区的东方田鼠(Microtusfortis)为实验对象。田鼠亚科(Microtine)啮齿动物具有典型地以母系为核心共占共享同一块区域作为巢区的特征,是研究家族群对成员个体觅食行为影响的理想材料。野外捕捉后,在实验室驯养,人工配对繁育,建立东方田鼠子一代家族群。在实验室条件下,以实验用动物喜食的马唐(Digitariasanguinalis)为食物,构建单一均质的食物斑块。测定东方田鼠家族群在食物斑块觅食时,其成员个体的觅食行为过程、序列及参数;探明成员个体在觅食活动中的各类警觉行为动作、持续时间及参数;检验家族群对群内成员个体各觅食行为参数及摄入率的影响。检验的特定假设为,在可利用食物密集斑块,家族群成员个体的存在能通过影响个体觅食活动中的警觉行为及动作,使其警觉动作时间在分配上发生变异,进而导致摄入率降低。
1 材料与方法
1.1 实验用动物
本研究以栖息于湖南省洞庭湖区的东方田鼠为实验用动物。于2015年6月份捕自岳阳(华容县)、益阳(南县)及常德(汉寿县)三地,三地捕捉点的空间距离为40—80 km。野外捕捉后,带回实验室以干木屑为巢垫,兔颗粒饲料为主要食物,附加一定量新鲜草料,供给充足的饮水,单只驯养于透明饲养笼。控制实验室温度为(20±1)℃,光照周期12 L∶12 D。同年9月,以捕自不同区域的个体进行雌、雄鼠配对。配对鼠于翌年2月开始陆续分娩,分娩后幼子与双亲共同生活,饲养条件同上。2个月后,从中选取窝仔数为3—4只的家族群,幼子年龄2.5—3.0个月,平均体重(53.3±8.1) g,共40个家族群作为实验对象。
1.2 食物斑块配制
采用木质纤维板构建80 cm×30 cm及120 cm×60 cm两种斑块,分别作为单只个体及家族群成员个体的觅食活动空间。采用两种大小不同的斑块构建实验对象的觅食活动空间,目的在于使家族群成员个体在觅食空间及生物量密度配置与单只个体相一致。前期研究[27]表明,随着叶片大小增加,东方田鼠摄入率显著增大,当叶片大小达到6 mg(干重)左右时,摄入率稳定于一定的水平,不再增大。以(6.08±0.10) mg(干重)马唐叶片配置食物斑块,以2.0 cm的空间距离于斑块上钻孔,孔口直径约0.5 cm,以小型塑胶软塞固定叶片于孔口处,构成动物食物密集斑块。
1.3 行为观测装置
为了有效控制实验个体,便于准确地测定个体的觅食行为及过程,在食物斑块上部周边设立观测箱。观测箱由透明玻璃构成,底部面积与食物斑块大小一致,高度为40 cm。观测箱由投放区和觅食区两部分构成(图1)。在投放区周边玻璃上覆以牛皮纸,作为觅食个体规避风险的隐藏区。两区间以底部带孔的隔板相隔,此孔可作为实验个体在两区间自由活动的通路。其中单只个体觅食行为观测箱的投放区面积为40 cm×30 cm,觅食区面积为80 cm×30 cm,用于测定单只个体的觅食行为;家族群成员个体觅食行为观测箱的投放区面积为40 cm×60 cm,觅食区面积为120 cm×60 cm,用于测定家族群成员个体的觅食行为。
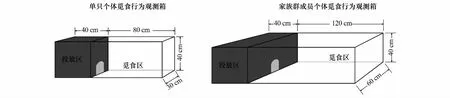
图1 东方田鼠觅食行为观测装置示意图Fig.1 Samples of behavioral observing equipment in reed vole foraging
1.4 行为参数定义
参照Spalinger和Hobbs[28]、Hobbs和Gross等[29]、陶双伦等[27, 30- 31]对植食性哺乳动物觅食及警觉行为参数的定义,测定如下行为参数:采食时间(h):口鼻部接触到食物,至分离食物的时间(min);处理时间(t):咀嚼和吞咽食物的时间(min);采食回合时间(TBB):开始采食至下1次采食活动开始所持续的时间(min),期间短暂中断觅食活动的时间不超过0.5 min;口量(S):通过一次或多次采食动作,经连续咀嚼,1次吞咽的食物量(g);觅食回合时间(TFB):为1次连续采食活动所持续的时间(min),期间至少由2—3个采食回合组成,当采食活动中断至少有1 min,定义该采食活动时间为觅食回合时间;一般扫视时间(Sc):包括平视和抬头扫视时间,其中平视时间(Hsc)为头部转动观察周围环境持续的时间(min);抬头扫视时间(Rsc)为,前肢离地身体向上未完全伸展方式观察周围环境所持续的时间(min);直立扫视时间(Usc):前肢离地,背部与地面几近垂直方式观察环境所持续的时间(min);盯视时间(Star):身体面向特定方向,头部不动,眼睛注视特定个体或物体所持续的时间(min);静听时间(L):停留于原地或站立抬头,一动不动,短暂停止一切活动所持续的时间(min);嗅闻时间(Sn):口鼻部前升,鼻孔部周围肌肉及毛发有明显颤动所持续的时间(min);觅食中断时间(TFI):停止咀嚼,站立抬头向周围扫视、监听及嗅闻等所花费的时间(min);觅食决定时间(TFD):单只个体及家族群成员个体投放于投放区至开始采食第一口食物的时间(min);觅食中断时间比例(TFIP):觅食回合内,个体于采食回合的觅食中断时间占采食回合时间的比例;摄入率(I):单位时间,实验个体摄入食物干物质的量(mg/min)。
1.5 实验程序及觅食行为参数估计
在吉首大学基础生物学实验室测定实验个体的觅食行为。行为测定开始前15日,以新鲜马唐(Digitariasanguinalis)茎秆和叶片为主兔颗粒饲料为辅饲喂实验用动物。每天8: 00—10: 00及15: 00—17: 00分两次将单只个体及单个家族群分别投放至配置有唐草叶片的实验观测箱(图1),使其适应观测装置15min。每天重复2次,重复2周。待其充分适应实验装置后,从5个实验用动物家族群中分别选择2只觅食活跃、毛色光亮的子一代个体,共计10只,作为单只觅食组;从20个实验用动物家族群中选择10个群内成员个体觅食活跃、毛色光亮的家族群(每个家族群的子一代数目为3—4只)作为家族群觅食组。为便于准确记录和辨识家族群各个体的觅食行为过程,采用4种剪毛方式标志家族群子一代个体,标记样式分为:左臀右肩、右臀左肩、左臀左肩及右臀右肩。
行为测定前,饥饿单只实验个体及实验群体4—6 h。测定时,从观测箱投放区投放单只个体或单个家族群,观测者位于观测箱一侧中部,采用焦点取样法,以人工计数及录像机录像测定实验对象上述觅食行为过程、序列及参数,同时,观测者记录焦点个体每口消费的叶片占单个叶片大小的比例[32]。测定结束后,分离、称重每个被食叶片的剩余部分。回播慢放录像,记录和估计焦点个体在觅食活动中的上述觅食行为参数。行为测定结束后,以自来水彻底冲洗斑块,去掉遗留气味,避免影响后续实验。待斑块阴干后,重新配置叶片,重复上述试验。在实验测定期,行为测定均由同一观测者完成。
采用警觉-功能反应模型I=S/(h+t+TFI)[27]计算个体的摄入率。式中参数定义同1.4。
1.6 统计分析
各警觉行为动作时间比例的计算方式为:个体于采食回合的各警觉行为动作时间占其采食回合时间的比例。
家族群因成员个体差异,部分家族群少数个体滞留于投放区,至行为测定结束时都未能进入食物斑块觅食,因此,在统计分析时排除了这部分个体。
实验个体的觅食决定时间、摄入率、觅食中断时间比例及各警觉行为动作时间比例以平均值±标准差(M±SD)表示;采用独立样本t检验分析法,检验单只雌、雄个体、家族群雌、雄成员个体及单只个体与家族群成员个体的觅食决定时间及摄入率的差异显著性;采用Mann-Whitney检验分析法,检验单只雌、雄个体、家族群雌、雄成员个体及单只个体与家族群成员个体的觅食中断时间比例、一般扫视动作(Sc)时间比例、直立扫视动作(Usc)时间比例、盯视动作(Star)时间比例、静听动作(L)时间比例及嗅闻动作(Sn)时间比例的差异显著性。
2 结果
2.1 性别对觅食行为参数的影响
独立样本t检验结果发现,单只雌、雄个体和家族群雌、雄成员个体的觅食决定时间及摄入率均无显著差异;Mann-Whitney检验结果发现,单只雌、雄个体和家族群雌、雄成员个体的觅食中断时间比例及各警觉行为动作时间比例亦均无显著差异(表1,表2)。
上述检验结果表明,不论是单只独立觅食抑或是家族群成员共同觅食,其雌、雄个体的觅食行为参数均无显著差异,因此,本文重点探讨家族群成员个体与单只个体各觅食行为参数的差异。
2.2 家族群对觅食行为参数的影响
2.2.1觅食决定时间
家族群成员个体的觅食决定时间明显低于单只个体,其中,单只个体n=10,TFD=1.317±0.332(min),家族群成员个体n=26,TFD=0.995±0.339(min)。独立样本t检验结果显示,家族群成员个体与单只个体的觅食决定时间差异显著(t(34)=-2.397,P=0.024<0.05)。说明,东方田鼠家族群成员共同觅食能显著地缩短成员个体的觅食决定时间。
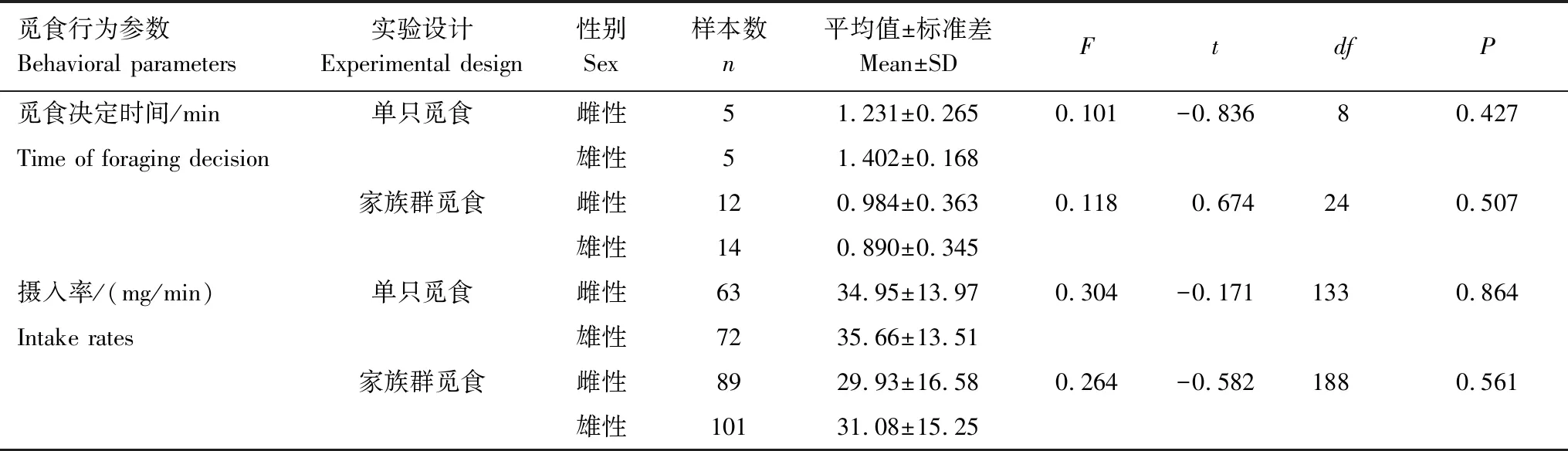
表1 东方田鼠雌雄个体觅食行为参数独立样本t检验分析结果*
*P>0.05,差异不显著

表2 东方田鼠雌雄个体觅食行为参数Mann-Whitney检验分析结果*
*P>0.05,差异不显著
2.2.2摄入率
家族群成员个体的摄入率明显小于单只个体的,其中,单只个体n=135,I=35.37±13.68(mg/min);家族群成员个体n=190,I=30.55±15.64(mg/min)。独立样本t检验结果显示,二者的差异极显著(t(323)=-2.943,P=0.003<0.01)。说明,东方田鼠家族群成员共同觅食能极显著地降低成员个体的摄入率。
2.2.3觅食中断时间比例
家族群成员个体分配于采食回合的觅食中断时间比例明显大于单只个体的,其中,单只个体n=135,TFIP=0.386±0.175,家族群成员个体n=190,TFIP=0.485±0.209。Mann-Whitney检验结果显示,二者的差异极显著(n=190,135;Z=-4.392,P=0.000<0.01)。说明,东方田鼠家族群因成员个体间的相互干扰性影响能极显著地增大个体的觅食中断时间。
2.2.4警觉行为参数

图2 东方田鼠警觉行为动作时间比例 Fig.2 Proportion of time of vigilance behaviour of reed vole foragingSc:一般扫视动作Scanning;Usc:直立扫视动作Upright Scanning;Star:盯视动作Staring;L:静听动作Listening;Sn:嗅闻动作Sniffing
与单只个体相比较,家族群成员个体分配于采食回合的各警觉行为动作时间比例呈现出不同的分配格局(图2)。Mann-Whitney检验结果显示,家族群成员个体的一般扫视(Sc)(n=190,135;Z=-1.687,P=0.039<0.05)、盯视(Star)(n=190,135;Z=-3.672,P=0.000<0.01)及嗅闻(Sn)(n=190,135;Z=-4.580,P=0.000<0.01)动作时间比例均显著或极显著地高于单只个体的;而直立扫视(Usc)(n=190,135;Z=-2.248,P=0.025<0.05)及静听(L)(n=190,135;Z=-3.151,P=0.002<0.01)动作时间比例则均显著或极显著地低于单只个体的。
上述检验结果充分地说明,在可利用食物密集斑块,家族群成员个体的存在能通过影响个体觅食活动中的警觉行为及动作,使其警觉动作时间在分配上发生变异,导致摄入率降低。
3 讨论
本项研究结果发现,在均质密集的马唐叶片食物斑块上,不论是单只独立觅食还是家族群成员共同觅食的环境,东方田鼠个体的性别差异均未对其觅食行为参数——觅食决定时间、摄入率、觅食中断时间比例及各警觉行为动作时间比例造成影响。尽管没有检测到性别对东方田鼠觅食行为的影响,但这并不意味着性别对其没有影响。本项研究没有涉及非均质食物斑块对东方田鼠觅食行为的影响,但自然界植食性哺乳动物的觅食环境复杂多样,食物组成种类丰富、形态及空间结构各异,增大了个体对食物选择和利用的难度[33]。已有的研究[34]发现,哺乳动物对食物的选择和利用存在性别差异,因此,性别可能会潜在地影响个体的觅食行为,因此,有必要在空间异质性食物斑块条件下就性别对动物觅食行为的影响开展研究。
东方田鼠家族群影响成员个体的觅食决定时间和摄入率。相较于单只个体的独立觅食活动,东方田鼠家族群觅食尽管能明显缩短成员个体的觅食决定时间,但却使成员个体的摄入率显著地降低。如前文述及,群体觅食尽管能给群内成员个体在规避风险、提高觅食效率等方面带来诸多好处[8- 11],但这些好处却是以降低成员个体的摄入率为代价的。
相较于单只个体,家族群成员个体之所以能缩短觅食决定时间快速地进入觅食期,与群体的风险稀释效应[8, 35]有关,即在相同风险压力条件下,家族群成员个体所面对的风险压力由群内所有成员个体共同承担而得到缓解和稀释,而单只个体则须独自承担风险压力,因此,相较而言,家族群每一个体平均承担的风险压力要远小于单只个体。觅食决定时间反映了特定环境条件下个体对收益—风险的权衡。
东方田鼠家族群成员个体摄入率降低与个体觅食时因警觉而引起的个体间相互干扰密切关联。植食性哺乳动物的觅食行为过程及序列典型地由搜寻与采食和处理(咀嚼和吞咽)食物动作组成,期间伴随各种警觉行为动作如一般扫视、直立扫视、静听和嗅闻[27, 30- 31, 36]。群体觅食时,成员个体除了发生上述警觉行为动作,同时还因个体间的相互干扰性动作如警报性鸣叫、警惕性姿态、最小空间距离保持、对同一个食物项目的争夺等[3, 11, 19, 37-39],会使觅食活动中的警觉行为动作—盯视动作从单只个体独立觅食时很不明显到群体觅食时变得非常明显。根据植食性哺乳动物警觉-功能反应理论[27],在食物密集斑块,个体的摄入率不仅受采食和处理食物调控的制约,而且还受因警觉而引起的觅食活动短暂中断的影响。与单只个体的觅食中断时间相比较,家族群成员个体的觅食中断时间显著增大。本项研究配置的食物斑块无论食物大小、食物密度及生物量密度于单只个体和家族群成员个体均为一致,依据警觉-功能反应模型理论,决定摄入率大小的变量—采食时间(h)与处理时间(t)在两种觅食条件下基本维持稳定,然而觅食中断时间却呈现显著地差异,因此,我们认为,植食性小型哺乳动物这种单只个体独立觅食行为特征与群体成员个体的迥异之处,是导致家族群成员个体觅食中断时间增大和摄入率降低的直接动因。
检测东方田鼠觅食行为特征及引起觅食活动短暂中断的警觉行为动作参数动态变化,发现家族群成员个体的一般扫视、盯视及嗅闻动作时间比例均显著地大于单只个体的,而直立扫视和静听动作时间比例则显著地小于单只个体的(图2)。究其原因,如上文所述,这与家族群成员个体在觅食活动中因同伴在场而受到来自同伴的各种警觉行为动作的干扰影响密切相关[11, 18- 19, 24- 26, 36]。相较于单只个体,家族群成员个体因同伴在场,会以一般扫视和盯视动作观察同伴及周围环境即可收集和评价环境信息,因而增大了一般扫视和盯视动作时间比例,相应地减少了直立扫视动作时间比例。
植食性小型哺乳动物体型小,视力极易受到环境的制约,为补偿眼睛收集信息的不足,在长期进化过程中发展出了极为灵敏的听觉和嗅觉器官[40-43]。但家族群成员个体间的相互干扰,增大了个体辨识和收集外界环境信息的难度,使得静听动作时间比例相应地减少。家族群成员个体嗅闻动作时间比例增加与成员个体排泄的粪便尿液以及身体散发出的不同化学分子气味增大了嗅闻监测环境信息的难度密切相关。然而,令人困惑的是,家族群成员共同觅食时,成员个体会发出各种警觉声音,这些声音非但没能增大个体的静听动作时间比例,反而会使其减小?相反,群体发出的各种气味则能使成员个体的嗅闻动作时间比例增大?究其原因,我们认为主要在于听觉和嗅觉动作所检测的环境风险性质不同[30- 31],前者与视觉动作的功能一致,目的在于检测潜在的紧迫性风险,能通过视觉动作加以弥补;而后者检测的则主要为潜在的非紧迫性风险,因其监视的风险具有非直接性、非突发性、非紧迫性及潜在性等特征,不能为视觉、听觉等其他警觉行为动作所替代,致使嗅闻动作时间比例增大。因此,无论是单只个体抑或是家族群成员个体,觅食活动中采用嗅闻动作监视环境风险所花费的时间比例均远高于其他警觉行为动作的。
本文定义的觅食决定时间与已有的研究有所不同。已有的研究所定义的觅食决定时间是斑块模型边际值理论[44]的重要概念,它阐释了植食性哺乳动物在食物呈斑块状分布的环境条件下需通过以口量决定的摄入率来选择食物斑块。当动物在特定食物斑块的摄入率大于在环境中的平均摄入率时,会滞留于该斑块进行觅食活动,否则会放弃该斑块,选择进入新的其他的食物斑块。而本文定义的觅食决定时间是确保实验用动物在维持最大摄入率(实验配置了东方田鼠采食唐草叶片所需的最大口量)的前提下,从其投放于投放区至其在食物斑块上采食第一口食物时所花费的时间。行为测定发现,家族群成员个体与单只个体从投放区进入觅食区的觅食行为明显不同。植食性哺乳动物的觅食活动由众多个觅食回合所组成[27]。家族群成员个体投放于投放区,在投放区滞留0.5—1.0 min后,会在其中一个体带领下,逐次进入食物斑块进行觅食活动,直至进行多个觅食回合方先后返回投放区;而单只个体在投放区滞留1.0—3.0 min后方进入食物斑块,并沿着斑块周边巡视一周后才进行觅食活动,期间以静听、扫视和边走边嗅闻等警觉行为动作收集环境信息,至少需要进行0.5—1.0 min或更长时间的警惕后,才进入觅食期。由于家族群成员个体与单只个体在觅食行为及过程上的巨大差异,使得经典的斑块模型边际值理论[44],针对单只个体的觅食活动而定义的觅食决定时间,不能反映群体觅食在满足个体摄入率条件下对食物斑块的行为选择过程,而新定义的觅食决定时间概念则能很好地区分和反映二者的相异之处。
综上所述,东方田鼠家族群成员个体因觅食活动中的警觉行为动作引起的相互干扰,引致警觉行为动作发生变异,使得觅食中断时间比例增加,增大了收获每口食物的时间,致使其摄入率减小。说明,东方田鼠以家族群进行觅食活动尽管能在稀释及回避风险上带来某些好处,但这些好处是以降低个体摄入率为代价的。因此,我们推测,尽管家族群成员个体的摄入率降低,但因在稀释及回避风险上的好处,群体成员会采用延长觅食时间维持其日食物摄入量的策略,满足其日能量需求[32]。植食性哺乳动物的觅食行为作为其生命活动的重要组成部分,其觅食行为特征及策略的进化适应性意义越来越受到人们的关注[39, 45-46],开展此类研究不但能揭示动物觅食行为的各种适应性策略,而且亦为长江以南地区,尤其是洞庭湖周围地区的东方田鼠的管理等提供理论支撑。
猜你喜欢
文学教育(2023年7期)2023-08-10 17:48:40
辽河(2022年1期)2022-02-14 05:15:04
作文大王·低年级(2021年11期)2021-12-22 00:58:13
小天使·一年级语数英综合(2021年2期)2021-05-08 07:48:34
小学生(看图说画)(2020年4期)2020-04-01 11:58:18
小哥白尼(军事科学)(2019年9期)2019-12-21 02:09:34
电影(2019年3期)2019-04-04 11:57:18
外语学刊(2018年6期)2018-06-19 08:39:42
阅读(低年级)(2018年11期)2018-05-14 09:37:53
少儿科学周刊·少年版(2017年3期)2017-06-29 14:01:15
