1-磷酸鞘氨醇及其信号通路在炎症相关性疾病中的作用
2019-08-20 01:40卜妍红孙明慧戴学静
中国药理学通报 2019年8期
卜妍红,吴 虹,孙明慧,张 衡,戴学静,王 言,占 翔
(安徽中医药大学药学院,新安医学教育部重点实验室,中药复方安徽省重点实验室,安徽 合肥 230012)
鞘脂及其代谢酶是体内的关键介质,鞘氨醇激酶(sphingosine kinases,SphKs)及其脂质产物1-磷酸鞘氨醇(sphingosine 1-phosphate,S1P)在信号传导以及疾病,特别是在慢性炎症疾病和自身免疫疾病的发生、发展中起重要作用[1]。S1P是一种具有多种生物活性的鞘脂类代谢产物,广泛存在于血小板、红细胞、中性粒细胞和淋巴液、血液等细胞和体液中。神经酰胺(ceramide,Cer)经神经酰胺合成酶脱羧化作用后,转化成鞘氨醇(sphingosin,Sph),不同细胞因子和生长因子,如肿瘤坏死因子α(tumor necrosis factor α,TNF-α)、白介素-1β(interleukin-1β,IL-1β)和血管内皮生长因子(vascular endothelial growth factor,VEGF)可激活SphKs,然后SphKs催化Sph磷酸化生成S1P。S1P经S1P裂解酶(S1P lyase,SPL)和S1P磷酸酶(S1P-phosphatases,SPP)发生不可逆的降解,以维持机体内S1P的动态平衡。其中,“鞘磷脂变阻器”(Cer/S1P rheostat)是决定细胞的存亡和增殖的关键[2],S1P促进细胞的存活和增殖,其前体Cer和Sph则抑制细胞存活、促进细胞凋亡。细胞内的Cer、Sph和由SphKs介导的酶促反应共同维持动态平衡,从而决定细胞的存活,维持细胞的生理功能。
S1P生成后,可在细胞内、外发挥作用。一方面,通过自分泌和旁分泌的方式作用于细胞表面受体,发挥其细胞外效应,包括细胞增殖和迁移、炎性细胞因子浸润、血管生成、自身免疫等。另一方面,发挥细胞内第二信使功能,直接作用于细胞内靶点,调控Ca2+水平、基因转录、蛋白质修饰等。
1 S1P胞外作用方式
S1P的主要作用方式是胞外作用,即质膜上产生的S1P分泌到细胞外,与胞膜上S1P受体(S1P receptors,S1PRs)结合发挥作用。S1PRs是G蛋白偶联受体(G protein-coupled receptors,GPCRs)家族成员,包括S1PR1~S1PR5五个亚型。S1PR1~S1PR5均与Gi/o偶联,S1PR2、S1PR3与Gq偶联,除S1PR1外,其余的S1PRs均与G12/13偶联[3],可以激活Ras/ERK1/2、PI3K/Akt等下游信号通路,发挥多种生物学效应,包括调节细胞增殖、迁移、凋亡以及炎症因子和黏附因子生成。
1.1 Ras/Raf/MEK/ERK途径细胞外信号调节激酶(extracellular signal regulated kinase,ERK)是一类丝/苏氨酸蛋白激酶,ERK家族有5个亚族,为ERK1~ERK5,ERK1和ERK2是ERK家族中研究最彻底的。作为传递丝裂原信号的信号转导蛋白,其正常情况下存在于胞质,受多种刺激因子如细胞因子、生长因子、癌基因、GPCRs的配体等激活,激活后转移至细胞核,调节转录因子活性,介导不同的细胞效应。其中,最经典的为Ras/Raf/MEK/ERK激活途径。
当S1P与S1PRs结合后,生长因子受体结合蛋白2(growth factor receptor-bound protein 2,Grb2)识别激活的受体并与之结合,同时Grb2的SH3结构域与鸟苷酸交换因子SOS的C端序列相结合,形成S1PRs-Grb2-SOS复合物。该复合物促进胞质蛋白SOS向膜聚集,并在Ras蛋白附近形成高浓度的SOS,SOS与Ras-GDP结合,并转化为Ras-GTP,Ras构象改变,由失活态转变为激活态,Ras活化。Raf在N端和C端结构域中有与Ras结合的重要区域,Ras作为上游激活蛋白,与c-Raf的两个区域高度亲和后,Raf被募集到质膜进行活化,激活Raf。Raf被激活后,其C端催化区能与MEK(MAPKK)结合,并磷酸化其催化区中2个丝氨酸,从而激活MEK。MEK作为一种双重特异性激酶,催化ERK酪氨酸和苏氨酸残基磷酸化,从而高度选择性地激活ERK[4-5]。
Ras/Raf/MEK/ERK不仅广泛地调节细胞功能,还在疾病的发病机制及病理过程中发挥重要作用。S1PR1经MEK1/2-ERK1/2信号通路,调节人脐静脉内皮细胞的迁移,血管内皮细胞迁移对于维持血管正常功能和结构稳定具有不可忽视的作用,从而发挥如抗动脉粥样硬化、抗炎以及内皮保护功能等[6]。S1PR1和S1PR3通过ERK信号转导,增加人骨髓间充质干细胞(mesenchymal stem cell,MSC)迁移,MSC介导的软骨损伤修复在于增加基质合成、软骨细胞增殖、调节免疫反应。软骨损伤后,大量分泌多种炎性细胞因子,包括IL-1β、IL-6、TNF-α和基质金属蛋白酶(matrix metalloproteinase,MMP)等,导致炎症加重、软骨基质降解和软骨凋亡。但当ERK信号通路活化后,MSC迁移至损伤部位,减少炎性细胞的浸润和炎性细胞因子的分泌[7]。此外,在动物实验中,抑制c-Raf-1和c-Myc的表达,可明显减弱类风湿性关节炎(rheumatoid arthritis,RA)患者成纤维样滑膜细胞(fibroblast-like synoviocytes,FLS)对SCID鼠模型软骨的侵蚀[8],其中,c-Raf-1是ERK信号通路的上游分子,c-Myc是ERK信号通路下游转录因子。以上结果提示,ERK信号通路在介导RA发病过程中发挥重要作用,通过调节ERK通路能改善RA的病情发展。因此,为了探索生命活动最基本的生理机制和过程,寻找针对性的治疗方法,我们深入研究信号转导过程及其涉及到的上游蛋白和各种激酶的激活机制。
1.2 PI3K/Akt途径PI3K/Akt信号通路是炎症、肿瘤等发生发展过程中一条重要的信号通路。磷酯酰肌醇-3激酶(phosphatidylinositol 3-kinase,PI3K)是由催化亚基p110和调节亚基p85所构成的异源二聚体,具有脂类激酶(磷脂酰肌醇激酶)和蛋白激酶(丝氨酸/苏氨酸蛋白激酶)的双重活性,能被多种因子,如血小板衍生因子、表皮生长因子等激活,发挥促进细胞增殖和抑制细胞凋亡的正性调节作用。Akt是一种丝氨酸/苏氨酸蛋白,多种生长因子、激素、抑癌基因PTEN和Ras均可调节Akt的活化[9]。
S1P与膜表面受体结合后,PI3K将收到来自GPCRs的信号,p85调节亚基即被募集到质膜附近,p110亚基通过与p85亚基结合,催化PIP2磷酸化以产生脂质第二信使PIP3,PTEN使PIP3去磷酸化,抑制PI3K的产生。PIP3和含有PH结构域的PKB结合,促使Akt从细胞质转移至细胞膜,并在3-磷酸肌醇依赖性蛋白激酶1(3-phosphoinositide-dependent protein kinase D1,PKD1)和哺乳动物雷帕霉素靶复合体2(mammalian target of rapamycin complex 2,mTORC2)的作用下,分别磷酸化Akt蛋白上的苏氨酸磷酸化位点(Thr308)和丝氨酸磷酸化位点(Ser473),激活Akt[10]。活化的Akt通过磷酸化作用激活或抑制其下游靶点蛋白,进而调控细胞的增殖、凋亡、分化、代谢等(Fig 1)。
内皮祖细胞(endothelial progenitor cells,EPCs)在内皮细胞修复与炎症血管新生中起重要作用。有研究显示[11],抑制Akt的活性和磷酸化可以减少S1P诱导的EPCs增殖,S1P增强EPCs的增殖主要通过S1PR3介导,减少H2O2诱导的EPCs凋亡主要通过S1PR1介导;S1PR1/S1PR3/PI3K/Akt信号通路在S1P刺激EPCs后,有助于血管新生和内皮化。因此,增加EPCs的数量和提高这些细胞的活性可能是血管炎症疾病的一个治疗靶点。众所周知,EPCs能够通过增殖、迁移修复受损血管,并最终分化为内皮细胞(endothelial cell,EC)。在EC中,S1PR2通过促进PI3K/Akt信号通路,增强TNF-α诱导的促炎因子和黏附因子,如VEGF、细胞间黏附因子1(intercellular cell adhesion molecule-1,ICAM-1)、单核细胞趋化蛋白1(monocyte chemotactic protein-1,MCP-1)的表达来加重血管壁炎症[12]。VEGF促进血管新生,新生血管不仅为滑膜增生提供氧气和营养物质,另一方面,随着炎性细胞和所产生的炎性介质的流入,也促进了滑膜炎症的持续存在。而PTEN过度表达或者PI3K/Akt抑制剂可以使VEGF表达减少,因此,抑制其表达可改善RA的病理过程;上调黏附分子的水平有助于多形核细胞对炎性组织的募集、介导中性粒细胞的黏附性,以便于中性粒细胞通过血管内皮屏障并和上皮细胞相互作用,从而起到促动脉粥样硬化的作用。
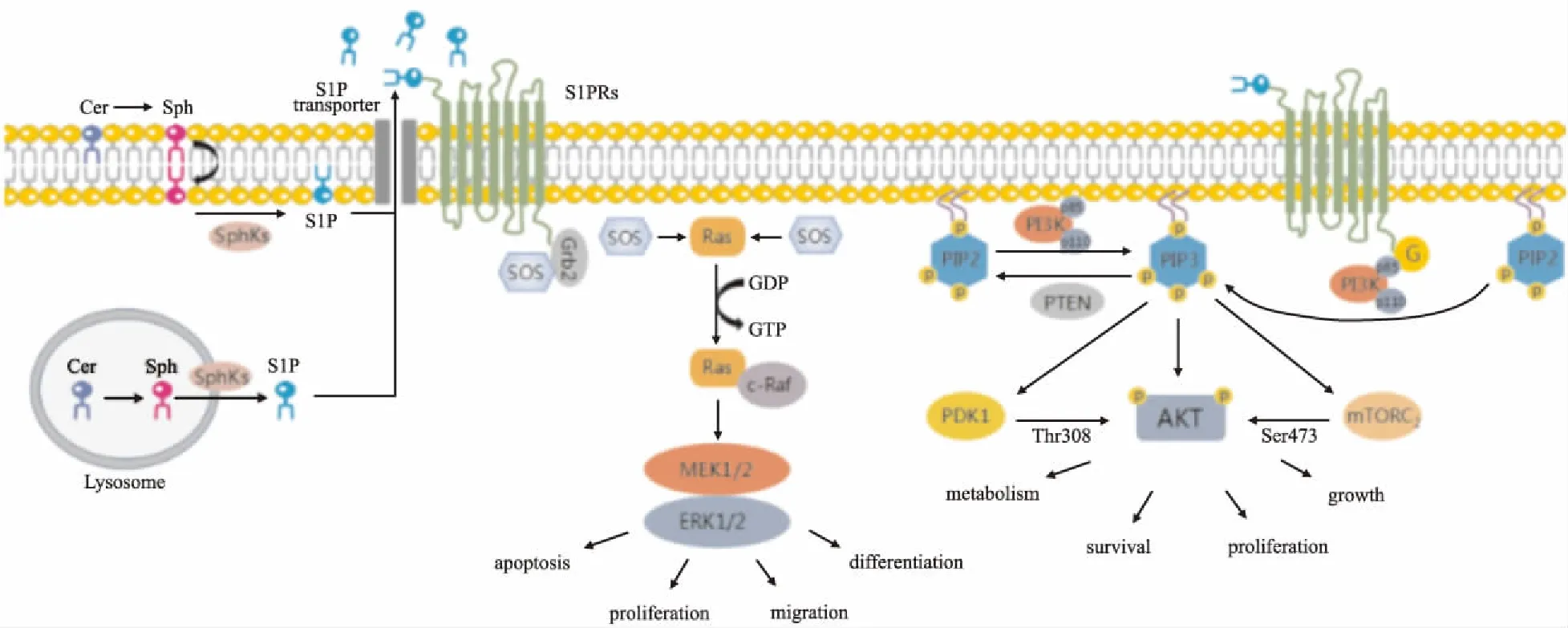
Fig 1 Extracellular function of S1P
Cer in lysosome and plasma membrane is decarboxylated and converted to Sph. Activated SphKs catalyzes the phosphorylation of Sph, leading to the production of S1P. S1P transports outside the cell and acts on GPCRs on cell surface. 1.Ras/Raf/MEK/ERK pathway: When S1P binds to S1PRs, Grb2 recognizes and binds to the activated receptors. Meanwhile, Grb2 binds to SOS to form the S1PRs-Grb2-SOS complex, which promotes SOS aggregation in Ras and mediates cell apoptosis, proliferation, migration and differentiation after Ras/Raf/MEK/ERK cascade activation. 2. PI3K/Akt pathway: After the binding of S1P to S1PRs, the p85 subunit is recruited and bound to the P110 subunit, which catalyzes the phosphorylation of PIP2 to produce PIP3. Under the action of PKD1 and mTORC2, PIP3 phosphorylates Thr308 and Ser473 on Akt protein respectively and activates Akt, which regulates cell growth, proliferation and metabolism.
2 S1P胞内作用方式
随着S1P的发现,人们对S1PRs多种多样的生物学功能进行了大量研究,但已有证据表明,S1P的某些下游生物学效应并不直接依赖于胞外受体,而是S1P行使类似第二信使的功能直接作用于胞内靶点,调节相关生物过程。目前较为明确的S1P胞内靶点是组蛋白去乙酰化酶(histone deacetylase,HDAC)和肿瘤坏死因子受体辅助因子2(TNF receptor-associated factor 2,TRAF2)。
2.1HDACHDAC是一类蛋白酶,在哺乳动物细胞中分为4类,其中,HDAC1、HDAC2、HDAC3和HDAC8组成的Ⅰ类HDAC,在所有组织细胞的细胞核中普遍表达。作为表观遗传调节因子,HDAC使核组蛋白和非组蛋白可逆地脱乙酰化,广泛调节基因的表达。组蛋白乙酰化转移酶(histone acetyl transferases,HATs)和HDAC之间存在的平衡,影响组蛋白的乙酰化和去乙酰化水平。当平衡发生改变时,疾病相关基因的失调将导致慢性炎症和癌症等疾病[13]。
S1P是HDAC内源性调节因子,作用类似于HDAC抑制剂,通过影响HDAC的活性调控基因表达。细胞核内的SphKs选择性富集,产生的S1P通过特异性结合HDAC抑制其活性,进而保护组氨酸末端的赖氨酸不被去乙酰化。组蛋白乙酰化水平升高后,DNA与组蛋白八聚体解离,使各种转录因子能与DNA特异性结合,激活基因转录(Fig 2)。HDAC抑制剂具有调节炎症基因表达、细胞增殖和DNA修复,对机体的不良反应小的特点,因此,被广泛用于各种慢性炎症疾病及恶性肿瘤的治疗。RA的一大病理特征是FLS的异常增殖和炎性细胞浸润,HDAC抑制剂有效下调了RASFs中促炎因子IL-6、IL-8 mRNA表达,来减轻滑膜细胞异常增殖和关节炎症[14]。HDAC1在RA患者的滑膜组织中高度表达,因此,设计靶向HDAC1的HDAC抑制剂作用于TNF-α刺激的单核细胞和破骨细胞,减少了炎性细胞因子和趋化因子,如IL-1、MCP-1、巨噬细胞炎性蛋白1α的mRNA表达,减轻炎症和骨质流失,从而达到治疗RA的效果[15]。HDAC抑制剂不仅广泛作用于不同类别的HDAC靶标,还在转录调节和蛋白质修饰中起重要作用,因此,HDAC抑制剂具有治疗潜力。
2.2TRAF2TRAF是衔接蛋白家族,通过连接细胞表面受体和下游激酶级联反应,导致转录因子激活,参与炎症、细胞凋亡、细胞存活等多种反应。TRAF家族由TRAF1-TRAF7组成,它们共享1个由N端卷曲螺旋结构TRAF-N和TRAF-C组成的TRAF结构域,虽然不同TRAF蛋白的TRAF结构域高度相似,但TRAF-C结构域的差异决定了与特异性受体和下游效应蛋白的相互作用,从而介导细胞中不同的生理作用[16]。表达最广泛的TRAF2作为衔接蛋白和调控因子,在几乎所有分支通路中起作用,主要激活的NF-κB和JNK在许多生理病理过程中发挥重要作用,如免疫、炎症反应、细胞增殖与凋亡、肿瘤的发生。
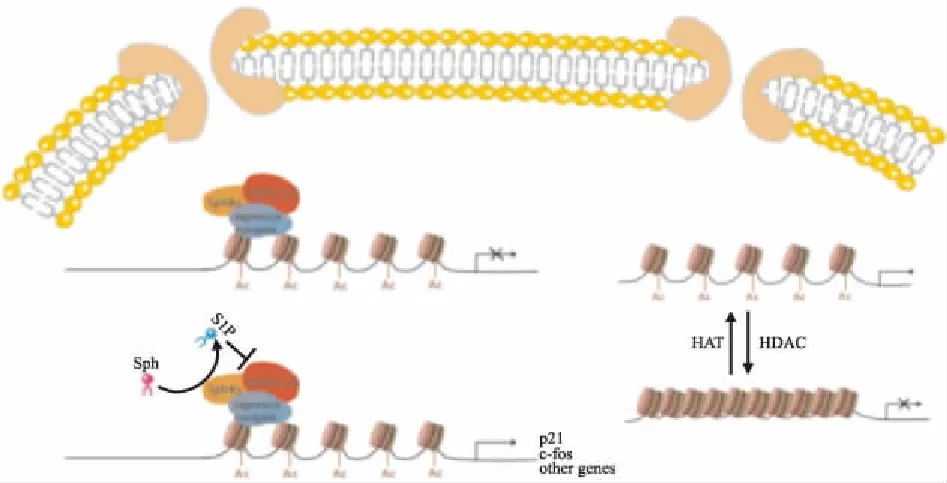
Fig 2 Intracellular function of S1P——HDAC pathway
HAT acetylates histones, dissociates DNA from histone octamers, and activates gene transcription by specific binding of transcription factors to DNA. HDAC deacetylates histones, allowing histones to bind tightly to DNA. Chromatin is densely curled and gene transcription is inhibited. SphKs catalyzes the production of S1P, which is similar to HDAC inhibitors. By specifically binding to HDAC, the activity of S1P is inhibited and the level of histone acetylation increases, which promotes gene transcription.
TRAF2不仅是S1P在胞内的直接作用靶点,也是TNF-α触发NF-κB信号的关键元件。胞质中的SphK1结合于TRAF2的指环结构域(ring domain),产生的S1P可以直接与TRAF2结合,并激活E3泛素连接酶活性。活化的TRAF2可以使受体相互作用蛋白(receptor-interacting protein,RIP)多泛素化,之后募集并激活IκB激酶,这些启动NF-κB通路的反应是由细胞内的S1P介导的,与细胞膜表面的GPCRs无关[17]。激活的NF-κB易位至细胞核,作用于靶基因特异性结合位点,并促进基因的表达(Fig 3)。有研究表明[18],小胶质细胞(microglia,MG)作为中枢神经系统中的固有免疫细胞活化后,产生过量的促炎性细胞因子,如TNF-α、IL-1β和其他促炎介质,参与神经元炎症损伤。在MG中,SphK1通过TRAF2/NF-κB途径,抑制IL-17的减少以及神经元凋亡。因此,阻断TRAF2/NF-κB轴可能是控制神经在脑内的免疫反应的潜在治疗目标。另有研究表明[19],若经TRAF2激活下游NF-κB信号通路,活化的TRAF2分子参与介导PGE2-EPs-cAMP信号通路的异常,进一步降低细胞内cAMP水平,加重滑膜功能紊乱。滑膜组织在RA中起到重要的作用,滑膜细胞不仅增殖,还增加迁移和流动性,侵入并破坏软骨,从而加重病情。此外,在IL-Iβ诱导的骨关节炎(osteoarthritis,OA)中,软骨细胞活力下降、细胞凋亡率增加,IL-6、IL-8和TNF-α的蛋白表达水平明显上升。与正常软骨组织相比,OA患者软骨组织中TRAF2水平明显升高,TRAF2沉默恰好反转IL-1β对软骨细胞增殖、凋亡、炎症反应的影响,从而减缓OA的发生、发展[20]。
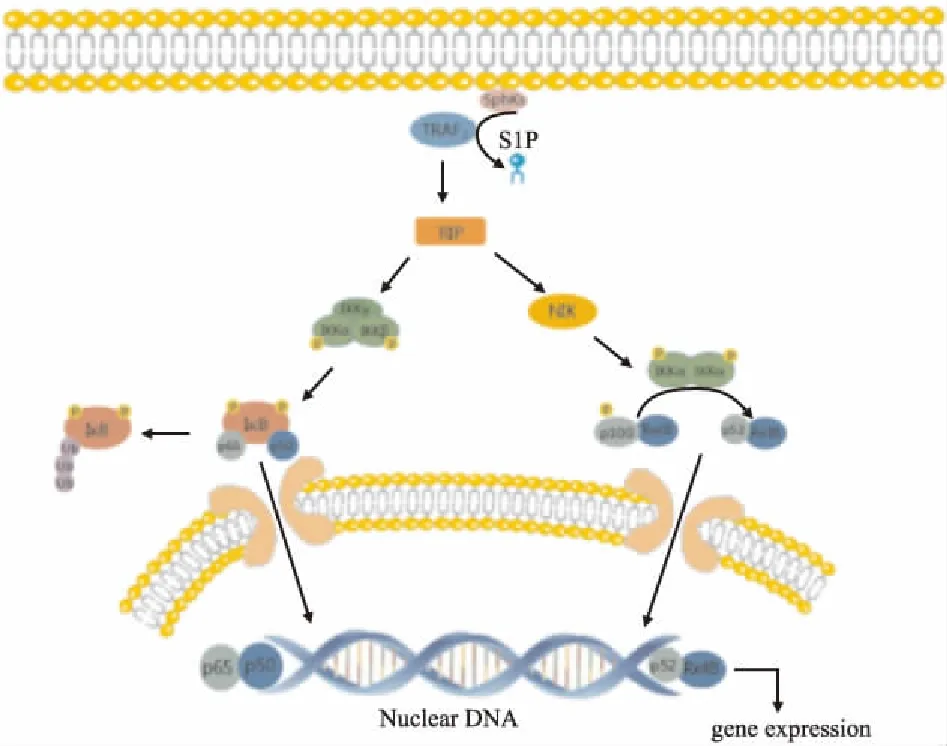
Fig 3 Intracellular function of S1P——TRAF2/NF-κB pathway
SphK1 binds to TRAF2, allowing the S1P direct binding to TRAF2 and then multiubiquitizes RIP to initiate the NF-κB pathway. 1. Classic pathway: RIP recruits and activates IKK, which degrades IκB and releases NF-κB from trimers. NF-κB specifically binds to the κB site of the target gene to regulate the transcriptional activity of downstream genes. IKKβ plays a major role in this process. 2. Non-classical pathway: NIK activated by RIP activates IKKα selectively, which degrades P100 into p52. The dimer of p52 and Rel B enters the nucleus and activates the downstream gene transcription by activating NF-κB. This process depends on NIK and IKKα, not IKKβ and IKKγ.
3 S1P信号通路相关药物研究进展
根据对S1P的研究,主要是从3个方向对S1P信号通路进行调控。S1P受体调节剂是通过对不同S1PRs亚型激动或抑制进行调控,如研究较多的S1P类似物芬戈莫德(fingolimod,FTY720);SphKs抑制剂是影响S1P的合成和降解过程;S1P特异性抗体是直接靶向S1P的抗体,通过清除血液或其他部位的S1P分子来发挥作用。
3.1 S1P受体调节剂靶向S1P信号传导的药物大多数都针对于S1PRs,而不是配体或其他代谢途径,是因为GPCRs是高度多样化的细胞表面受体,介导几乎所有细胞类型的基本过程;其次,尽管已发现S1P的细胞内靶标,但S1P的作用主要还是由S1PRs介导。FTY720正是通过S1PRs介导发挥作用,说明S1PRs是经临床验证的治疗药物靶标。
FTY720是首个口服用于治疗多发性硬化症(multiple sclerosis,MS)的新型免疫抑制剂。作为鞘氨醇类的前药,并不直接抑制淋巴细胞的激活和增生,而是经SphK1磷酸化成活性代谢物p-FTY720,与特定的S1PRs结合(除了S1PR2),通过影响趋化因子介导的淋巴细胞归巢,减少外周血淋巴细胞数,并抑制淋巴细胞活性,发挥免疫抑制的作用[21]。正常情况下,S1PRs与S1P结合后很快失活,并内化到细胞质囊泡中,但会再循环到质膜以用于随后的信号传导。相反,与FTY-720结合后,S1PRs不会再循环到质膜,它被保留在囊泡中被溶酶体降解,最终导致细胞表面的功能活性受体S1PRs耗尽。现已证明[22],FTY720可降低滑膜中IL-6和TNF-α的表达来缓解滑膜炎症和炎症细胞浸润关节组织,最终起到治疗RA的作用。此外,由于其亲脂性,FTY720穿过血脑屏障,作用于血脑屏障的内皮细胞时,会减少炎性细胞因子诱导的细胞死亡;FTY720处理明显下调脂多糖刺激的MG释放炎症介质,缓解神经炎症[23]。FTY720的临床成功,极大地鼓励了研究人员以S1P为切入点展开研究,尤其是S1P代谢中涉及的重要蛋白质,如S1PRs、SphKs和SPL。
3.2 SphKs抑制剂由于调节S1P代谢途径相对简单明确,该代谢途径中主要的酶SphKs成了研究的目标。尽管此方法靶向性不高,无法针对特定的受体,但能够调节整体S1P水平,当治疗的靶受体不明确时或未知时,调节总S1P水平成为了可行的方法。
SphKs抑制剂的作用机制主要是抑制SphKs的活性,减少Cer/Sph向S1P的转化,不仅能降低促细胞存活与增殖的S1P合成,同时还能增加促凋亡与周期阻滞的Cer/Sph生成,SphKs是维持这种平衡的重要调节剂。在多种慢性炎症疾病及癌症中SphK1过表达,参与细胞的增殖迁移以及血管生成,新生血管不仅为细胞增殖提供营养物质,另一方面, 随着炎性细胞和所产生的炎性介质流入,也促进了炎症的持续存在。在许多类型的癌症和炎症性疾病中,SphK1过度表达,因此,SphK1是治疗炎症发展的主要目标。ABC294640作为特异性SphK2抑制剂增加了Cer的积累,降低存活的S1P含量,抑制了小鼠肝脏炎症并激活细胞凋亡机制,从而抑制肿瘤的生长。尽管SphKs抑制剂对于各种癌症模型都有效,但在关节炎中的作用存在着争议。大鼠实验使用siRNA降低SphK2表达,不能明显改善关节炎的症状,ABC294640却引起关节炎症状加重[24]。因此,与阻断SphK1的明显抗炎作用相比,SphK2虽可作为癌症治疗或增强免疫反应的候选者,但在自身免疫或自身炎症疾病中应避免其阻断。针对鞘氨醇激酶途径的治疗需要特异性地靶向鞘氨醇激酶,以更好地利用其治疗潜力。
4 展望
自从发现S1P作为GPCRs家族的高亲和力配体后,鞘脂生物学领域已成为一大研究热点。S1P参与多种重要的生物过程,其功能不仅存在于它的化学结构中,还通过不同的信号通路引起多样的生物学变化,包括生理病理过程。这些累积的发现证明,S1P是调节免疫应答、炎症过程、肿瘤以及心血管系统的关键信号分子。目前,对S1P介导生物功能的研究越来越多,但缺乏对作用机制和各个生物功能关联性的研究。对S1P多样的受体亚型、复杂的下游信号和信号通路的重叠表达的深入探索,正契合了炎症免疫反应软调节这一概念,多种靶细胞的关键分子活性的平衡调控,是研究炎症相关性疾病治疗药物的重要方向[25]。靶向配体和受体功能的药物已经开发并处于临床前和临床发展的不同阶段,这一领域的明显进步是由遗传学和药理学方法所促进的,各种复杂的信号系统是潜在的有趣的治疗靶点,可能在未来带来更大的发现。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年12期)2021-12-30
中老年保健(2021年3期)2021-12-03
天津医科大学学报(2021年3期)2021-07-21
中华养生保健(2020年9期)2021-01-18
世界农药(2019年2期)2019-07-13
科学24小时(2018年1期)2018-01-10
医学研究杂志(2015年2期)2015-06-10
中国民族民间医药·上半月(2013年1期)2013-11-01