
茶树根系跨膜吸收氟的生理与分子机制
2019-08-21 01:11徐佳佳广敏史书林郜红建
茶叶科学 2019年4期
徐佳佳,广敏,史书林,郜红建*
茶树根系跨膜吸收氟的生理与分子机制
徐佳佳1,2,广敏2,史书林2,郜红建1,2*
1. 茶树生物学与资源利用省部共建国家重点实验室,安徽农业大学茶与食品科技学院,安徽 合肥 230036;2. 农田生态保育与污染防控安徽省重点实验室,安徽农业大学资源与环境学院,安徽 合肥 230036
茶树[(L.) O. Kuntze]是高富集氟的植物,氟在叶片中被大量累积。饮茶是人们摄取氟的重要途径,氟的过量摄入会影响人体健康。茶树主要通过根系从土壤中吸收富集氟,但是根系跨膜吸收氟的生理与分子机制尚不清楚。本文综述了茶树根系吸收氟的主动和被动途径,总结根系H+-ATPase和Ca2+-ATPase介导氟的跨膜主动吸收过程与分子机制;剖析离子通道和Al-F络合在根系被动吸收氟过程中的作用及微观过程;分析影响根系吸收富集氟的主要因素及其调控措施。提出通过研究茶树根系氟跨膜吸收相关转运蛋白及其相关基因的克隆、表达和功能验证,以揭示跨膜吸收氟的分子机制;进而研究调控根系对氟的选择吸收,以保障茶叶质量安全和饮茶健康。
茶树根系;氟;跨膜吸收;生理与分子机制;影响因素
氟对人体骨骼和牙齿的生长发育至关重要[1],适量的氟可以促进骨骼和牙齿的钙化,增强骨骼强度,但摄入过多会引起氟斑牙、氟骨病等氟中毒症状[2]。氟虽然不是植物的必需元素[3],但是,因茶树具有高量吸收、转运和积累氟的特殊机制和能力,可被茶树大量吸收[4]。叶片中氟的积累量占茶树全株的98%,尤其是成熟叶和老叶,其含量分别达到500 mg·kg-1和2 000 mg·kg-1以上[5-6]。在没有大气氟污染的情况下,根系是茶树吸收、转运氟的主要器官[7],因此,减少根系对氟的吸收,是降低叶片累积氟的重要途径。已有的研究表明,茶树根系可以通过主动和被动两种途径吸收富集氟[8-9],其中包括H+-ATPase[10]/Ca2+-ATPase[11]参与的跨膜吸收,Al-F络合吸收[12-13],离子通道[4]参与吸收等。这些吸收过程与根细胞膜上的转运/通道蛋白密切相关,但其生理与分子作用机制尚不明确。本文着重阐述转运/通道蛋白介导的茶树根系吸收氟的主动和被动过程及生理与分子机制,为调控茶树对氟的吸收提供理论依据。
1 茶树根系吸收的氟形态
茶树能从土壤中吸收并在体内累积氟[14],土壤中氟的形态及其有效性是影响茶树根系吸收氟的主要因素[15]。土壤氟主要以难溶态、交换态和水溶态等形态存在[7],难溶态和可交换态氟很难被茶树吸收,水溶态是茶树吸收富集的主要氟形态[16],且是土壤溶液中活性最高的氟形态,并以F-、HF2-、H2F3-、H3F4-、AlF63-、FeF63-等离子或络合物形式存在[17]。研究表明,Al-F络合物比F-更容易被茶树根系吸收和转运到新梢[6,18],土壤中的Al-F络合物主要以AlF2+、AlF2+、AlF3形式等被根系吸收,并以AlF2+、AlF2+、AlF4-等形态被木质部运输[13,19]。Ruan等[19]认为,F-也易被茶树根系吸收,最终大部分被运输到叶片,并以F-形式被积累[20]。张显晨等[21]发现,低浓度Al3+(1.05~5.26 mmol·L-1)条件下,氟主要以F-形态被茶树根系吸收,并促进氟在根系的富集;高浓度Al3+(10.52~31.57 mmol·L-1)条件下,氟主要以Al-F络合态被吸收,并抑制了氟的富集。
2 茶树根系吸收氟的机制
2.1 外界氟浓度影响氟吸收途径
外界氟浓度决定了其进入根细胞的途径。当外界低氟质量浓度(0.1~10 mg·L-1),茶树根系主动吸收氟,吸氟过程不仅符合Michaelis-Menten动力学模型,而且依赖能量,高氟质量浓度范围内(50~100 mg·L-1),根系被动吸收氟,并随着氟浓度升高呈线性增长[4,9]。彭传燚等[9]认为,主动吸收是茶树根系摄取氟的主要途径。
2.2 茶树根系对氟的主动吸收
2.2.1 H+-ATPase介导氟的跨膜主动吸收
茶树根系吸收氟是一个与跨质膜电化学势梯度、代谢能量和载体蛋白密切相关的主动运输过程[10]。离子的跨膜吸收和主动运输主要依赖质膜H+-ATPase水解ATP在细胞膜内外两侧建立起来的ΔH+(H+浓度梯度)和电化学势[22],ΔH+的增加,可以刺激植物吸收营养物质[23]。排出的H+导致质子梯度并产生Δp(质子驱动力),激发根细胞质膜中的质子偶联转运蛋白吸收阴离子,如NO3-(NRT1和NRT2转运蛋白),PO43-(PHT1转运蛋白),SO42-(SULTR1转运蛋白),Cl-等[23-25]。被激活的质膜H+-ATPase会增强膜外的电化学势,驱使离子通过次级运输系统进入细胞以提高植物对养分的吸收[26]。Zhang等[27]认为,Al3+提高茶树根系H+-ATPase活性,增加H+的跨膜外排,进而促进了茶树对氟的吸收。有研究表明,H+-ATPase的水解能增加活性H+转运[28]。而如果H+-ATPase的水解受到抑制,会减少H+的产生,主动吸收所需的ΔH+合成受阻,从而降低根系质膜内外的Δp,使茶树根部对氟的吸收和累积不断减少[10]。因此,H+-ATPase水解和活性的增加是茶树根系对F-的跨膜主动吸收的驱动力。
2.2.2 Ca2+-ATPase介导氟的跨膜主动吸收
茶树对氟的吸收与质膜Ca2+泵和钙转运ATPase有关,尤其是Ca2+-ATPase(ACAs),同类型ACA9可能参与茶树氟吸收或氟胁迫的响应[11]。Ca2+-ATPase催化质膜内侧ATP水解,释放能量,驱动Ca2+泵出细胞质。质膜H+-ATPase产生的H+梯度,也可以促进Ca2+泵活动,植物体内Ca2+信号的传导可调控应激条件下离子跨质膜的选择吸收[29]。如Ca2+信号调控NFA(尼氟酸)对玉米根茎中阴离子(NO3-、Cl-和I-)的选择性吸收[30]。Al3+刺激Ca2+跨膜转运、诱导茶树根成熟区Ca2+信号,在Al3+处理下CaM(钙调蛋白)促进氟的吸收[31]。研究表明,茶树根系质膜Ca2+-ATPase活性被抑制时,引起细胞内Ca2+的外排,产生Ca2+信号,激活CaM,从而促进茶树对氟的吸收[27]。另有研究认为,氟激发了RLK(类受体蛋白激酶)基因的表达(RLKs是由植物基因组编码的信号转导基因的超家族),进而激活了Ca2+-ATPase活性,活化的Ca2+-ATPase可作为茶树摄取氟的载体促进氟的吸收和转运(图1)[11]。
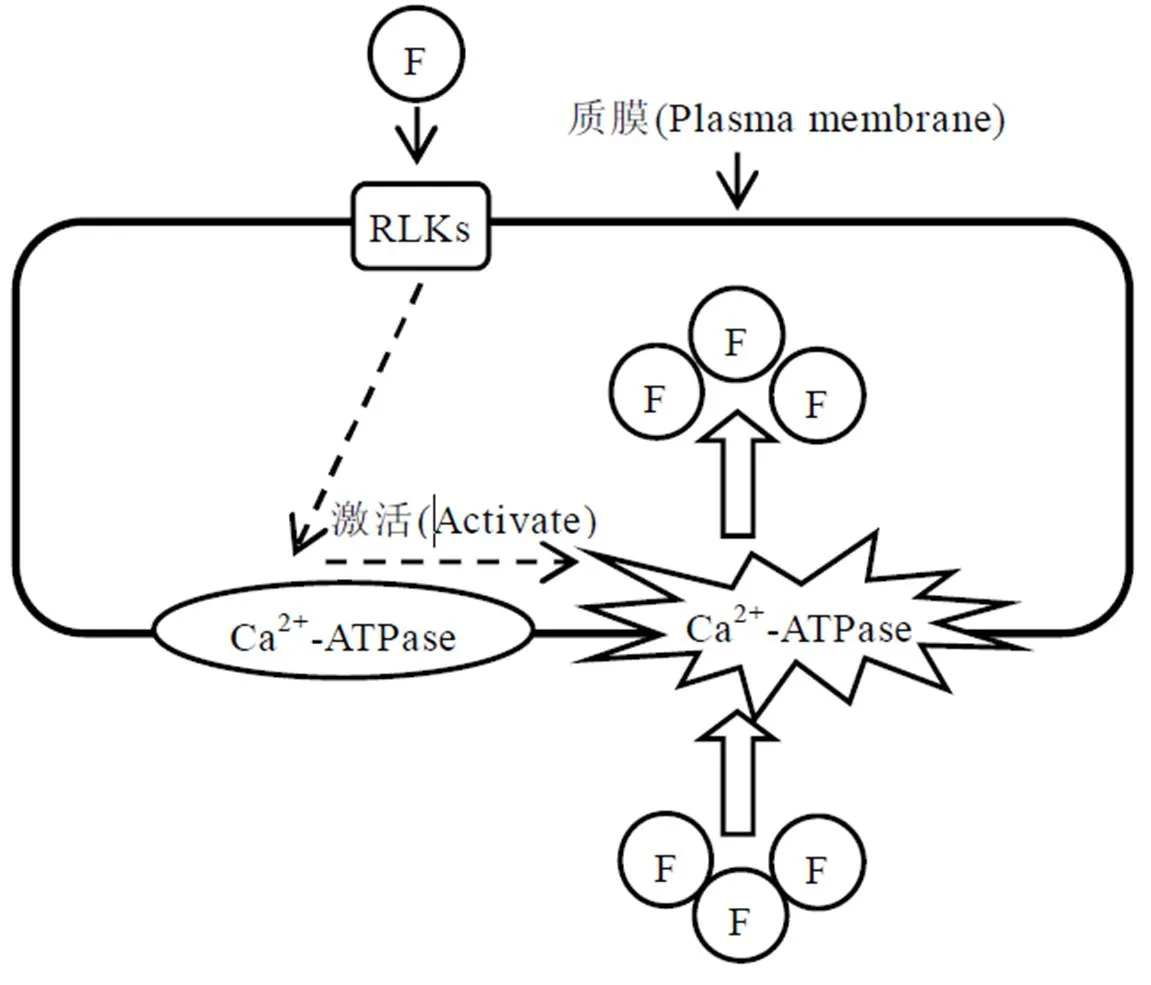
注:RLK在感知氟信号后激活Ca2+-ATPase,Ca2+-ATPase帮助氟进入细胞
Note: The RLKs activate Ca2+-ATPase after perceiving F signals, and Ca2+-ATPase helps F enter into cells
图1. 氟吸收的假设示意图
Fig. 1 Hypothetical schematic diagram of fluorine uptake
2.3 茶树根系对氟的被动吸收
2.3.1离子通道
离子穿过通道是沿跨膜浓度梯度或电位梯度扩散的顺势流动,与利用载体蛋白进行协助扩散相似,但区别于逆电化学势梯度的主动运输[32]。在分子水平上,协助扩散由单向转运体或通道介导[25]。有研究认为,茶树跨膜吸收氟涉及水通道和阳离子通道,但都不是主要途径,阴离子通道才是重要途径之一[4,33]。植物中氟的跨膜转运位点可能与转运Cl-的通道有关[34],Cl-通道是阴离子通道中的主要成员之一,可以介导Cl-、NO3-等阴离子的跨膜转运[35-37]。保卫细胞中的R型阴离子通道可渗透多种阴离子,且遵循SCN->NO3->Br->F-> I->Cl-的渗透顺序;S型阴离子通道除了SCN-不能被渗透,其他均可被渗透[38]。
许多单细胞生物和高等植物在其细胞膜中用氟输出蛋白来保持细胞质低氟浓度。现已知两个独立的氟输出蛋白家族,CLCF(Cl-通道F-/H+反向转运蛋白)和Fluc(小膜蛋白家族,又被称为crcB)[39]。前者是CLC超家族阴离子转运蛋白的一个亚类,它对F-/Cl-的选择性有特别强的电压依赖性,并表现出F-比Cl-较高的选择性,能从细胞质中逆转运F-[40-41];后者传输F-是通过跨膜通道进行热力学被动电扩散[39]。研究发现,氟输出蛋白家族的F-通道是“单管”转运蛋白,含有两个孔,其中孔Ⅱ在F-转运中起作用[42]。
2.3.2 Al-F络合吸收机制
铝是植物组织中氟的最强配体[43]。土壤中水溶性铝与水溶性氟含量呈显著正相关,表明土壤中的铝和氟能够以络合形态合作进入茶树体内[44]。研究表明,AlFn3-n络合体是茶树吸收氟的载体,而非运输形态[45]。Yang等[43]研究发现,茶树根细胞氟信号在NaF处理下比NaF和AlCl3共同处理强烈,说明氟铝溶液中的氟形态可能以AlF2+,AlF2+和AlF30等复合物存在,表明氟主要是以Al-F络合物被根系吸收。张显晨等[21]认为,高浓度Al3+存在时,茶树根系吸收主要氟形态是Al-F络合态。前人研究也证明了土壤中能形成Al-F配合物,更容易被茶树吸收和运输,并且AlF3促进了更多的氟吸收和向叶片的转运[45-46]。我们推测茶树根系吸收Al-F络合物可能需要载体蛋白介导,与植物吸收Fe3+需要YS(Yellow Stripe)转运体介导的螯合机制可能相似[47]。
3 茶树根系吸收氟的影响因素
3.1 温度
温度会影响根系对氟的摄取。当外界温度低于5℃,茶树对氟的吸收速率很低;当温度升到15℃,吸收速率增加约1倍;当温度达到35℃,吸收速率显著提高,为5℃时的3倍左右[48]。Zhang等[4]研究表明,茶树暴露于低温(5℃)时根系对氟的摄取受到强烈抑制,高温(35℃)时根系对氟的吸收量显著提高。王玉梅等[10]认为,低温(4℃)可能抑制了茶树根系质膜H+-ATPase活性,阻碍ATP的合成,减少水解产生的H+,降低质膜内外的质子驱动力,从而削弱茶树根系吸收氟的过程。
3.2 pH
pH被认为是影响茶树根系吸收氟的主要因素,因为pH显著影响土壤的氟形态,其中对水溶态氟和可交换态氟的影响最大[49]。谢忠雷等[16]认为,茶树对土壤中氟的吸收量与pH呈显著负相关,酸性土壤中的氟更易被吸收。Ruan等[50]研究表明,当培养溶液pH值在4.0~6.0范围内,茶树根系氟吸收量随着pH的升高先增加后减少,在pH 5.5时达到最大值,pH 4.0时较低。Zhang等[4]认为,在外界低氟浓度(18 µmol·L-1)下pH影响净F-吸收,pH 2.0和pH 5.0时的净F-吸收速率显著大于pH 4.0和pH 6.0,而更高氟浓度(43、150 µmol·L-1)下净F-吸收不受pH的影响。
3.3 陪伴离子
实际上,促进离子吸收的转运蛋白很少是特异性的,相同价态的离子之间竞争进入通道或结合载体蛋白是常见的[25]。王玉梅等[10]研究表明,氯降低茶树对氟的吸收,可能与Cl-与F-竞争同一载体的吸附位点有关。这与Calvo-Polanco等[51]观点一致,他认为Cl-可能对F-的吸收转运具有竞争性抑制作用,同时会干扰Cl-的膜转运。Ruan等[19]也认为,氟的吸收受氯的负面影响。除阴离子的影响外,阳离子也影响根系对氟的吸收,其中Al3+显著影响茶树中氟的吸收和转运,铝处理(≤0.5 mmol·L-1AlCl3)增加了根系中氟的含量,但降低了叶片中氟的含量[43]。Ca2+降低茶树的氟吸收及叶片氟含量,不仅仅是由于溶液和土壤中的CaF2沉淀或根中Ca-F络合物,更可能是因为Ca2+改变了茶树根系细胞壁结构和细胞膜渗透性,以及土壤溶液中氟形态及其数量,进而影响氟进入茶树体内[13,50,52]。
3.4 代谢抑制剂
阴离子通道代谢抑制A-9-C(剂蒽-9-羧酸),CCCP(羰基氰化物间氯苯腙)强烈抑制茶树对F-的吸收,而DIDS(二氢-4,4'-二异硫氰基芪-2,2'-二磺酸)对F-的吸收没有显著影响[4]。但Zhang等[27]研究发现,DIDS显著减弱了Al3+促进茶树对氟的积累,并且Ca2+和CaM也显著降低。另外,Ca2+-CaM和质膜电位去极化参与了NPPB[5-硝基-2-(3-苯基丙基氨基)苯甲酸]抑制茶树氟的积累[53]。这些结果表明离子通道抑制剂可以通过关闭阴离子通道而降低茶树对氟的吸收和积累。
4 研究展望
目前,有关报道茶树降氟措施的研究多侧重于栽培、加工、施肥等方面,而从分子机制方面研究降低茶树根系吸收氟的甚少。本文综述了茶树根系吸收氟的4种可能途径,即H+-ATPase或Ca2+-ATPase介导的跨膜主动吸收,离子通道蛋白和Al-F结合共转运的被动吸收,但其吸收过程和分子机制尚不清楚。
4.1 膜蛋白介导茶树根系吸收氟的机制
Ca2+-ATPase介导茶树吸收氟的RLK基因已被报道[11],一些质膜H+-ATPase基因也已经被鉴定与植物营养元素吸收有关。例如,玉米体内与NO3-摄取有关的和基因[54],水稻中参与磷的获取和转运的同种型基因[55]等。但是H+-ATPase介导的茶树吸收氟的相关基因目前还没有文献报道。研究表明,氟会提高茶树根系H+-ATPase活性[56],该酶活性的增加可能会促进茶树根系对氟的吸收,氟还可抑制玉米根系H+-ATPase[57]和酵母质膜H+-ATPase[58]活性,而该酶活性的变化部分地由该酶基因的表达模式变化引起[59]。因此,利用分子生物学手段,研究茶树跨膜吸收氟相关转运蛋白的作用及其基因的克隆和表达分析,将主效基因转化到烟草/拟南芥中进行功能验证,以揭示茶树跨膜吸收氟的分子机制,调控茶树根系对氟的选择吸收。此外,突破转基因茶树品种的选育及表达体系构建的障碍,为研究茶树根系跨膜吸收氟的分子机制提供试验材料,将是今后努力的重要方向。
4.2 调节茶树根系吸收氟的措施
按照农业部制定的行业标准(NY 659—2003),茶叶氟含量(F-)不能超过200 mg·kg-1。茶树新梢氟含量在100~300 mg·kg-1[60],成熟叶和老叶已经严重超标[5-6]。减少茶树根系对土壤氟的吸收富集是调控茶叶氟含量的重要措施,已有的研究多采用施肥,调节pH,添加生石灰和生物质炭改良剂等措施降低土壤氟的生物有效性,减少茶树对土壤氟的吸收富集[50,61-62]。但是,这些调控措施是否会影响茶树对其他营养元素的吸收,是否会影响茶叶中氨基酸、咖啡碱等次生代谢物质的含量及茶叶品质,目前还不清楚,值得进一步研究。此外,选育低氟茶树品种也是生产低氟茶叶产品的有效途径。
[1] Dey S, Giri B. Fluoride fact on human health and health problems: A review [J]. Medical & Clinical Reviews, 2016, 2(1): 2. DOI: 10.21767/2471-299X.1000011.
[2] 张楠, 张凌云. 茶叶中氟的安全性与检测方法研究进展[J]. 茶叶, 2009, 35(1): 3-6.
[3] Weinstein L H, Davison A. Fluorides in the environment: effects on plants and animals [M]. Oxfordshire: CABI, 2004.
[4] Zhang L, Li Q, Ma L, et al. Characterization of fluoride uptake by roots of tea plants (, (L.) O. Kuntze) [J]. Plant & Soil, 2013, 366(1/2): 659-669.
[5] Lu Y, Guo W F, Yang X Q. Fluoride content in tea and its relationship with tea quality [J]. Journal of Agricultural and Food Chemistry, 2004, 52(14): 4472-4476.
[6] Shu W S, Zhang Z Q, Lan C Y, et al. Fluoride and aluminium concentrations of tea plants and tea products from Sichuan Province, PR China [J]. Chemosphere, 2003, 52(9): 1475-1482.
[7] 董青华, 孙威江, 杨贤强. 茶树吸收氟的根际效应及富集机理研究进展[J]. 亚热带农业研究, 2009, 5(3): 162-166.
[8] 蔡荟梅, 彭传燚, 李成林, 等. 三个品种茶树氟富集特性及其在亚细胞中的分布[J]. 中国农业科学, 2013, 46(8): 1668-1675.
[9] 彭传燚, 陈静, 蔡荟梅, 等. 茶树对氟的吸收动力学特性研究[J]. 热带作物学报, 2013, 34(3): 495-500.
[10] 王玉梅, 柴如山, 郜红建. 茶树根系跨膜主动吸收氟的表观特征[J]. 农业环境科学学报, 2016, 35(8): 1473-1479.
[11] Li Q S, Lin X M, Qiao R Y, et al. Effect of fluoride treatment on gene expression in tea plant () [J]. Scientific Reports, 2017, 7(1): 9847. DOI: https://www.nature.com/articles/s41598-017-08587-6.
[12] Nagata T, Hayatsu M, Kosuge N. Identification of aluminium forms in tea leaves by27Al NMR [J]. Phytochemistry, 1992, 31(4): 1215-1218.
[13] Nagata T, Hayatsu M, Kosuge N. Aluminium kinetics in the tea plant using27Al and19F NMR [J]. Phytochemistry, 1993, 32(4): 771-775.
[14] Cao J, Luo S F, Liu J W, et al. Safety evaluation on fluoride content in black tea [J]. Food Chemistry, 2004, 88(2): 233-236.
[15] Yi X Y, Qiao S, Ma L F, et al. Soil fluoride fractions and their bioavailability to tea plants (L.) [J]. Environmental Geochemistry & Health, 2017, 39(5): 1005-1016.
[16] 谢忠雷, 陈卓, 孙文田, 等. 不同茶园茶叶氟含量及土壤氟的形态分布[J]. 吉林大学学报(地球科学版), 2008, 38(2): 293-298.
[17] 谢正苗, 吴卫红. 环境中氟化物的迁移和转化及其生态效应[J]. 环境工程学报, 1999(2): 40-53.
[18] Ruan J Y, Wong M H. Accumulation of fluoride and aluminium related to different varieties of tea plant [J]. Environmental Geochemistry and Health, 2001, 23(1): 53-63.
[19] Ruan J T, Ma L, Shi Y, et al. Uptake of fluoride by tea plant (L.) and the impact of aluminium [J]. Journal of the Science of Food & Agriculture, 2003, 83(13): 1342-1348.
[20] Horie H, Nagata T, Mukai T, et al. Determination of the chemical form of fluorine in tea infusions by19F-NMR [J]. Bioscience Biotechnology and Biochemistry, 1992, 56(9): 1474-1475.
[21] 张显晨, 郜红建, 张正竹, 等. 铝对氟在茶树体内吸收与分配的影响[J]. 食品科学, 2013, 34(5): 147-150.
[22] Palmgren M, Harper J. Pumping with plant P-type ATPases [J]. Journal of Experimental Botany, 1999, 50: 883-893.
[23] Sperandio M V L, Santos L A, Bucher C A, et al. Isoforms of plasma membrane H+-ATPase in rice root and shoot are differentially induced by starvation and resupply of NO3−, or NH4+[J]. Plant Science, 2011, 180(2): 251-258.
[24] Teakle N L, Tyerman S D. Mechanisms of Cl-transport contributing to salt tolerance [J]. Plant Cell & Environment, 2010, 33(4): 566-589.
[25] White P J. Ion uptake mechanisms of individual cells and roots: short-distance transport [J]. Marschners Mineral Nutrition of Higher Plants, 2012, 1(5): 7-47.
[26] Shen H , Chen J , Wang Z , et al. Root plasma membrane H+-ATPase is involved in the adaptation of soybean to phosphorus starvation [J]. Journal of Experimental Botany, 2006, 57(6): 1353-1362.
[27] Zhang X C, Gao H J, Yang T Y, et al. Al3+-promoted fluoride accumulation in tea plants () was inhibited by an anion channel inhibitor DIDS [J]. Journal of the Science of Food & Agriculture, 2016, 96(12): 4224-4230.
[28] Pedchenko V K, Nasirova G F, Palladina T A. Lysophosphatidylcholine specifically stimulates plasma membrane H+-ATPase from corn roots [J]. FEBS Letters, 1990, 275(1/2): 205-208.
[29] Pottosin I, Velarde-Buendía A M, Bose J, et al. Polyamines cause plasma membrane depolarization, activate Ca2+-, and modulate H+-ATPase pump activity in pea roots [J]. Journal of Experimental Botany, 2014, 65(9): 2463-2472.
[30] Roberts S K. Plasma membrane anion channels in higher plants and their putative functions in roots [J]. New Phytologist, 2006, 169(4): 647-666.
[31] Zhang X C, Gao H J, Wu H H, et al. Ca2+and CaM are involved in Al3+pretreatment-promoted fluoride accumulation in tea plants (L.) [J]. Plant Physiology and Biochemistry, 2015, 96: 288-295.
[32] 何龙飞, 刘友良, 沈振国, 等. 植物离子通道特征、功能、调节与分子生物学[J]. 植物学报, 1999, 16(5): 517-525.
[33] Zhang X C, Gao H J, Zhang Z Z, et al. Influences of different ion channel inhibitors on the absorption of fluoride in tea plants (L.) [J]. Plant Growth Regulation, 2013, 69(1): 99-106.
[34] Chapman B E, Kuchel P W. Fluoride transmembrane exchange in human erythrocytes measured with19F NMR magnetization transfer [J]. European Biophysics Journal, 1990, 19(1): 41-45.
[35] Chen Z, Beck T L. Free energies of ion binding in the bacterial CLC-ec1 chloride transporter with implications for the transport mechanism and selectivity [J]. Journal of Physical Chemistry B, 2016, 120(12): 3129-3139.
[36] Tyerman S D. Anion channels in plants [J]. Annual Review of Plant Biology, 1992, 43(1): 351-373.
[37] 戴松香, 陈少良. 植物根细胞离子通道研究进展[J]. 北京林业大学学报, 2005, 27(3): 98-103.
[38] White P J, Broadley M R. Chloride in soils and its uptake and movement within the plant: a review [J]. Annals of Botany, 2001, 88(6): 967-988.
[39] Stockbridge R B, Robertson J L, Ludmila K P, et al. A family of fluoride-specific ion channels with dual-topology architecture [J]. eLife, 2013, 2: e01084. DOI: 10.7554/eLife.01084.
[40] Stockbridge R B, Lim H H, Otten R, et al. Fluoride resistance and transport by riboswitch-controlled CLC antiporters [J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(38): 15289-15294.
[41] Brammer A E, Stockbridge R B, Miller C. F-/Cl-selectivity in CLCF-type F-/H+antiporters [J]. The Journal of General Physiology, 2014, 144(2): 129-136.
[42] Berbasova T, Nallur S, Sells T, et al. Fluoride export (FEX) proteins from fungi, plants and animals are ‘single barreled’ channels containing one functional and one vestigial ion pore [J]. Plos One, 2017, 12(5): e0177096. https://doi.org/10.1371/journal.pone.0177096.
[43] Yang Y, Liu Y, Huang C F, et al. Aluminium alleviates fluoride toxicity in tea () [J]. Plant & Soil, 2016, 402(1/2): 179-190.
[44] Xie Z L, Chen Z, et al. Distribution of aluminum and fluoride in tea plant and soil of tea garden in central and southwest China [J]. Chinese Geographical Science, 2007, 17(4): 376-382.
[45] Takmaz-Nisancioglu S, Davison A W. Effects of aluminium on fluoride uptake by plants [J]. New Phytologist, 1988, 109(2): 149-155.
[46] Xie Z M, Ye Z H, Wong M H. Distribution characteristics of fluoride and aluminum in soil profiles of an abandoned tea plantation and their uptake by six woody species [J]. Environment International, 2001, 26(5): 341-346.
[47] Kobayashi T, Nishizawa N K. Iron uptake, translocation, and regulation in higher plants [J]. Ann. rev. plant Biol, 2012, 63(1): 131-152.
[48] 张磊. 茶树氟吸收动力学特性的研究[D]. 北京: 中国农业科学院, 2008.
[49] 吴卫红, 谢正苗, 徐建明, 等. 不同土壤中氟赋存形态特征及其影响因素[J]. 环境科学, 2002, 23(2): 104-108.
[50] Ruan J, Lifeng M A, Shi Y, et al. The impact of pH and calcium on the uptake of fluoride by tea plants (L.) [J]. Annals of Botany, 2004, 93(1): 97-105.
[51] Calvo-Polanco M, Zwiazek J J, Jones M D, et al. Effects of NaCl on responses of ectomycorrhizal black spruce (), white spruce () and jack pine () to fluoride [J]. Physiologia Plantarum, 2009, 135(1): 51-61.
[52] 马立锋, 阮建云, 石元值, 等. 钙[Ca(NO3)2和CaO]对茶树氟吸收的影响[J]. 土壤通报, 2005, 36(1): 85-87.
[53] Zhang X C, Gao H J, Yang T Y, et al. Anion channel inhibitor NPPB-inhibited fluoride accumulation in tea plant () is related to the regulation of Ca2+, CaM and depolarization of plasma membrane potential [J]. International Journal of Molecular Sciences, 2016, 17(1): 57. https://doi.org/10.3390/ijms17010057.
[54] Santi S, Locci G, Monte R, et al. Induction of nitrate uptake in maize roots: expression of a putative high-affinity nitrate transporter and plasma membrane H+-ATPase isoforms [J]. Journal of Experimental Botany, 2003, 54(389): 1851-1864.
[55] Chang C, Hu Y, Sun S, et al. Proton pump OsA8 is linked to phosphorus uptake and translocation in rice [J]. Journal of Experimental Botany, 2009, 60(2): 557-565.
[56] 王玉梅. 茶树根系跨膜吸收氟的微观机制和转录组学特征[D]. 合肥: 安徽农业大学, 2017.
[57] Facanha A R, De Meis L. Inhibition of maize root H+-ATPase by fluoride and fluoroaluminate complexes [J]. Plant Physiology, 1995, 108(1): 241-246.
[58] Pedersen J T, Falhof J, Ekberg K, et al. Metal fluoride inhibition of a P-type H+Pump: Stabilization of the phosphoenzyme intermediate contributes to post-translational pump activation [J]. Journal of Biological Chemistry, 2015, 290(33): 20396-20406.
[59] Han N, Ji X L, Du Y P, et. al. Identification of a novel alternative splicing variant of VvPMA1 in grape root under salinity [J]. Frontiers in Plant Science, 2017, 8: 1-10.
[60] 刘艳丽, 金孝芳, 曹丹, 等. 茶树铝、氟富集研究进展[J]. 植物科学学报, 2016, 34(6): 972-977.
[61] 张永利, 王烨军, 廖万有, 等. 施氮对茶园土壤氟和茶树新梢氟含量的影响[J]. 中国生态农业学报, 2015, 23(12): 1562-1570.
[62] Gao H J, Zhang Z Z, Wan X C. Influences of charcoal and bamboo charcoal amendment on soil-fluoride fractions and bioaccumulation of fluoride in tea plants [J]. Environmental Geochemistry and Health, 2012, 34(5): 551-562.
Physiological and Molecular Mechanisms of Transmembrane Fluoride Uptake by Tea Roots
XU Jiajia1,2, GUANG Min2, SHI Shulin2, GAO Hongjian1,2*
1. State Key Laboratory of Tea Plant Biology and Utilization, School of Tea and Food Science and Technology of Anhui Agricultural University, Hefei 230036, China; 2. Anhui Province Key Laboratory of Farmland Ecological Conservation and Pollution Prevention, School of Resources and Environment of Anhui Agricultural University, Hefei 230036, China
Tea plant is a fluoride hyper-accumulator and most of fluoride is accumulated in the leaves. Drinking tea is an important way to absorb fluoride for human, which affects human health. Fluoride is usually absorbed from the soil by tea roots. However, the physiological and molecular mechanisms of transmembrane fluoride uptake by the tea roots were still poorly documented. Therefore, this paper reviewed the active and passive pathways of fluoride uptake by the roots of tea. The active transmembrane uptake fluoride process and molecular mechanism by H+-ATPase and Ca2+-ATPase, the role and microscopic process of ion channel and Al-F complexation by passive fluoride uptake were analyzed. The main influencing factors and control measures of fluoride accumulation in tea roots were also investigated. In order to reveal the molecular mechanism of transmembrane fluoride uptake, regulate the selective fluoride uptake in tea roots, and ensure the tea quality and safety for consumption, cloning, expression, and functional verification of transport proteins and genes related to fluoride transmembrane uptake in tea plant should receive more attention in future studies.
tea roots, fluoride, transmembrane uptake, physiological and molecular mechanism, influencing factor
S571.1
A
1000-369X(2019)04-365-07
2019-01-17
2019-03-22
安徽省自然科学基金(1808085QC56)
徐佳佳,女,硕士研究生,主要从事茶树吸收氟元素的生理与生物学方面的研究。*通信作者:hjgao@ahau.edu.cn
猜你喜欢
工业水处理(2022年12期)2023-01-05
工业水处理(2022年12期)2022-12-29
中华实验眼科杂志(2022年8期)2022-11-15
茶道(2022年3期)2022-04-27
中国药房(2022年7期)2022-04-14
山西大同大学学报(自然科学版)(2021年2期)2021-05-04
福建茶叶(2020年5期)2020-12-23
福建茶叶(2020年10期)2020-12-22
农技服务(2020年1期)2020-12-17
医药前沿(2020年28期)2020-12-02
