
牦牛Linc24063的克隆鉴定及其与miRNAs表达水平的相关性分析
2019-08-08 08:06王会柴志欣朱江江钟金城张成福信金伟
中国农业科学 2019年14期
王会 ,柴志欣 ,朱江江 ,钟金城 ,张成福 ,信金伟
(1西南民族大学,青藏高原动物遗传资源保护与利用教育部重点实验室,成都 610041;2西南民族大学,青藏高原动物遗传资源保护与利用四川省重点实验室,成都 610041;3西藏自治区农牧科学院, 省部共建青稞和牦牛种质资源与遗传改良国家重点实验室,拉萨 850002)
0 引言
【研究意义】在机体内,lncRNA可通过miRNA发挥调控作用。一方面,lncRNA可作为miRNA吸附剂,与miRNA靶基因竞争性结合miRNA;另一方面,lncRNA可作为竞争性内源RNA(ceRNA),与具有相同miRNA结合元件的mRNA竞争miRNA,抑制翻译、调节可变剪切或调节 mRNA降解[1]。牦牛主要分布于海拔3 000 m以上的青藏高原地区,为藏区人民提供肉、奶产品。牦牛奶具有高蛋白、高乳脂率的特点,研究 lncRNA在牦牛乳腺组织中的具体功能及调控机制,对更好地利用牦牛遗传资源具有重要意义。【前人研究进展】在牦牛等哺乳动物中,由于早期配子遗传的精子和卵子的表观遗传修饰不同,导致接近1%的蛋白质编码基因表达具有亲本特异性,这种亲本依赖的表观遗传现象被定义为基因组印记[2-3]。迄今为止,在人类、小鼠、牛和绵羊等哺乳动物中分别鉴定出350个、236个、45个和18个印记基因(http://igc.otago.ac.nz),这些印记基因通常以簇的形式存在。Dlk1-Dio3印记域位于人染色体14q32、小鼠染色体12q、牛染色体 21q和绵羊染色体18q上,因其在胚胎发育[4]、癌症的发生[5]、细胞增殖分化[6]及机体各代谢调控通路[7]中发挥重要作用而备受关注,是目前研究较多的印记区域之一。Dlk1- Dio3印记域包括3个父源表达的编码蛋白基因(Dlk1、Dio3和 Rtl1/Peg11)与母系表达的长链非编码 RNA(如lncRNA:Meg3、Meg8、Meg9)、miRNA 和 snoRNA[8]。研究表明,Dlk1-Dio3印记域在物种间高度保守,广泛参与到机体各生理及病理过程中。Dlk1-Dio3印记区域的激活与小鼠干细胞多能性水平相关[9];Dlk1参与调控多能干细胞[10]、肌细胞的增殖与分化[7];Dlk1启动子区域甲基化可增加非小细胞肺癌细胞的侵袭能力[11]。LncRNA是长度大于200 bp的一类非编码RNA,在胚胎发育[12]、细胞增殖分化[13]、精子发生[14]等多种生命过程中发挥重要的调控作用。Dlk1-Dio3印记域上有多个具备重要功能的 lncRNA,如高脂饮食可增加小鼠Meg3表达量,Meg3通过调控FoxO1的表达增强肝脏胰岛素抵抗,干扰Met3可逆转小鼠由高脂饮食引起的血液甘油三酯含量的上调[14]。近年来,人们在不同物种Dlk1-Dio3印记域间发现并鉴定到了新lncRNA,如在鼠Meg8与Meg9、Meg9与Dio3之间分别发现并鉴定了2个lncRNA,即lncRNAB830012L 14Rik和AK044800,lncRNAB830012L 14Rik在小鼠大脑、肺脏、心脏和肝脏组织中广泛表达[15],而 lncRNAAK044800主要在胚胎时期的前脑组织中表达[16];对牛Dlk1-Dio3印记域lncRNA鉴定发现,该区域包含Linc24061、Linc24063、Linc24064三个 lncRNAs,它们在在心脏、肾脏及肌肉组织中均广泛表达[17-18]。【本研究切入点】前人研究表明,Dlk1-Dio3印记域内的lncRNA,能参与机体多种生物学过程调控。但在牦牛上,未见此印记域内 lncRNA的研究报道。【拟解决的关键问题】本研究旨在牦牛Dlk1- Dio3印记域Meg8与Meg9间鉴定牦牛Linc24063,通过生物信息学对其靶miRNA进行预测,结合组织表达谱分析其表达量在乳腺组织中与靶 miRNAs的相关性,从而为牦牛Linc24063通过miRNA发挥功能的调控机制研究提供试验依据。
1 材料与方法
1.1 试验材料
2018年4月,在西藏自治区昌都市类乌齐县,选取处于第一泌乳期、体重接近、健康的12头母牦牛,屠宰后采集卵巢、心脏、肝脏、肾脏、乳腺和大脑组织,DEPC反复冲洗后锡箔纸包装,迅速置于液氮保存,带回实验室备用。
1.2 RNA提取及牦牛Linc24063扩增
各组织RNA提取参照Trizol(Invitrogen)试剂说明书。RNA经纯度和质量检测合格后备用。
利用牛Dlk1-Dio3印记域Meg8与Meg9间的序列进行搜索并与牦牛基因组比对,鉴别出牛Linc24063(NCBI登录号:KU956000.1)序列与牦牛基因组序列高度保守,根据此序列利用 Primer premier 5设计牦牛Linc24063基因5′ RACE(cDNA末端快速扩增)和3′ RACE引物。将牦牛各组织RNA等量混合后,利用SMARTer RACE cDNA Amplification Kit(Clontech, Palo Alto, CA, USA)按照操作说明书进行 RACE试验,获取Linc24063的 5′端和 3′端。5′RACE 引物为:5′-TTAAACATTCCTAACATCTGCC TAC-3′,3′ RACE 引物为 5′-GGGACCCTGAGGCCC AGTCCATTC-3′。
1.3 牦牛Linc24063序列分析
利用NCBI BLAST对牦牛Linc24063序列在物种间的保守性及染色体上的定位进行分析;利用在线软件Coding Potential Calculator(http://cpc2.cbi.pku.edu.cn/)对Linc24063、已知 lncRNAMeg9(登录号:NR_132275.2)和编码基因β-酪蛋白(CSN2,登录号:MH378280.1)的编码能力进行预测;利用 miRanda和mireap软件分析与Linc24063相互作用的miRNAs;利用 DAVID在线软件(https://david.ncifcrf.gov/)对与Linc24063相互作用的miRNAs的靶基因进行GO富集和KEGG信号通路分析。
1.4 牦牛Linc24063原核表达
将牦牛Linc24063全长亚克隆至 Pet-28a载体的EcoRⅠ和HindⅢ酶切位点上,获得Linc24063-28a原核表达质粒,转化至BL21感受态细胞,获得含重组质粒Linc24063-28a表达菌种,提取质粒用EcoRⅠ和HindⅢ酶切鉴定。CSN2-28a原核表达质粒为本实验室保存。
Linc24063-28a和CSN2-28a诱导表达步骤参照文献[19]。取 1 mL诱导后的菌液沉淀,用 100 μL的1×SDS-PAGE Loading Buffer重悬沉淀,沸水中煮沸10 min,保证菌体完全裂解。10 000×g离心5 min后,取上清10 μL用于SDS-PAGE分析。凝胶用考马斯亮蓝R-250染色液染色,脱色液脱色,凝胶成像。
1.5 牦牛Linc24063组织表达分析
利用PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa, 大连)试剂盒将各组织总RNA反转录为cDNA,Linc24063表达量的检测参照SYBR Premix Ex TaqTM试剂盒(Takara)说明书,内参基因选择核糖体蛋白 S9(RPS9,登录号:XM_005899362.2)、广泛表达蛋白(UXT, 登录号:XM_014483477.1)[20];对于miRNA表达量的检测利用Stem-loop方法[21],内参基因选择5S rRNA。利用Bio-Rad CFX96(伯乐,美国)定量仪进行牦牛Linc24063及 miRNAs组织表达谱分析。扩增体系均为:2×SYBR®Premix Ex TaqTMⅡ10 μL,cDNA 模板 1 μL,上下游引物各 1 μL,ddH2O 7 μL。qPCR 条件为:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,39个循环。所用引物列于表1。
1.6 数据分析
基因表达量差异分析均使用 2-△△ct法,对于Linc24063的定量选择UXT和RPS9的几何平均值进行分析。利用 SPSS 19.0皮尔逊系数(双尾)反应Linc24063与miRNA的相关性。相关性以|R |≥0.8为极强相关,0.6≤|R |≤0.8为强相关,0.4≤|R |≤0.6为中度相关,|R |≤0.4为不相关。结果表示为平均值+标准误。*,P<0.05为差异显著,**,P<0.01为差异极显著。

表1 实时荧光定量PCR引物Table1 Specific primers used for RT-qPCR
2 结果
2.1 长链非编码RNA Linc24063序列特征
利用SMARTer RACE cDNA Amplification Kit 扩增牦牛长链非编码 RNALinc24063,结果表明,5′RACE片段大小为476 bp,3′ RACE片段大小为356 bp(图1-A),测序分析表明Linc24063大小758 bp(图1-B),位于牦牛21号染色体的Dlk1-Dio3印记域。NCBI blast分析物种间保守性,结果表明牦牛Linc24063与牛、绵羊、山羊的序列相似性分别为98%、90%和90%,而与人和小鼠的序列相似性较低。
2.2 长链非编码RNA Linc24063的鉴定
利用在线软件Coding Potential Calculator对长链非编码RNA Linc24063、已知lncRNAMeg9和编码基因CSN2的编码能力预测分析,结果表明:Linc24063与长链非编码RNAMeg9类似,编码潜能均较低,明显区别于编码基因CSN2,属于非编码RNA(表2)。
为验证Linc24063是否具备编码能力,本研究将Linc24063连接pET-28a原核表达载体,提取质粒进行EcoRⅠ和HindⅢ双酶切鉴定,由图2-A可知,Linc24063-28a和CSN2-28a分别检测到一条758 bp和 780 bp大小的片段,进一步通过测序验证目的片段成功插入原核表达载体。利用原核表达系统在体外对Linc24063和编码基因CSN2进行翻译,结果表明:Linc24063在30 kD附近没有检测到蛋白表达,表明其不能有效的翻译蛋白,而在相同试验条件下,编码基因CSN2在30 kD附近有蛋白表达,表明CSN2能很好的表达蛋白(图2-B)。以上研究结果表明,牦牛Linc24063是一个真正的长链非编码lncRNA。

图1 Linc24063 5′ Race和3′ Race电泳图(A)及序列(B)Fig.1 The 5′ Race and 3′ Race electrophoresis analysis (A) and the sequence (B) of Linc24063
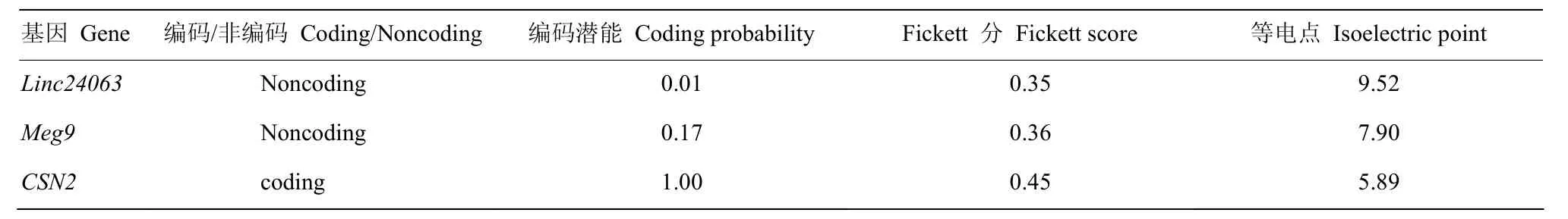
表2 Linc24063、Meg9和CSN2基因编码潜能分析Table2 The coding probability analysis of Linc24063, Meg9 and CSN2

图2 Linc24063的原核表达Fig.2 The prokaryotic expression of Linc24063
2.3 长链非编码RNA Linc24063组织表达谱
利用RT-qPCR技术,笔者研究了Linc24063在牦牛不同组织的表达谱。结果显示,Linc24063在乳腺表达量最高,显著高于其他组织(P<0.05),其次大脑和肾脏,在肝脏和卵巢中表达量较低(图3)。

图3 Linc24063在各组织中的表达量Fig.3 The expression of Linc24063 in tissues
2.4 长链非编码RNA Linc24063生物信息学分析
下载miRbase 22数据库中普通牛(牦牛的近源物种)和绵羊所有的miRNAs,利用miRanda和mireap软件分析与Linc24063具有相互作用的miRNA,共发现21个物种间保守miRNAs(表3)。
由2.3研究表明,Linc24063在乳腺的表达量最高,我们推测其在乳腺发育或乳合成过程中具有重要作用,由此结合前人在乳腺中miRNAs表达谱的研究[22-23],发现13个miRNAs可能在乳腺组织中与Linc24063具有相互作用。对这13个miRNAs的靶基因进行GO富集和KEGG信号通路分析,结果表明,这些miRNAs显著富集到RNA聚合酶II相关基因的转录等生物学过程中(表4)。KEGG信号通路分析表明,这些miRNAs显著参与到TGF-beta信号通路、PI3K-Akt信号通路、ErbB信号通路、MAPK信号通路等(表5)。
2.5 长链非编码RNA Linc24063与miRNAs表达相关性分析
为探究Linc24063与miRNAs的功能相关性,本研究利用RT-qPCR在乳腺组织中检测Linc24063与 miR-200a 、miR-24、miR-141及 miR-27a的表达量,并进行了皮尔逊相关性分析(图4),结果表明:Linc24063在12头牦牛乳腺组织中的表达量与 miR-200a(R=-0.834,P=0.001)和 miR-141(R=-0.657,P=0.02)的表达量均显著负相关,与miR-27a的表达量显著正相关(R=0.647,P=0.023),而与miR-24无显著相关性(P>0.05)。以上研究结果表明,Linc24063在牦牛乳腺组织中可能通过调控miR-200a、miR-141和miR-27a基因的表达,从而参与到乳腺发育或乳成分合成及代谢的生理过程中。

表3 与Linc24063相互作用的保守miRNAsTable3 The conserved miRNAs interacting with Linc24063

表4 与Linc24063相互作用的miRNAs靶基因的前10条GO富集条目Table4 The top ten GO terms of the conserved miRNAs interacting with Linc24063

图4 12头牦牛乳腺组织Linc24063与miRNAs表达量相关性分析Fig.4 The correlation between expression levels of Linc24063 and miRNAs in 12 mammary gland of yaks
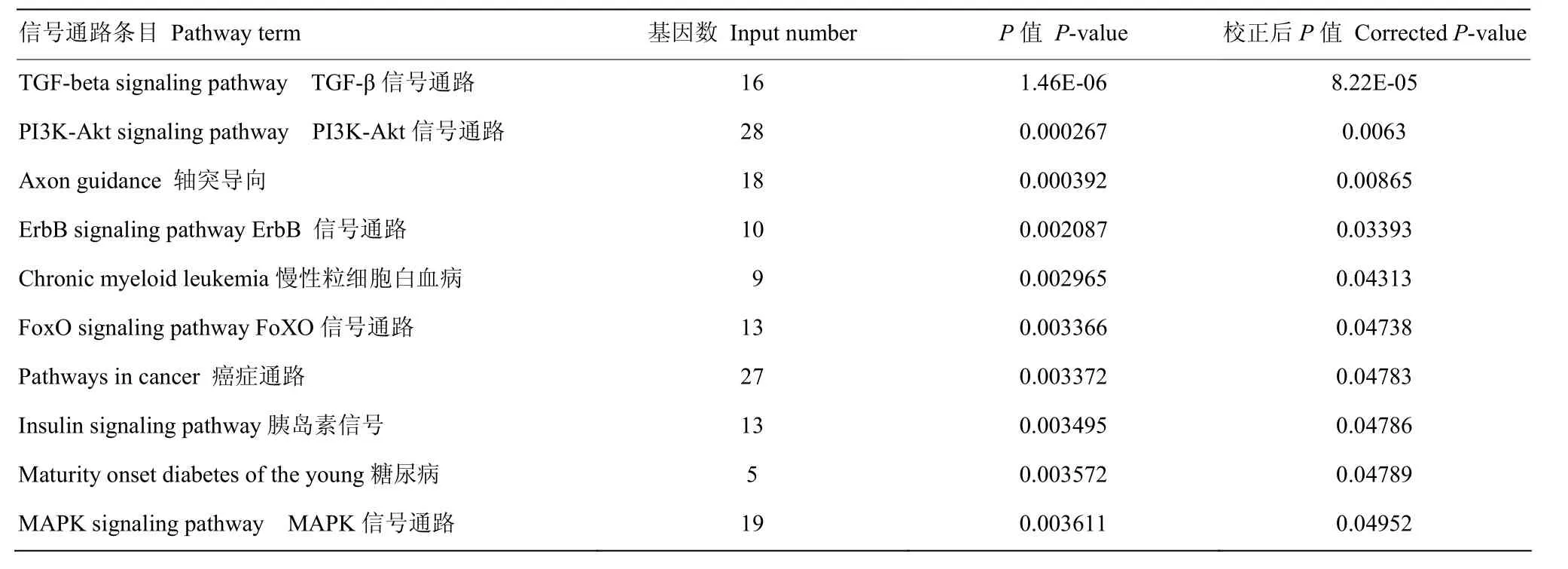
表5 与Linc24063相互作用的miRNAs靶基因的信号通路Table5 The KEGG pathway of the conserved miRNAs interacting with Linc24063
3 讨论
目前,关于lncRNA的研究不仅局限于模式动物及人上,在猪、牛、羊上也开展了大量研究,但是关于牦牛lncRNA的研究较少。本研究首次通过RACE技术获取并鉴定了牦牛Linc24063,位于牦牛21号染色体的Dlk1-Dio3印记域,在反刍动物牛、羊中具有较高的保守性(>90%),与前人对Dlk1-Dio3印记域的研究表明该印记域在物种间具有较强的保守性的结果一致[24]。组织表达谱分析结果表明,Linc24063在所检测的6个组织中均有表达,且在乳腺中表达量最高,其次是大脑和肾脏,表明Linc24063在这些组织中可能均具备生物学功能。
生物信息学分析结果表明:共筛选到 21个与Linc24063相互作用的物种间保守miRNAs 。结合前人研究对miRNAs在乳腺组织中的表达谱研究[22-23],笔者筛选了 13个 miRNAs可能在乳腺组织中与Linc24063具有相互作用。对这13个miRNAs的靶基因进行 GO富集和 KEGG信号通路分析表明:与Linc24063相互作用的 miRNAs的靶基因富集到TGF-beta信号通路、PI3K-Akt信号通路、胰岛素信号通路及与神经发育、保护的轴突引导(Axon guidance)信号通路中。TGF-beta信号通路[25]、PI3K-Akt信号通路[26-27]、胰岛素信号通路是与乳腺发育、乳脂乳蛋白合成密切相关的信号通路,结合Linc24063在乳腺组织中表达量最高的特点,推测Linc24063可能在乳腺发育及乳脂乳蛋白合成等生物学过程中发挥重要作用。
为探索Linc24063可能的作用机制,本研究随机挑选了4个可能与其具有相互作用的miRNAs,在乳腺组织中对其表达量进行了相关性分析。结果表明,Linc24063与 miR-200a、miR-141、miR-27a均显著相关。前人研究表明miR-200a在乳腺上皮细胞中通过调控β-酪蛋白和E-钙粘蛋白以维持上皮细胞的极性[28],同时在乳脂、乳蛋白合成信号通路中发挥重要功能[29];miR-141和 miR-27a可分别通过调控STAT5蛋白、PPARγ蛋白的表达,参与到乳蛋白、乳脂的合成中[30]。由此可以推测,Linc24063可能通过调控miR-200a、miR-141、miR-27a参与到牦牛乳蛋白及乳脂的生物合成中,但是其具体功能和作用机制还需进行研究。
4 结论
牦牛Dlk1-Dio3印记域存在一个在反刍动物中保守的长链非编码RNALinc24063,全长758 bp。在乳腺及大脑组织中具有较高表达水平,生物信息学分析表明Linc24063可能参与到PI3K-Akt、胰岛素等与乳脂乳蛋白密切相关的信号通路中。在乳腺组织中Linc24063与乳脂乳蛋白合成相关的 miRNAs,如 miR-200a、miR-27a和miR-141具有显著的相关性。以上研究结果为Linc24063的功能研究提供了基础数据。
猜你喜欢
合成技术及应用(2022年2期)2023-01-02
青海湖(2022年3期)2022-06-09
现代苏州(2022年9期)2022-05-26
滇池(2022年5期)2022-04-30
散文诗(2021年22期)2022-01-12
昆明医科大学学报(2021年4期)2021-07-23
铁路通信信号工程技术(2021年3期)2021-04-03
小哥白尼(野生动物)(2020年9期)2021-01-18
人大建设(2020年5期)2020-09-25
红岩春秋(2019年5期)2019-05-31
