
基于代谢组学的多西他赛调控肺癌细胞代谢重编程研究
2019-08-26 06:08孙海超朴海龙齐欢颜敏刘宏旭
中国肺癌杂志 2019年4期
孙海超 朴海龙 齐欢 颜敏 刘宏旭
肺癌不仅是中国发病率和死亡率最高的恶性肿瘤,也是全世界范围内病死率最高的恶性肿瘤[1],其中非小细胞肺癌(non-small cell lung cancer, NSCLC)患者约占肺癌患者总数的80%[2]。40%-50%的NSCLC患者在确诊时已经处于局部晚期或发生转移,5年生存率仅为5%。多西他赛(Docetaxel,多西紫杉醇)属于紫杉醇类,是一种微管稳定剂[3],通过稳定微管蛋白质,抑制微管解聚和肿瘤细胞有丝分裂,发挥其抗肿瘤作用,临床上主要用于治疗NSCLC、乳腺癌、前列腺癌和宫颈癌等[4]。目前,多西他赛是治疗局部晚期或发生转移的NSCLC患者的一线用药[5],其对顺铂治疗产生耐药性的患者也有效[6]。但是,多西他赛治疗NSCLC的分子机制尚未完全阐明。
代谢物变化是对细胞所进行生理活动的最直观反应,代谢物对研究细胞生理和病理机制具有重要意义,目前已鉴定出的生物体内代谢物大于4万个[7]。代谢组学是一种新兴的研究方法,是对整个生物体系总的代谢物所进行的全面定性定量研究[8]。其和基因组学、转录组学、蛋白质组学一起,成为系统生物学的重要研究方法。色谱-质谱联用以其高分辨力、高通量和高灵敏度的优势,已成为代谢组学的主流研究技术之一。基于气相色谱-质谱联用(gas chromatography-mass spectrometer,GC-MS)技术的代谢组学方法在检测糖酵解、三羧酸(tricarboxylic acid, TCA)循环、氨基酸和有机酸等代谢物中具有优势。并且,相对于其他代谢组学技术,GCMS技术较成熟、仪器更加稳定、价格便宜[9]。
1922年瓦博格首次发现,肿瘤的发生发展与糖酵解代谢异常密切相关。在氧气充足的条件下,细胞大量摄取葡萄糖生成乳酸,激活相关信号通路和改变肿瘤微环境。相关研究表明,许多调控代谢的相关蛋白质可以调控细胞凋亡的发生,同时某些调控细胞凋亡的蛋白质也在代谢通路中发挥信使作用。靶向肿瘤代谢治疗的研究是基于代谢组学的方法,通过分析肿瘤异常代谢物和代谢通路,使用现有的生物化学技术合成新的靶向代谢的药物,为治疗肿瘤寻找新的突破点,为预防肿瘤寻找新的策略[10]。靶向肿瘤代谢的基本研究方法包括:①靶向代谢突变基因和激活的代谢基因;②回补肿瘤内缺失的代谢基因;③靶向代谢通路重编程等。
1 材料与方法
1.1 仪器与试剂 使用GCMS-QP 2010Plus系统进行GC-MS代谢组学分析(日本岛津公司)。自动进样器:AOC-20i autosampler(日本岛津公司)。观察细胞使用倒置显微镜(德国Leica公司)。Western blot分析使用Fusion Fx化学发光仪(法国VILBER公司)。CO2培养箱、96孔板、细胞培养皿等均购自美国Thermo Fisher Scientific公司。
超纯水来自于Milli-Q水纯化系统(美国Millipore公司)。40 mg/mL多西他赛(溶于吐温80溶剂)购于齐鲁制药有限公司。甲氧胺盐酸盐、吡啶、二氯甲烷及N-甲基-N-(三甲基硅基)三氟乙酰胺(MSTFA)和十三酸均购自美国Sigma-Aldrich公司。色谱纯甲醇购自德国Merck公司。RPMI-1640培养基、胎牛血清(fetal bovine serum,FBS)、青霉素链霉素液(penicillin-streptomycin, PS)、磷酸缓冲盐溶液(phosphate buffer saline, PBS)、通用细胞冻存液和0.25%EDTA胰蛋白酶均购自美国Gibco公司。
人源NSCLC细胞A549和H1299购自中国科学院上海细胞库,并使用含10%FBS及1%PS的RPMI-1640培养基进行培养。细胞培养在5%CO2、37 ℃恒温孵箱中。细胞培养至密度为80%-90%时进行传代。
1.2 CCK-8测定细胞活力 活细胞内线粒体脱氢酶可与细胞增殖及毒性检测试剂(cell counting kit-8, CCK-8)反应变橙色。使用96孔板培养细胞,消化处于对数生长期的细胞,96孔板每孔接种3,000-5,000个细胞,24 h后A549和H1299细胞按照梯度0.3 nM、1 nM、3 nM、10 nM、30 nM、100 nM、300 nM、1,000 nM处理多西他赛,平行设置三个对照组。继续培养24 h、48 h、72 h后,吸取上清后每孔按照1:9(CCK-8:培养基)加入CCK-8试剂100 μL,培养2 h后使用酶标仪(450 nm波长)测取并计算平均吸光度值A,细胞活力(%)=(A处理组-A空白组)/(A对照组-A空白组)。
1.3 蛋白质免疫印迹分析 分别取对数生长期的A549和H1299细胞2×106个接种于4个10 cm2细胞培养皿。24 h后分别在培养基中加入30 nM和100 nM的多西他赛,处理0 h、6 h、12 h和24 h。弃去培养基,PBS清洗3次后。加入RIPA裂解液,刮下细胞,并将含细胞碎片的裂解液移至新离心管中。冰上裂解细胞1 h,每10 min涡旋一次。4 ℃,13,000g离心15 min,取上清。通过BCA法测定蛋白质浓度,等量蛋白质加入蛋白质上样缓冲液后,97 ℃变性10 min。通过SDS-聚丙烯酰胺凝胶电泳分离不同分子量大小的蛋白质,再将蛋白质转移至PVDF膜上。室温封闭1 h,4 ℃孵育一抗过夜。室温孵育二抗1 h,加入显影剂,化学发光成像仪检测蛋白质条带。本研究所用一抗:Vinculin,购自美国Sigma-Aldrich公司,1:1,000稀释使用;PARP、IDH1和IDH2均购自美国Proteintech公司,1:1000稀释使用。
1.4 样品提取及衍生 分别取对数生长期的A549和H1299细胞2×106个接种于10个10 cm2细胞培养皿。24 h后,5组作为处理组分别加入药物多西他赛30 nM(A549)或100 nM(H1299),5组作为对照组仅换液。药物处理24 h后,弃去培养基,使用5 mL冷PBS缓冲液轻柔冲洗细胞两次,立即液氮淬灭。加入含10 μg/mL十三酸的甲醇:水(4:1)1 mL,刮下细胞至(Eppendorf, EP)管中。涡旋两次,每次1 min。4 ℃,13,000g离心15 min,取上清,真空干燥冻干机(美国Labconco公司)中冻干[11]。
取出冻干样品,每个样品加50 μL甲氧吡啶(20 mg/mL),涡旋60 s,37 ℃水浴肟化1.5 h。再加入40 μL MSTFA,涡旋30 s,37 ℃水浴,硅烷化1 h。4 ℃,13,000g离心15 min。弃去不溶颗粒沉淀,取上清行GC-MS分析[12]。
1.5 GC-MS条件 色谱条件:使用DB-5 MS(30 μm×250 μm×0.25 μm;美国J&W Scientific公司)毛细管柱;使用程序性升温进行分析,起始温度为70 ℃,保持3 min后,以5 ℃/min速率升高温度至300 ℃,保持10 min;接口温度230 ℃;电离模式70 eV;进样量1 μL;载气线性速度40.0 cm/s,恒流模式流速1.19 mL/min;采用全扫描模式;扫描范围33 m/z-600 m/z。
针对当前农村小学生口语交际信心不足的现状,教师应当在日常的教学活动中,加强和学生之间的沟通交流,树立学生口语交际的自信心。首先,构建和谐互动的课堂教学氛围,在潜移默化中培养学生善于交际的能力。农村小学语文教师要改变过去“一言堂”的教育模式,在课堂教学中积极和学生之间交流互动,形成融洽的交际氛围,消除学生的心理障碍和畏难情绪;其次,针对个别自信心尤其不足的孩子,农村小学语文教师要积极沟通,了解原因,积极鼓励,帮助学生树立信心,为培养学生的口语交际和表达能力奠定基础。
1.6 数据处理与统计分析方法 GC-MS所获取的原始质谱数据(CDF格式)通过ChromaTOF 4.43软件(美国LECO公司)进行处理,结合数据库及组内标样,进行代谢物定性。通过GC-MS solution软件进行代谢物积分定量。得到代谢物峰表后,使用SIMCA-P 11.0软件(瑞典Umetrics公司)进行偏最小二乘法判别分析,绘制PLS-DA得分图,可视化组间聚集及离散程度。运用统计学t检验方法(P<0.05),得到对照组和多西他赛处理组的A549和H1299细胞差异代谢物。使用MeV热图软件绘制差异代谢物热图,使用MetaboAnalyst 3.0在线软件进行通路富集分析,使用GraphPad Prism 5绘图软件制作柱状图。
2 结果
2.1 多西他赛对NSCLC细胞A549和H1299的抑制作用 为探讨多西他赛对NSCLC增殖的影响,本实验采用CCK-8试剂盒测定了多西他赛对A549和H1299细胞活力的作用。图1A-图1B为多西他赛8个浓度梯度和3个作用时间对A549和H1299细胞活力的影响。随着多西他赛药物浓度升高和作用时间延长,A549和H1299细胞活力下降,多西他赛对NSCLC的增殖抑制作用具有明显的浓度和时间依赖性。
为探讨多西他赛对NSCLC凋亡的作用。通过Western blot实验,测定多西他赛作用A549和H1299细胞0 h、6 h、12 h和24 h时,凋亡敏感指标PARP蛋白质表达。结果表明,随着多西他赛处理时间延长,A549和H1299细胞中的PARP蛋白质均逐渐被激活裂解形成P89片段(图1C),细胞发生凋亡。上述结果提示多西他赛能诱导NSCLC发生凋亡。
2.2 GC-MS代谢组学技术分析多西他赛处理前后的A549和H1299细胞差异代谢物 基于代谢组学中需要活细胞提取代谢物进行分析的基础上,综合考虑多西他赛对NSCLC增殖和凋亡的影响,保证多西他赛的药理作用,同时保持相当程度的细胞活力。根据CCK-8实验结果,选择作用A549细胞的药物浓度为30 nM,作用H1299细胞的药物浓度为100 nM,作用时间为24 h,A549和H1299细胞活力均在70%-80%之间,实验具有可行性。
提取代谢物前使用液氮淬灭细胞,去除微环境变化对细胞代谢状态的影响[14]。依前述方法样品经提取硅烷化衍生,利用GC-MS技术分析细胞代谢物。得到对照组和多西他赛处理组A549和H1299细胞代谢物的GC-MS总离子流图。
考虑到非药物因素的影响,排除复杂的环境条件干扰,最大化组间分离,并寻找各组间的差异代谢物。进一步通过偏最小二乘法判别分析(partial least square discriminant analysis, PLS-DA)经行多元数据处理。图2是对照组和处理组A549和H1299细胞代谢物全谱数据的PLS-DA得分图,该模型包含两个主成分[14],其中R2X=0.831、R2Y=0.983和Q2=0.878,提示该模型具有较高的稳定性和较好的预测率。在PLS-DA得分图上,对照组和处理组A549和H1299细胞代谢物可以明显的区分开。
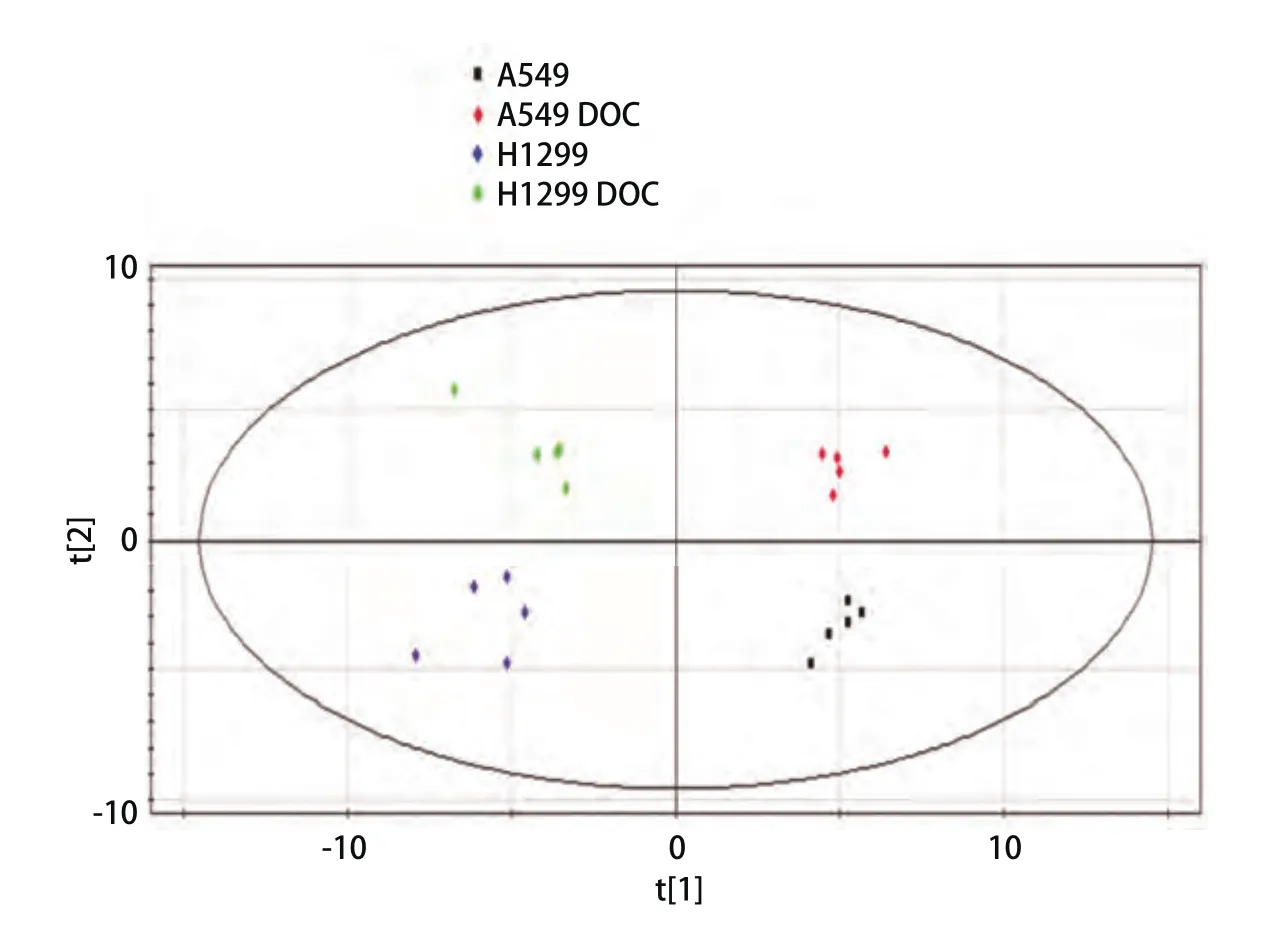
图2 对照组和多西他赛处理24 h的A549和H1299细胞代谢物的PLS-DA得分图(n=5)Fig 2 PLS-DA score plot of control and docetaxel treated A549 and H1299 cells for 24 h (n=5)
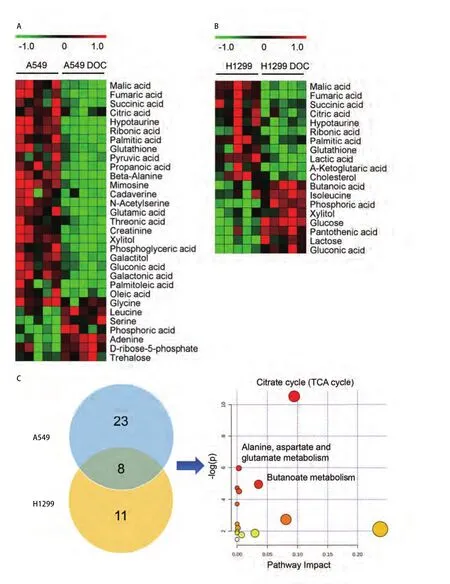
图3 多西他赛对A549和H1299细胞中代谢物和代谢通路的影响(n=5)。A-B:对照组和多西他赛处理24 h组的A549细胞(A)和H1299细胞(B)差异代谢物的热图;C:韦恩图统计A549和H1299细胞中共同的差异代谢物,并对8种差异代谢物进行通路富集分析。Fig 3 Effects of docetaxel on metabolites and metabolic pathways in A549 and H1299 cells (n=5).A-B: Heatmap of metabolites in A549 cells (A)and H1299 cells (B) following docetaxel treatment for 24 h; C: Venn diagram of commonly differential metabolites in both A549 and H1299 cells.Pathway Impact analysis results by 8 differential metabolites.
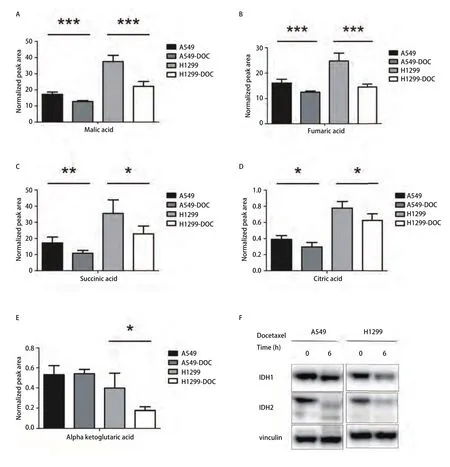
图4 多西他赛对A549和H1299细胞的TCA循环的影响。A-E:对照组和多西他赛处理24 h的A549和H1299细胞中TCA循环中间产物:苹果酸、富马酸、琥珀酸、柠檬酸、α-酮戊二酸变化情况;F:对照组和多西他赛处理6 h的A549和H1299细胞的TCA循环关键酶IDH1和IDH2蛋白质的表达。n=5,*P<0.05,**P<0.01及*** P<0.001。Vinculin作为内参蛋白质。Fig 4 Effects of docetaxel on TCA cycle in A549 and H1299 cells.A-E: Alterations of intermediate metabolites of TCA cycle, included malic acid,fumaric acid, succinic acid, citric acid, α-ketoglutaric acid, in control and docetaxel treatment group in A549 and H1299 cells for 24 h; F: The expression of the key enzymes protein of TCA cycle, included isocitrate dehydrogenase 1 and isocitrate dehydrogenase 2, in control and docetaxel treatment group in A549 and H1299 cells for 6 h.n=5, *P<0.05, ** P<0.01, *** P<0.001 vs the control group.Vinculin was used as a loading control.
结合t检验的统计方法,得到多西他赛处理前后的A549差异代谢物31种和H1299细胞差异代谢物19种,MeV热图分析提示处理组的大部分代谢物含量下降 (图3A-图3B)。处理组A549细胞的丙酮酸、丙酸、甘氨酸等25种代谢物含量下降,丝氨酸、磷酸、腺嘌呤等6种代谢物含量上升。处理组H1299细胞的乳酸、琥珀酸、胆固醇等11种代谢物含量下降,乳糖、木糖醇、葡萄糖等8种代谢物含量上升。
2.3 多西他赛可下调NSCLC的TCA循环代谢途径 多西他赛处理组A549和H1299的共同差异代谢物包括:苹果酸(malic acid)、富马酸(fumaric acid)、琥珀酸(succinic acid)、柠檬酸(citric acid)、亚牛磺酸(hypotaurine)、核糖酸(ribonic acid)、棕榈酸(palmitic acid)和谷胱甘肽(glutathione),在药物处理后含量下降。通过使用在线工具MetaboAnalyst 3.0软件,进行8种共同差异代谢物的通路富集分析(图3C),结果显示主要影响TCA循环代谢途径(P=0.002,14)。
图4A-图4E所示为多西他赛处理后含量下降的TCA循环中间产物。苹果酸、富马酸、琥珀酸、柠檬酸在处理组的A549细胞中含量下降。苹果酸、富马酸、琥珀酸、柠檬酸、α-酮戊二酸(α-ketoglutaric acid)在处理组的H1299细胞中含量下降。
异柠檬酸脱氢酶(isocitrate dehydrogenase, IDH)是TCA循环过程中的关键酶,可催化异柠檬酸转化为α-酮戊二酸。对对照组和多西他赛处理6 h后的A549和H1299细胞进行Western blot分析,药物处理6 h后IDH1和IDH2蛋白质的表达量下降(图4F)。推测多西他赛通过抑制TCA循环关键酶,下调NSCLC细胞的TCA循环。
3 讨论
本研究通过CCK-8实验、Western blot分析,使用基于GC-MS技术的代谢组学方法,结合多种统计软件,探索多西他赛对A549和H1299细胞代谢的影响。结果表明,多西他赛对NSCLC的主要作用有:①可浓度和时间依赖地抑制细胞活力和诱导细胞凋亡;②下调TCA循环的中间代谢物:苹果酸、富马酸、琥珀酸和柠檬酸含量,同时也下调亚牛磺酸、核糖酸、棕榈酸、谷胱甘肽的含量;③下调TCA循环的关键酶异柠檬酸脱氢酶蛋白质水平。
TCA循环是产生ATP的一条重要电子传递链,主要在生物体细胞的线粒体内进行[15],是生物能量的重要来源,同时也为肿瘤细胞增殖提供了必要条件。TCA循环与NSCLC的发生发展密切相关[16]。TCA循环中的关键酶:琥珀酸脱氢酶(succinate dehydrogenase)、异柠檬酸脱氢酶和延胡索酸水合酶(fumarate hydratase)突变表达的非小细胞肺癌患者有着更高的复发率和死亡率[17]。研究[18]表明,通过分析注射同位素13C标记葡萄糖肺癌患者的肺组织,相比正常肺组织,肺癌组织内大量的13C富集在琥珀酸和柠檬酸中。大量葡萄糖经过TCA循环转化为脂质、蛋白质和核酸以满足肺癌组织生长的高合成代谢需求。
靶向代谢重编程药物的研究,主要是针对肿瘤细胞内存在的异常代谢。瓦博格提出,在氧气绝对充足的条件下,相比于正常组织,肿瘤组织的细胞内氧化磷酸化被抑制[19],糖酵解水平升高200倍以上(瓦博格效应)。糖酵解通过转化为谷氨酰胺合成α-酮戊二酸,参与TCA循环,促进肿瘤的进一步发展[20]。添加外源柠檬酸,负反馈调节抑制肿瘤细胞的TCA循环可抑制A549细胞的增殖和裸鼠异体移植瘤的生长[21],研究结果表明对代谢重编程的调控可以治疗NSCLC。
代谢组学是研究生物体系内各种生理活动调节后最终状态的学科,为本研究提供了重要的理论基础。GCMS气相色谱质谱联用技术,为本研究提供了技术上的可行性。通过对差异代谢物的代谢组学分析,发现多西他赛作用于NSCLC细胞后TCA循环等代谢通路的改变,证明了多西他赛对代谢通路调控作用。后续实验需进一步验证多西他赛作用于TCA循环的机制,三羧酸循环影响NSCLC的机制和多西他赛对代谢调控作用的动物实验。希望可通过本研究,为靶向代谢重编程药物的开发和药理学研究提供一种新的策略。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
中山大学学报(自然科学版)(中英文)(2022年4期)2022-08-05
江西水产科技(2022年2期)2022-05-17
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2021年4期)2021-07-23
现代仪器与医疗(2021年1期)2021-06-09
智慧健康(2021年33期)2021-03-16
天津医科大学学报(2021年1期)2021-01-26
农药科学与管理(2019年5期)2019-08-13
食品界(2018年8期)2018-09-03
