
围产前期添加山楂和黄芪混合物对奶牛血浆代谢组的影响
2019-08-08 03:32张萌刘国林李向龙陈永宏白玲荣罗芳李亚超陶金忠
生物技术通报 2019年8期
张萌 刘国林 李向龙 陈永宏 白玲荣 罗芳 李亚超 陶金忠
(宁夏大学农学院,银川 750021)
围产期是奶牛最重要的时期,同时也是奶牛能量代谢病的高发时期[1]。围产期奶牛动员脂肪组织,导致神经内分泌因子增加和免疫状态发生改变。神经内分泌因子增加促进非酯化脂酸(Non-esterified fatty acids,NEFA)和甘油进入血液及肝脏,从而引起由脂肪代谢产生一系列变化,导致能量代谢障碍性疾病[2]。而免疫力降低,奶牛抵御外来入侵能力减弱,影响围产期奶牛健康状况,易发各种产科疾病发生,造成巨大的经济损失。
黄芪味甘性温,归脾、肺经,属补气药物,具有保护心肌细胞、强心肌、降低血脂、提高免疫功能等药理作用[3-4]。黄芪含有多糖类、皂苷类、黄酮类和氨基酸类等多种活性成分。其中黄芪多糖具有免疫调节、抗肿瘤、抗动脉粥样硬化、降血糖及抗衰老等功效[5-6]。据报道,黄芪多糖粉可提高奶牛机体的免疫功能[7],被作为免疫增强剂应用于奶牛养殖[8]。有研究发现发酵黄芪粉可以显著提高奶牛乳蛋白率,提高奶牛血清总抗氧化能力[9-10]。黄芪茎叶生物发酵物可以提高奶牛产奶量和降低乳房炎发病率[11]。刘超等[12]发现黄芪具有调节小鼠高脂血症脂代谢和增强抗脂质过氧化的作用。山楂为蔷薇科植物的干燥成熟果实。现代药理学研究已证实山楂中降血脂的主要活性成分为山楂黄酮和三萜类物质[13],也有研究发现山楂活性成分果胶五糖有降血脂作用[14]。朱光等[15]研究发现山楂善消肉食油腻之积,对降脂具有很好疗效。目前应用山楂对大小鼠研究较为深入,用于反刍动物研究较少。吴志嵩等[16]以山楂山药汤对高脂血症小鼠血脂水平的影响进行研究,发现山楂可降低脂类及胆固醇的浓度。林秋实等[17]发现山楂及山楂黄酮能显著降低大鼠血清总胆固醇,降低低密度脂蛋白-胆固醇,显著升高高密度脂蛋白-胆固醇。在免疫方面,高东雁等[18]以山楂煎剂对大鼠细胞免疫功能有促进作用,减少炎性细胞浸润和肾组织的炎症反应[19]。
为此,本研究通过对产前奶牛日粮中添加山楂和黄芪混合物,探究对奶牛血浆代谢物的变化情况,探讨山楂、黄芪对围产期奶牛的影响因素,为了解山楂、黄芪维持围产期奶牛健康状况提供参考。
1 材料与方法
1.1 材料
1.1.1 试验仪器设备 1290 UHPLC超高效液相(美国Agilent公司)、Triple TOF 6 600高分辨质谱(AB Sciex)、QTOF 6550高分辨质谱(美国Agilent公司)、Heraeus Fresco17离心机(Thermo Fisher Scientific)、BSA124S-CW 天平(Sartorius)、JXFSTPRP-24研磨仪(上海净信科技有限公司)、明澈D24 UV纯水仪(Merck Millipore)、PS-60AL超声仪(深圳市雷德邦电子有限公司)、ACQUITY UPLC BEH Amide 1.7 μm 2.1×100 mm 色谱柱(Waters)。
1.1.2 试验材料与试剂 67-56-1甲醇Methanol(CNW Technologies)、75-05-8 乙腈 Acetonitrile(CNW Technologies)、631-61-8醋 酸 铵 Ammonium acetate(CNW Technologies)、1336-21-6氨 水 Ammonium hydroxide(CNW Technol-ogies)、103616-89-3 L-2-氯苯丙氨酸2-Chloro-L-phenylalanine(上海恒柏生物科技有限公司)。
1.2 方法
1.2.1 试验动物和分组 在宁夏某大型奶牛场,选择20头2-4胎、预产期相近、年龄和体况相近(体况评分BCS3.0-3.5)和上一个产奶周期产奶量相近的健康荷斯坦奶牛。根据预产期(妊娠天数280 d)计算围产期,于产前21±3 d进入围产圈,开始试验。随机分为2组,对照组A组(n=10):对照组饲喂基础日粮,日粮配方见表1。试验组B组(n=10):试验组在基础日粮的基础上添加山楂和黄芪各150 g,每日投料3次(每日6:00、12:00和19:00投料),全混合日粮(Total mixed ration,TMR)饲喂,自由饮水,连续饲喂14 d(试验期间奶牛未产犊牛)。

表1 围产前期奶牛基础日粮组成(干物质基础)
1.2.2 血液的采集保存 于试验第14天晨饲前(5:00-6:00)尾静脉采集血液10 mL,肝素钠抗凝,迅速3 000 r/min离心10 min,收集上层血浆,置于-80℃冰箱冷冻保存,以备后续分析。
1.2.3 检测方法
(1)代谢物提取 取100 μL样本,加入400 μL含有内标的提取液(甲醇乙腈体积比=1∶1,内标浓度 5 μg/mL),涡旋混匀 30 s;超声 5 min(冰水浴);零下20℃静置1 h;将样本4℃,12 000 r/min离心15 min;小心地取出425 μL上清于EP管中;在真空浓缩器中干燥提取物;向干燥后的代谢物加入100 μL提取液(乙腈水体积比:1∶1)复溶;涡旋30秒,冰水浴超声10 min;将样本4℃,12 000 r/min离心15 min;小心地取出60 μL上清于2 mL进样瓶,每个样本各取10 μL混合成QC样本,再取60 μL上机检测。
(2)上机检测 安捷伦1290超高效液相控制下按照表2中的流动相参数进行分析。所使用的色谱柱为购自Waters的UPLC BEH Amide 色谱柱(1.7 μm×2.1×100 mm)。进样体积为 2 μL。
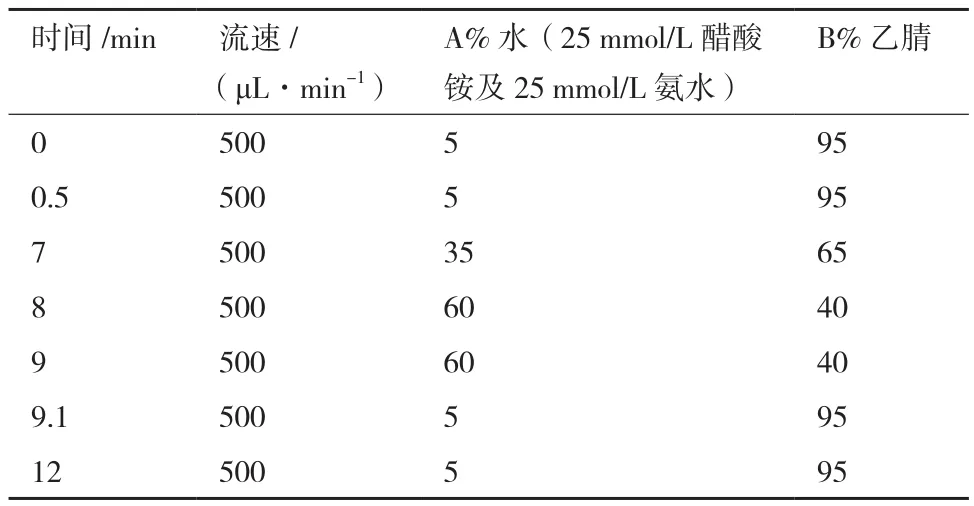
表2 液相色谱流动相条件
AB 6600 Triple TOF & Agilent 6550 QTOF质谱仪能够在控制软件(Analyst TF 1.7,AB Sciex)控制下基于IDA功能进行一级、二级质谱数据采集。在每个数据采集循环中,筛选出强度最强且大于100的分子离子进行采集对应的二级质谱数据。轰击能量:30 eV,15张二级谱图每50 ms。ESI离子源参数设置如下:雾化气压(GS1):60 Psi,辅助气压:60 Psi,气帘气压:35 Psi,温度:650℃,喷雾电压:5 000 V(正离子模式)或-4 000 V(负离子模式)。
1.2.4 数据处理和统计学分析 使用ProteoWizard软件将质谱原始转成mzXmL各式。再使用XCMS做保留时间矫正、峰识别、峰提取、峰积分、峰对齐等工作,minfrac设为0.5,cutoff设为0.6。同时使用自撰写R程序包和自建二级质谱数据库对峰进行物质鉴定。
对数据的完整性进行检查,对缺失值进行删除或者补充,删除极值,并对数据进行样本间和代谢物间的归一化处理,以确保各样本之间和代谢物之间可平行比较。对XCMS提取得到的数据,进行删除组内缺失值>50%的离子峰,不参与后续分析。
对数据进行总峰面积归一化后,使用SIMCA-P软件中对数据进行pareto-scaling处理后,进行多元统计分析:包括主成分分析(Principal component analysis,PCA),正交偏最小二乘判别分析(Orthogonal partial least squares-discriminant analysis,OPLSDA);同时在建模过程中对模型数据进行置换检验并计算变量投影重要度(Variable importance in the projection,VIP)。在此基础上对数据进行单变量统计分析包括:Students t-test 和变异倍数(Fold change,FC)分析,利用R软件根据各代谢物P值和FC值绘制火山图。
1.2.5 差异代谢物鉴定和生物学信息学分析 以VIP>1、P<0.05和 FC>1.3或 FC<0.77作为标准筛选显著性差异代谢物,以VIP>1和0.05<P<0.1作为差异趋势性代谢物,采用质荷比精确匹配(误差<25 ppm)和二级质谱图检索比对本地数据库的方法鉴定差异代谢物。代谢物数据使用MetaboAnalyst平台进行数据auto-scaling(以均值为中心,除以每个变量的标准差)标准化后,进行聚类和KEGG通路分析。
2 结果
2.1 代谢谱分析
由图1可以看到,QC样本TIC出峰保留时间和峰面积都重叠很好,说明仪器稳定性很好。从图2和图3可以看到QC样本密集分布,说明本次实验数据质量很高,并且在正负离子PCA-X一维分布图,可以看出所有QC样本分布于3STD之内,说明数据质量较高。
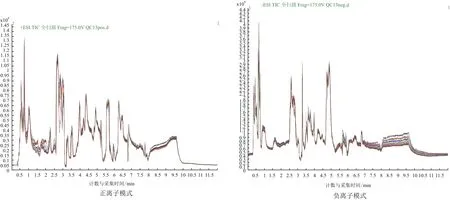
图1 QC样本TIC图
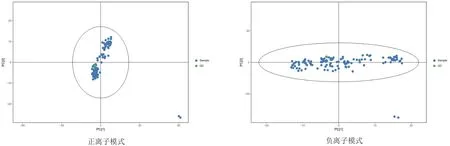
图2 正离子和负离子PCA得分图
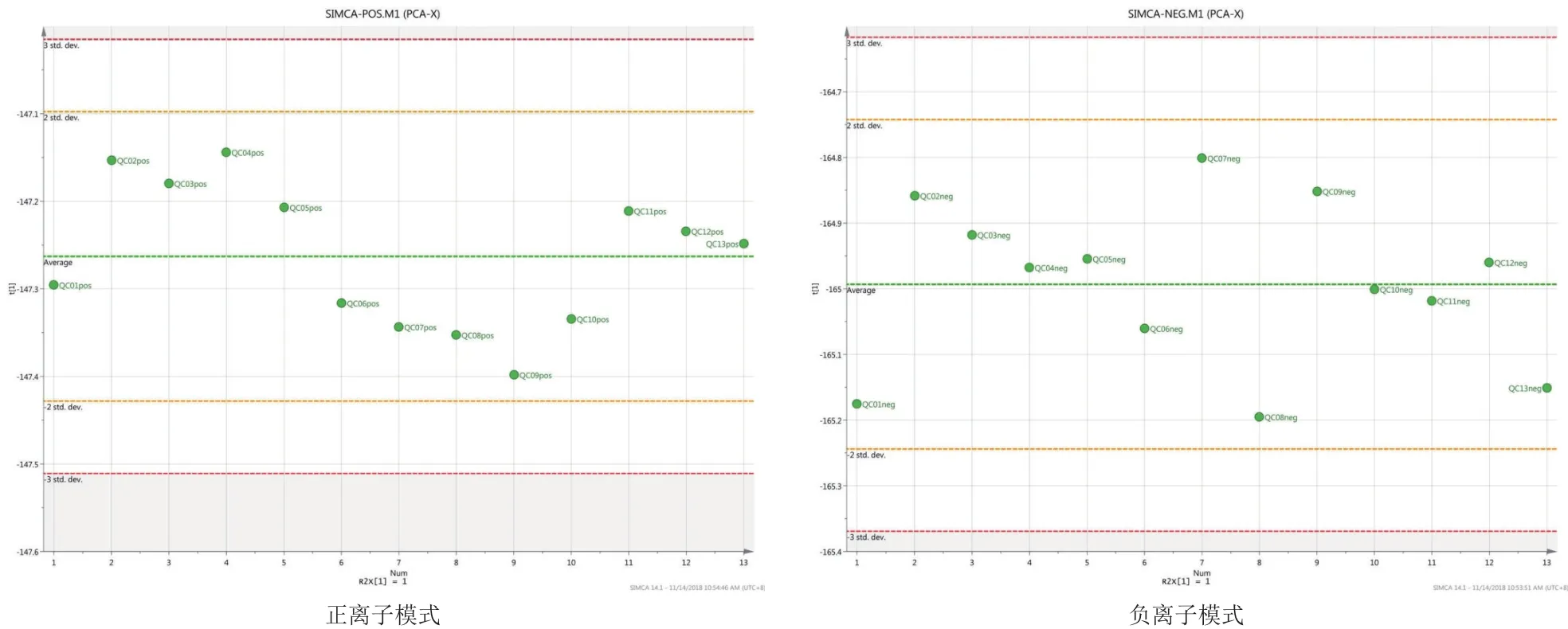
图3 正离子模式和负离子模式下QC样本PCA-X一维分布图
2.2 多元统计学分析
经pareto-scaling处理后,进行模式识别。分析A组和B组用以筛选饲喂山楂、黄芪后两组奶牛的差异代谢物。由图4可知,A与B组数据在负离子模式下有一定分离趋势,正离子模式下R2X=0.554,负离子模式下R2X=0.722。为了进一判定两组是否有着差异,构建有监督模型OPLS-DA。对数据进行滤除和修正得到OPLS-DA模型,模型得分图如图5所示。OPLS-DA模型参数正离子模式下R2Y=1和Q2=0.6,负离子模式下R2Y=0.937和Q2=0.625,R2Y和Q2均大于0.5,说明模型解释率和预测能力高,模型稳定可靠。由图6可知,正离子模式数据置换检验Q2截距 =-0.119(Q2intercept<0),负离子模式Q2截距 =-0.369(Q2intercept<0),说明模型未过度拟合。
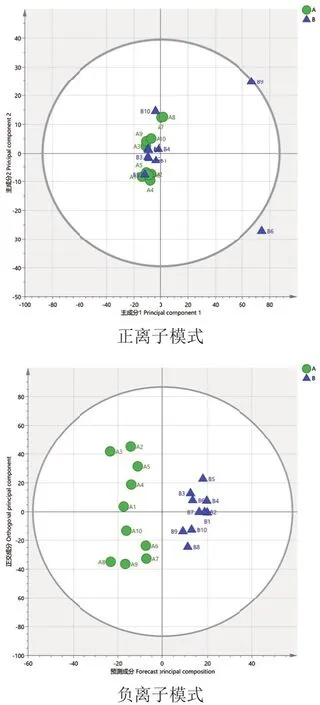
图4 A组和B组在正负离子模式下的PCA得分图
2.3 差异代谢物的筛选
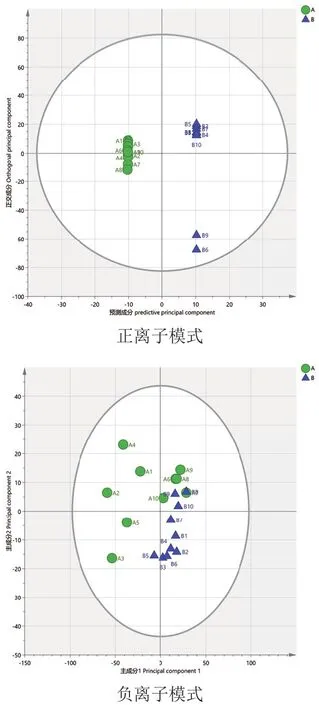
图5 A组和B组两组OPLS-DA得分图

图6 A组和B组两组置换检验图
对预处理后的两组数据进行单变量统计分析,根据t检验中的P值和变异倍数分析中FC值绘制火山图。由图7可知,在正负离子模式下检测的代谢谱呈正态分布,可进行进一步的筛选和鉴定。根据t检验中的P值和OPLS-DA模型获得的VIP值。以 OPLS-DA 模型获得 VIP>1、P<0.05和 FC>1.3或FC<0.77作为显著性差异代谢物筛选条件,以VIP>1和0.05<P<0.1作为差异趋势代谢物。筛选A组和B组的差异代谢物共有60种代谢物,其中正离子模式和负离子模式显著性差异代谢物分别有16和15个,差异趋势代谢物分别有19和10个。正离子模式下显著性差异代谢物2个显著低于对照组,14个显著高于对照组(P<0.05);负离子模式下显著性差异代谢物15个显著低于对照组(P<0.05)。显著性差异代谢物如表3和表4所示。
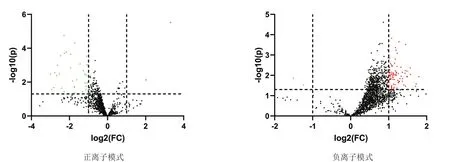
图7 A组和B组代谢物谱火山图表
2.4 生物信息学分析和生物标记物筛选
使用MetaboAnalyst分析平台分别A组和B组筛选出的显著性差异代谢物进行聚类分析和KEGG代谢通路分析。由图8可知,A组和B组中数据有明显的聚集趋势,没能完全聚集在一起可能试验样本组内有个别样本存在差异;由图9中KEGG通路分析结果可知A组和B组显著性差异代谢物参与20种不同代谢通路,其中通路影响力大于0.2为主要代谢通路,包括D-谷氨酰胺和D-谷氨酸代谢、亚油酸代谢、牛磺酸和亚牛磺酸代谢、丙氨酸,天冬氨酸和谷氨酸代谢、缬氨酸,亮氨酸和异亮氨酸的生物合成。对显著性差异代谢物进行单变量ROC曲线分析,进一步考察这些显著性差异代谢物对分类的识别能力,ROC曲线下的面积(AUC)大于0.5时,说明具有较好的识别能力,AUC越接近于1,即ROC曲线越靠近左上角,识别效果越好。本试验结果显示,A组和B组有9个代谢物AUC面积在0.8以上。如图10。这9个代谢物为:Creatinine:肌酐、5,2'-O-dimethylcytidine:5,2'-O-甲基胞苷、Myristoleic acid:肉豆蔻酸、L-Leucine:亮氨酸、Nicotinate:烟酸、3-Hydroxybenzoate:3-羟基苯甲酸、Hypoxanthine:次黄嘌呤、gamma-L-GlutamNicotinateyl-L-valine:γ-谷 氨 酰 缬 氨 酸、Hippuric acid:马尿酸。试验组和对照组相比,9个差异代谢物显著性上调。
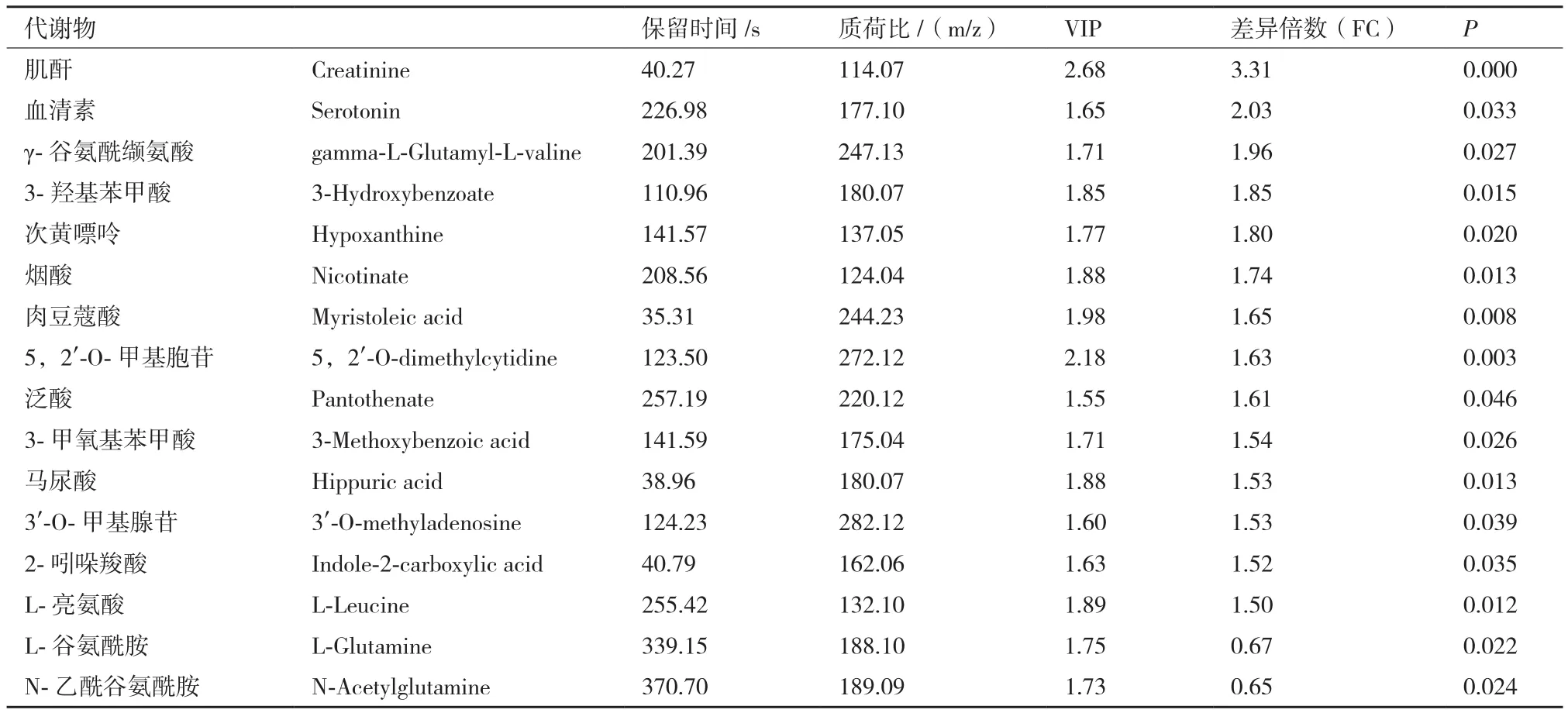
表3 A组和B组正离子模式下鉴定的差异代谢物
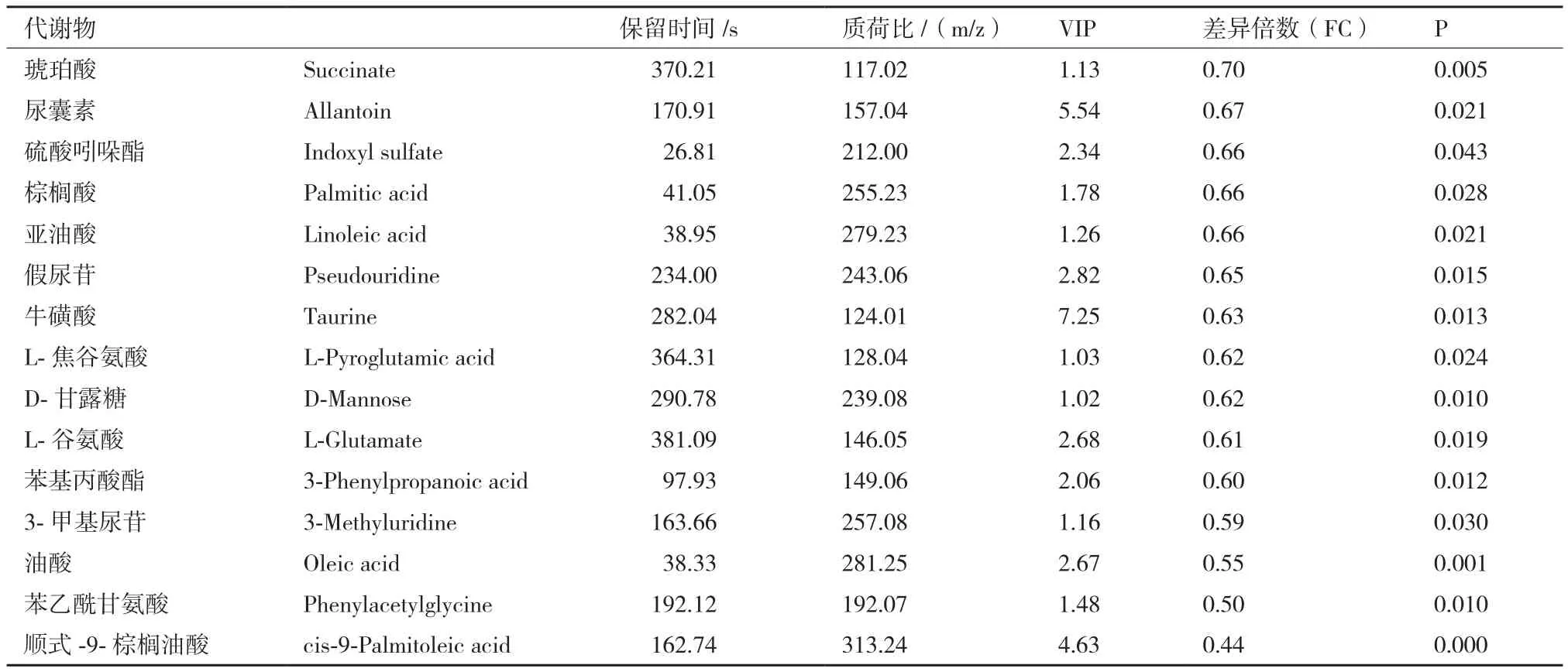
表4 A组和B组负离子模式下鉴定出的差异代谢物
3 讨论
3.1 牛磺酸和亚牛磺酸代谢
牛磺酸和亚牛磺酸代谢通路反映了机体的能量代谢水平[20]。牛磺酸具有维护细胞膜结构的稳定[21],可以促进肌肉组织对葡萄糖和必需氨基酸的摄取和利用,加速糖酵解,增加糖异生[22]。张瑞等[23]在研究黄芪的抗疲劳作用时,发现大鼠腓肠肌中代谢物牛磺酸水平降低。这和本研究中围产前期日粮添加山楂、黄芪后饲喂14 d试验组奶牛血浆代谢物牛磺酸显著下调试验结果一致。说明日粮添加山楂、黄芪可能通过加速奶牛血浆代谢物牛磺酸的代谢,用于调控葡萄糖和氨基酸的代谢。
3.2 D-谷氨酰胺和D-谷氨酸代谢及丙氨酸,天冬氨酸和谷氨酸代谢
谷氨酰胺属于生糖氨基酸被作为生糖物质,是肝糖原发生异生作用的重要底物,维持和调节生物机体的总氨基酸稳态[24]。谷氨酰胺参与调控葡萄糖代谢过程[25],增强免疫效应[26]。谷氨酰胺和谷氨酸同时参与D-谷氨酰胺和D-谷氨酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢两个代谢通路。D-谷氨酰胺和D-谷氨酸代谢通路中谷氨酰胺转化为谷氨酸,丙氨酸、天冬氨酸和谷氨酸代谢通路中谷氨酸在谷氨酰胺合成酶的作用下合成谷氨酰胺,谷氨酰胺被谷氨酰胺磷酸酶水解成谷氨酸,继续参与循环[27-28]。谷氨酸参与D-谷氨酰胺和D-谷氨酸代谢通路时,在谷氨酸脱氢酶的作用下被代谢为α-酮戊二酸。参与丙氨酸、天冬氨酸和谷氨酸代谢通路时,谷氨酸可生成琥珀酸[29]。琥珀酸是三羧酸循环的中间产物,在三羧酸循环中为机体提供能量[30]。有研究表明谷氨酸是中枢神经系统中主要的兴奋性神经递质[31],可以作为中间代谢产物将氨基酸代谢和蛋白质代谢连接起来[32]。本研究中在围产前期日粮添加山楂、黄芪试验组围产期奶牛血浆代谢物谷氨酰胺和谷氨酸分别显著上调和下调。造成这种结果可能是因为谷氨酸转化成谷氨酰胺增加或谷氨酰胺转化为谷氨酸减少,增多谷氨酰胺可能用于调控葡萄糖代谢过程,提高机体蛋白质的合成和葡萄糖能量的补充。
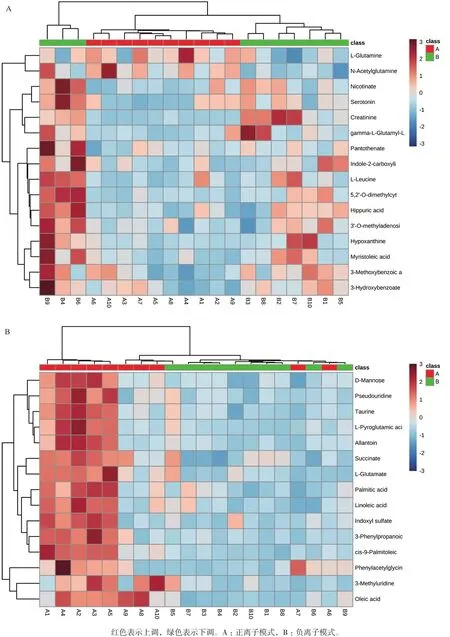
图8 A组和B组差异代谢物聚类热图
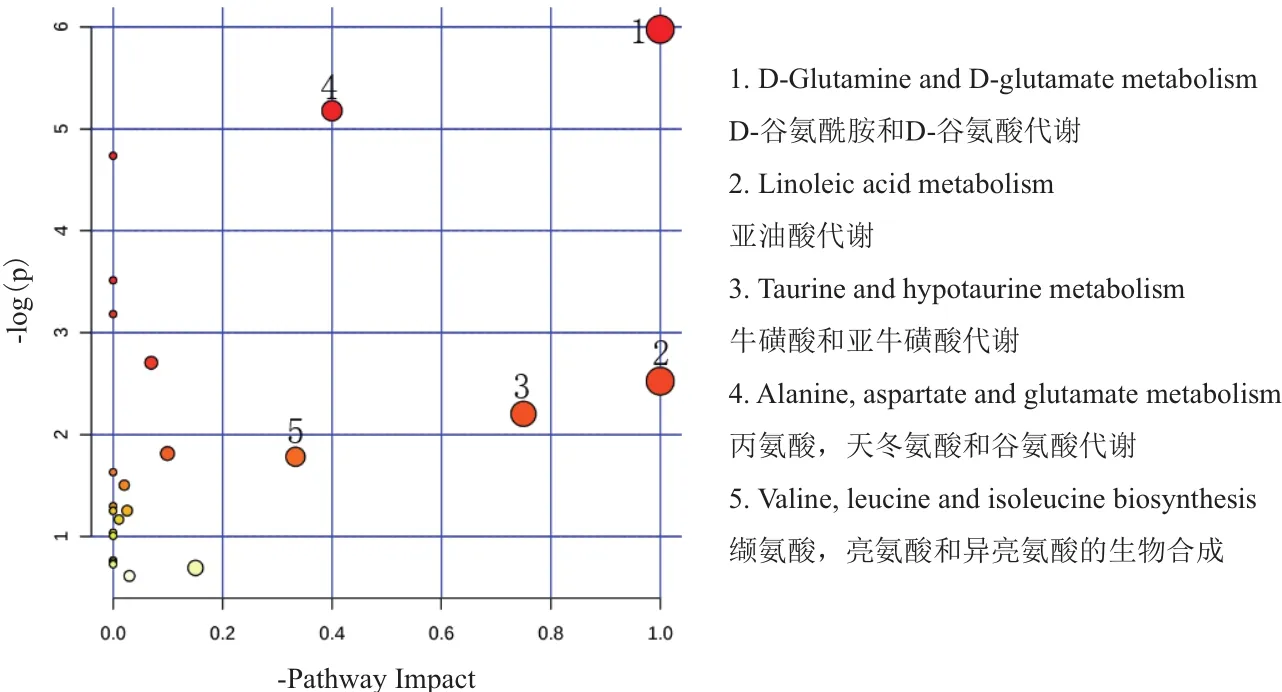
图9 A组和B组差异代谢物的代谢通路分析
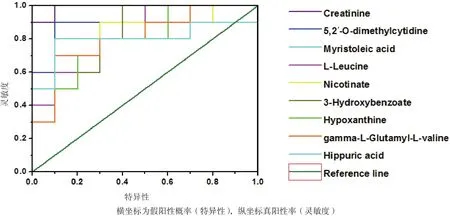
图10 十个显著性差异代谢物的ROC曲线
3.3 缬氨酸,亮氨酸和异亮氨酸的生物合成
亮氨酸是机体必需氨基酸中的支链氨基酸,也是重要的生酮氨基酸[33-34]。Arakawa和 Pleck 等[35-36]研究发现亮氨酸在调控糖脂代谢和免疫功能方面有重要作用。曲婷丽等和王二兵等[37-38]研究发现黄芪乙酸乙酯萃取物治疗小鼠白细胞减少症时血清代谢物亮氨酸显著上调。这和本研究血浆代谢物亮氨酸显著上调的结果一致。在围产前期奶牛连续饲喂14 d山楂、黄芪血浆代谢物亮氨酸上调,说明日粮添加山楂、黄芪有利于增强产前期奶牛免疫力,维持免疫系统稳定。
3.4 亚油酸代谢
亚油酸是组成游离脂肪酸的重要成分[39]。有研究发现亚油酸能诱导机体产生大量活性氧物质,引起机体红细胞的急性损伤[40]。李秋红等[41]发现荷叶黄酮具有良好的清除自由基能力,能有效抑制亚油酸的氧化。有研究认为,部分黄酮类化合物具有抗氧化作用[42],胡博路等[43]研究发现黄芪具有很强的抗氧化活性。由此推测黄芪中黄酮类化合物可能是抑制亚油酸氧化的物质,减少亚油酸氧化导致奶牛机体细胞损伤。有研究表明亚油酸对奶牛乳腺上皮细胞的生长有抑制作用,并且对奶牛乳腺上皮细胞液中脂质小滴的合成有促进作用[44]。本研究中在围产前期奶牛连续饲喂14 d山楂、黄芪血浆代谢物亚油酸显著下调,说明产前奶牛日粮中添加山楂、黄芪有利于乳腺上皮细胞生长及促进脂质小滴合成的作用。
4 结论
本研究中在围产前期奶牛连续饲喂14 d山楂、黄芪血浆代谢物中谷氨酰胺和亮氨酸显著上调,琥珀酸、牛磺酸、谷氨酸和亚油酸显著下调,这些物质调节围产期奶牛葡萄糖代谢和氨基酸代谢通路中牛磺酸和亚牛磺酸代谢、D-谷氨酰胺和D-谷氨酸代谢及丙氨酸、天冬氨酸和谷氨酸代谢和缬氨酸,亮氨酸和异亮氨酸的生物合成,及脂肪酸代谢通路中亚油酸代谢。
猜你喜欢
电脑报(2022年25期)2022-07-05
食品与生物技术学报(2021年4期)2021-01-17
食品与健康(2020年7期)2020-07-09
今日畜牧兽医(2018年6期)2018-02-13
中成药(2017年6期)2017-06-13
中国洗涤用品工业(2017年2期)2017-04-16
中国调味品(2017年2期)2017-03-20
中国比较医学杂志(2017年5期)2017-01-17
中外医疗(2015年11期)2016-01-04
医学研究杂志(2015年12期)2015-06-10
