
长江中下游稻区不同水旱轮作模式和氮肥水平对稻田CH4排放的影响
2019-08-08 08:06刘少文殷敏褚光徐春梅王丹英章秀福陈松
中国农业科学 2019年14期
刘少文,殷敏,褚光,徐春梅,王丹英,章秀福,陈松
(中国水稻研究所/水稻生物学国家重点实验室,杭州 311400)
0 引言
【研究意义】全球气候变暖正日益影响着人类的生存环境和质量[1],而其发生与温室气体浓度的增加有关[2]。CH4作为主要的温室气体之一,其增温效应约占全部温室气体总效应的 15%—20%,且每年以1.1%的速度增长[3]。研究发现农田是大气CH4的重要排放源,约17.9%的CH4排放来源于稻田[4]。水稻是中国重要的粮食作物,其种植面积约占世界水稻种植总面积的 23%[5]。深入研究稻田 CH4排放特点及其影响因素,并在此基础上制定相应的减排措施,对缓解全球增温效应意义重大。【前人研究进展】稻田CH4的产生是由产甲烷菌将土壤中有机物质通过厌氧发酵转化而成[6],CH4从厌氧沉积层或淹水土壤向大气传输主要经由分子扩散、气泡迸发和通过植株排放[7],其中超过90%的稻田CH4通过水稻植株排放到大气[8]。对稻田CH4排放及相应的减排措施已有大量研究。气候条件、土壤特性、耕作方式以及田间管理是影响稻田CH4排放的重要因素[9-11]。一般认为水旱轮作能够改善土壤的理化性质,降低土壤还原物质含量,减少 CH4的产生[12];而稻田持续淹水,会阻碍氧气的扩散,降低土壤氧化还原电位,易于产甲烷菌的快速繁殖,导致 CH4排放的增加[13];中期烤田则能减少36%—65%以上的CH4排放[14-15]。此外,研究表明,秸秆添加能显著增加稻田 CH4排放量,且排放总量和秸秆还田量呈正相关[16]。在生产上,水旱轮作过程中往往伴随着冬季作物秸秆全量还田,长江中下游地区主要的水旱轮作包括稻麦、稻油、稻绿、稻闲等模式[17-18],而近年来,秸秆覆盖种植马铃薯也被广泛推广[19]。合理的秸秆还田能增加土壤有机质含量,培肥地力、改善土壤结构[20],但不同属性的秸秆还田对温室气体排放的影响,目前尚未见系统报道。相关研究表明,田间氮肥施用对稻田 CH4排放亦有显著影响,但目前尚未有一致的结论,有研究认为氮肥会增加稻田 CH4排放[21-22],也有研究发现随着施氮量的增加,稻田 CH4排放减少[23]。由于氮肥会影响还田秸秆的分解及其 CH4转化过程[24],因此氮肥对 CH4排放的影响可能还与稻田的秸秆类型有关。【本研究切入点】目前关于水旱轮作模式(不同属性秸秆还田处理)与氮肥水平对稻田 CH4的排放特性的互作效应研究报道较少,而对其相关排放机理的研究则尚未见报道。【拟解决的关键问题】鉴此,本研究以多年长期定位试验(2003至今)为基础,选择 4种不同水旱轮作模式(不同属性秸秆还田处理),结合3种不同氮肥水平,通过田间气体采集与土壤样品分析,主要目的在于:(1)评估还田秸秆属性与氮肥水平互作对稻季 CH4的排放规律及累积排放量的影响;(2)结合土壤甲烷代谢微生物、土壤理化性质与秸秆属性综合分析,明确不同秸秆还田与氮肥处理下稻季CH4排放差异机理。以期为我国稻田温室气体排放以及合理减排措施的制定提供科学依据。
1 材料与方法
1.1 试验区概况
试验于2016—2017年,在浙江省富阳市中国水稻研究所农场进行(120.2°E,30.3°N,海拔11 m)。该农场位于长江中下游平原,属亚热带季风气候,年平均气温为13—20℃,其中7月份为35℃;年平均降水量为1200—1600 mm,其中4月份和9月份降雨量约占80%。本研究所需的稻田种植模式定位试验于2003年开始实施,于 2016—2017年测定稻田CH4排放,两年土壤基本性质年度间差异不显著,具体指标见表1。
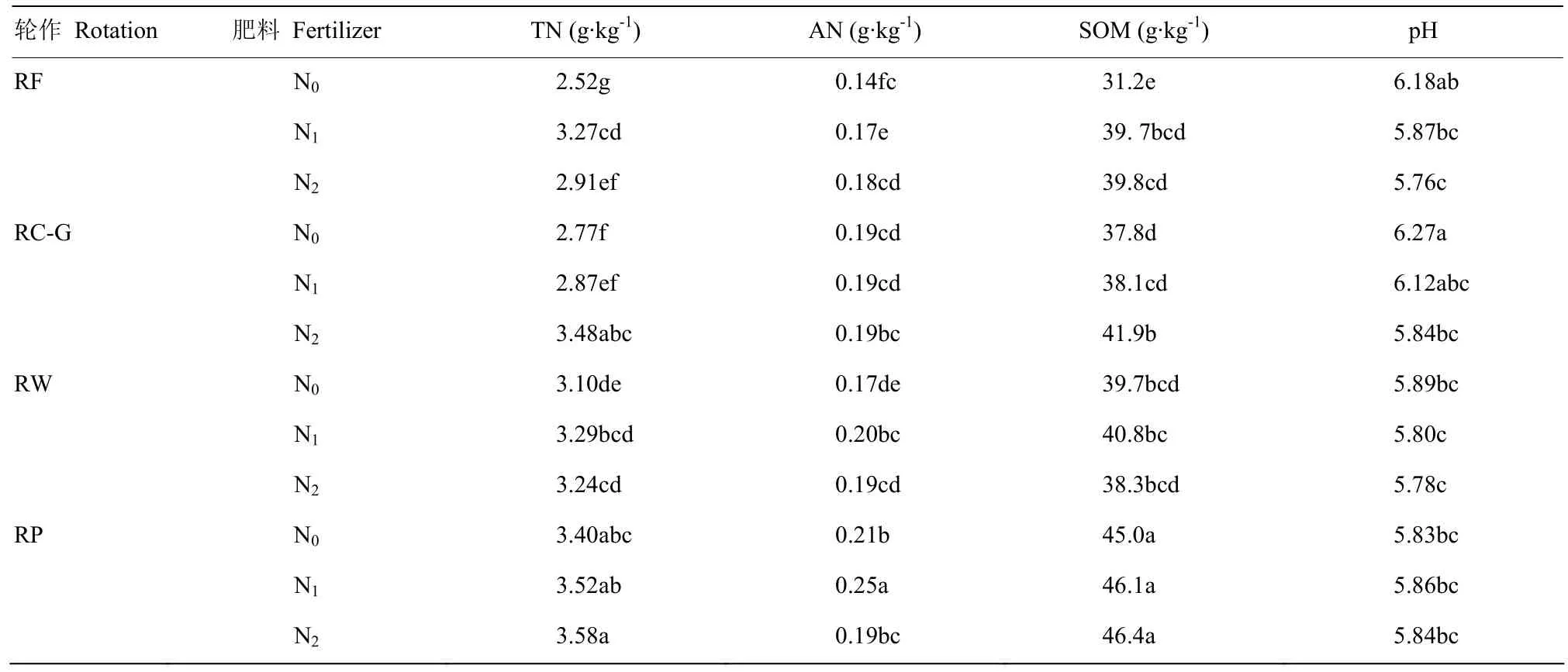
表1 不同轮作及氮肥水平下土壤基本性质(两年平均)Table1 Basic properties of soil under different rotation systems and different N rates(the two years’ average)
1.2 试验设计
本研究以不同水旱轮作模式长期定位试验为基础(2003至今)。于2016—2017年测定稻田CH4排放。定位试验采用裂区随机区组设计,4种不同轮作模式为主区,分别为水稻-休闲(RF),水稻-紫云英(RC-G),水稻-小麦(RW)和水稻-稻草覆盖种植马铃薯(RP);水稻季氮肥水平为副区,分别为N0(0)、N1(142.5 kg N·hm-2)和 N2(202.5 kg N·hm-2),小区位置和面积长期固定,每个小区长8 m、宽8 m,3次田间区组重复。
冬季作物栽培:RF,田间不作任何处理,空闲放置,次年水稻移栽前,翻耕耘田;RC-G,紫云英于10月中下旬播种,出苗后施入尿素15 kg·hm-2作为基肥,次年盛花期(3月底4月初)将其割翻,翻耕还田;RW,小麦于10月下旬11月初撒直播,苗期和抽穗期分别施入复合肥75 kg·hm-2(基肥)和尿素15 kg·hm-2(追肥),次年5月中旬收获,除籽粒外,其余秸秆翻耕还田;RP,稻草全程覆盖种植马铃薯,于1月下旬在田面上直接摆放种薯,并覆盖8—10 cm厚稻草,施入复合肥75 kg·hm-2作为基肥,5月中旬收马铃薯块茎,其余部分翻耕还田,还田的有机物为腐烂的稻草和马铃薯茎叶等。
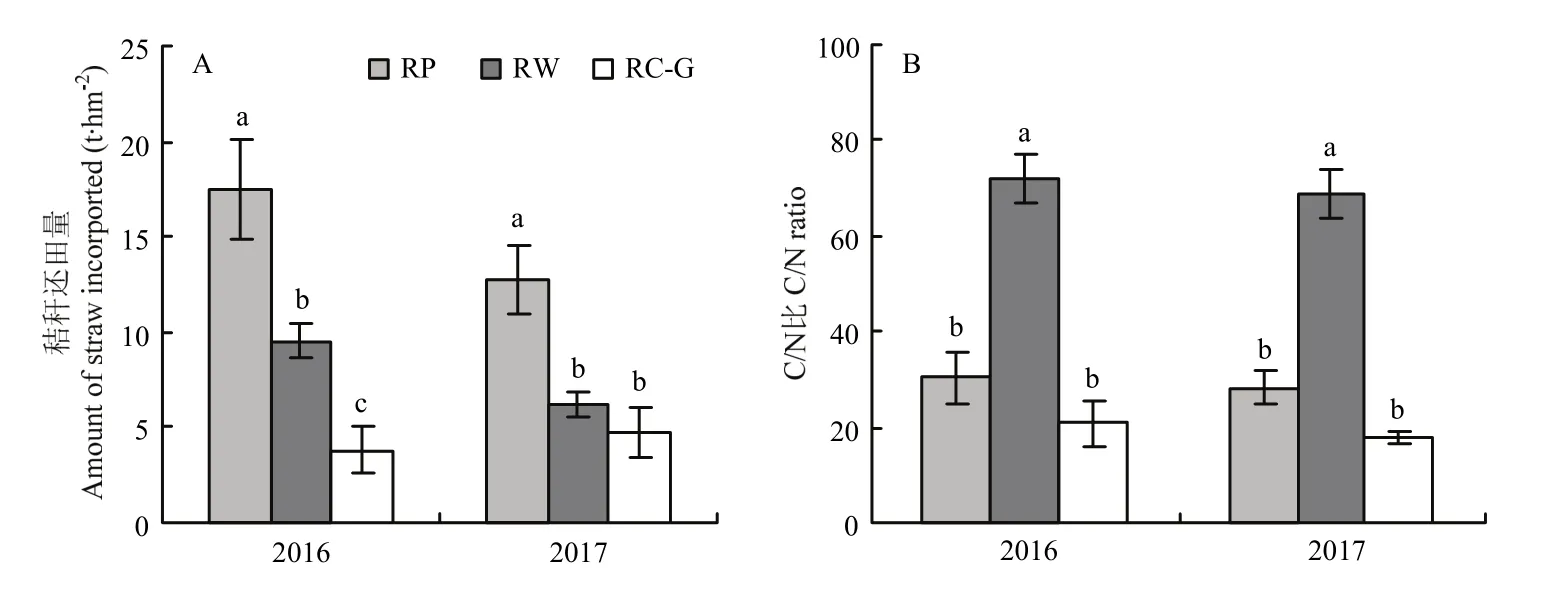
图1 不同轮作模式下秸秆还田量(A)和还田秸秆C/N比(B)Fig.1 Amount (A) and C/N ratio (B) of straw incorporated under different rotations
单季稻种植:供试品种为嘉58(常规粳稻)。水稻田于6月上旬翻耕。水稻浸种催芽后,5月中上旬播于秧田,至3叶1心左右移栽,行株距26.7 cm×13.3 cm。水稻季施氮肥,氮肥总量根据氮肥处理设置,其中基肥∶分蘖肥∶穗肥按照 40%∶30%∶30%施入;磷肥(过磷酸钙,60 kg·hm-2)作为底肥一次性施入;钾肥(氯化钾,135 kg hm-2)按基、蘖肥各50%分次施入。
秸秆还田处理:水稻人工收获,其秸秆全部移除用于冬季马铃薯覆盖栽培。冬季作物秸秆全量还田,不同轮作模式下秸秆还田量及还田秸秆C/N比如图1所示。不同轮作模式下秸秆还田量差异显著(图1-A,除2017年RW和RC-G差异不显著),与RP相比,RW和RC-G的秸秆还田量分别减少45.7%和78.3%(2016),51.6%和62.5%(2017)。长期还田秸秆的C/N比(图1-B)表现为RW显著大于RP和RC-G(RP和RC-G差异不显著);相较于RW,RP和RC-G轮作下C/N分别降低58.0%和71.3%(2016),58.9%和74.1%(2017)。因此,根据秸秆属性,可以将 4种轮作模式划分为RP(高秸秆量、低C/N比)、RW(高秸秆量、高C/N比)、RC-G(低秸秆量、低C/N比)和RF(对照-无秸秆)。
1.3 样品采集与分析
CH4气体采集于2016和2017年6—10月间开展,采用密闭式静态(透明)箱-气相色谱法测定田间CH4排放通量。气体采样箱由底座和采样箱组成,静态箱底横截面积为0.5 m×0.6 m,采样箱由PVC材质制成,箱体高度随水稻高度而增加(拔节前 0.5 m、拔节后1.2 m),中部固定风扇,箱内嵌有温度计,方便记录温度变化,底座为正方形U槽,嵌入地表,放置箱体后加水使之密封。
采集频率:水稻自移栽后第3天起,根据实际情况每5—10天采气1次。采样时间为8:00—11:00。气体采集前打开箱内顶部 12 V小风扇以充分混匀箱内气体,采样时,将采样箱垂直安放在底座5 cm深的凹槽内并加水密封,样品采集分别在关箱后的0、6、12、24 min进行。气体样中 CH4浓度由气象色谱(Agilent 7890A,美国安捷伦科技有限公司)分析测定。CH4检测器是 FID(氢火焰离子化检测器),检测温度300℃,柱温 60℃,载气为 99.999%高纯 N2,流速 30 mL·min-1。
土壤及秸秆属性测定:有机质含量采用水合热重铬酸钾氧化-比色法测定[67];全氮用全自动凯氏定氮仪(海能 K9840,济南海能仪器股份有限公司)测定;碱解氮采用碱解扩散法进行测定[67];土壤pH用pH计(F2,梅特勒-托利多仪器(上海)有限公司)进行测定。
鉴于田间试验处理,年度间基本一致,且经过多年定位试验,土壤性质基本稳定,因此于2018年分蘖期测定土壤氧化还原电位与土壤mcrA和pmoA基因丰度以探索CH4排放机理差异。
土壤氧化还原电位(Eh)测定(2018年):于水稻移栽后每5—7天测定一次。利用土壤氧化还原电位仪(FJA-6型,南京传滴仪器设备有限公司)测定根际土壤中的Eh值。
唐玉烟不好意思地笑了笑,道:“实在抱歉,那天,我蒙蒙眬眬地感觉四周不停地颠簸摇晃,就像地震了一般,后来又感觉一阵窒息,我在窒息中醒来,一睁眼,就发现自己正被一只怪物携着向前跑,我很害怕,于是拔出刀,割断了它的脖子。”
土壤mcrA和pmoA基因丰度测定:分别于水稻移栽后 3 d(2018/6/28)、14 d(2018/7/9)、28 d(2018/7/23)利用取土器采集每个小区距稻桩20 cm,耕层(0—20 cm)土壤样品(5点取样法取样),去杂质后混合待用。土壤mcrA和pmoA基因丰度采用实时荧光定量PCR分析。即用土壤DNA快速提取试剂盒提取土壤 DNA,用0.8%的琼脂糖凝胶电泳检测所提 DNA片段大小,并用超微量紫外分光光度计测定DNA浓度及质量。设计产甲烷菌的mcrA基因、甲烷氧化菌的pmoA基因引物,进行实时荧光定量PCR分析,反应体系为20 μL[25]。各基因定量PCR分析所用的引物和反应条件见表2,其中采集信号温度为72℃。

表2 光实时定量PCR扩增引物及反应条件Table2 Aplification primer and reaction condition of quantitative PCR
1.4 数据处理
温室气体排放通量由该气体在箱中浓度随时间的变化率计算得出,计算方程为气体排放通量计算公式如下:
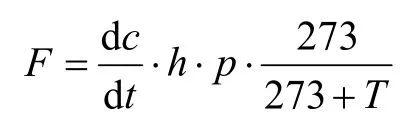
式中,F为排放通量(mg·m-2·h-1);dc/dt为采样过程中箱内气体浓度随时间的变化率(mL·m-3·h-1));h为采样箱体高度(m);ρ为标准状态下气体的密度(0.714 kg·m-3);T为采样时箱内温度(℃),273为气态方程常数。每次观测时各处理气体排放通量用3次重复的平均值表示;作物生长季节气体平均排放通量的计算则以相邻两次采样时间间隔为权重,将各次排放通量进行加权平均;气体累积排放量用平均排放通量和全生长季时间的乘积表示。
本文采用Excel 2010和SAS 9.2软件对数据结果进行计算和分析,采用 Duncan法分析数据间的差异显著性(P=0.05)。
2 结果
2.1 稻季CH4排放通量
水稻生长期稻田CH4排放通量的变化受水分管理影响显著,呈现明显的阶段性变化(图2),可观测到两个排放高峰期—分蘖期和孕穗期CH4排放阶段。其中分蘖期CH4排放阶段(约移栽后7—30 d)持续时间占全生育期的19.0%—19.7%。分蘖中后期(移栽后 37 d左右),稻田逐渐进入搁田期,各处理 CH4排放通量都表现不同程度下降。搁田复水后,CH4排放小幅上升,进入孕穗期 CH4排放阶段(约移栽后50—70 d),该阶段持续时间约占生育期的 16.7%—17.2%。之后进入低排放通量周期。
轮作模式对稻田CH4排放的影响因氮肥水平差异而有所不同。N0水平下,分蘖阶段CH4平均排放通量,RP、RW和RC-G无显著差异,但均显著高于RF,分别提高 72.1%—168.7%、70.8%—145.2%和 56.5%—145.2%(图2- A 和2-B)。孕穗阶段CH4排放通量,各模式间无显著差异,但2016年RP显著高于其他。此外,N0水平下分蘖期CH4排放表现为双峰特点(约为移栽后第12—15 d和25—30 d)。各轮作在移栽后25—30 d出现最大排放峰,RP、RW、RC-G的最高排放通量(两年平均)分别为55.0、42.9和38.6 mg·m-2·h-1。相比RF,RP、RW、RC-G的最高排放通量分别提高182.1%、120.0%和97.9%。
随着施氮量的增加,CH4排放通量对轮作的响应出现分化。RF、RP和RW的分蘖期和孕穗期CH4平均排放通量随着施氮量的增加而显著提高。N2水平下,RP 分蘖期 CH4平均排放通量为 36.2 mg·m-2·h-1,要比N1和N0分别高14.8%和40.4%;与RP相似,RW则要高13.9%和43.9%。RF则表现为分蘖期CH4排放通量,N1和N2显著高于N0,尤其在移栽后25—30 d,其平均排放通量达到23.8和27.5 mg·m-2·h-1(两年平均),相比N0,分别提高29.9%和50.3%。而RC-G分蘖期CH4排放通量保持相对稳定(2017)或略有下降(2016)。与 N0相比,增施氮肥下 RC-G分蘖期CH4平均排放通量降低20.0%—29.8%(N1)和11.9%—20.1%(N2)。
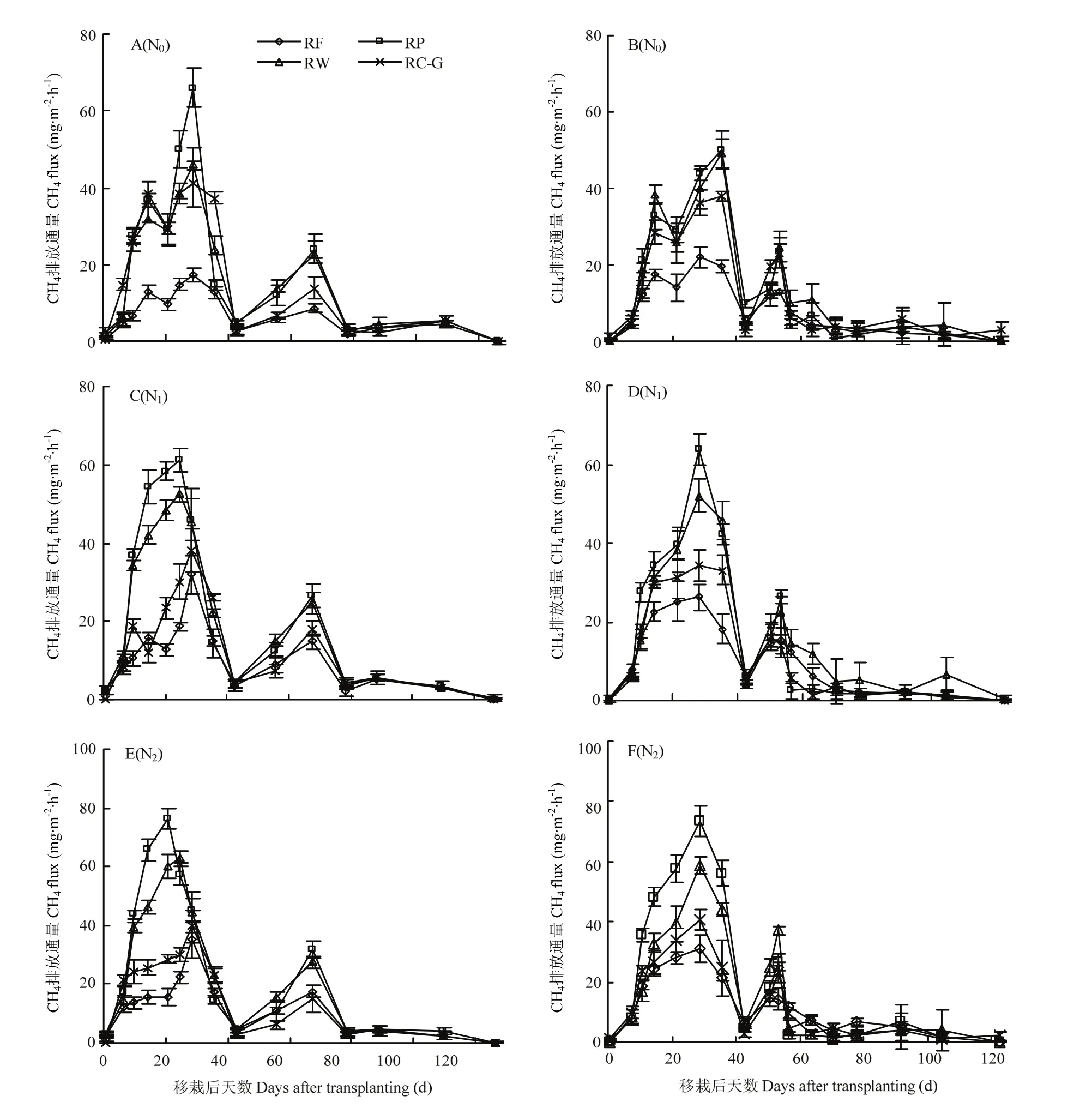
图2 2016(A、C和E)和2017(B、D和F)年度不同轮作模式及氮肥梯度下稻田CH4排放通量的季节变化Fig.2 Seasonal variations of CH4 fluxes in rice season under different rotation systems and N fertilizer rates in 2016 (A, C, E), 2017 (B, D, F)
2.2 稻季CH4累积排放量
由图3中可以看出,不同轮作模式及氮肥水平对稻季CH4排放量的影响存在显著互作。N0条件下,休闲(RF)模式的稻季CH4累积排放量要显著低于冬季作物种植(RP、RW 和 RC-G)。RF稻季年均 CH4累积排放仅 18.4—21.2 g·m-2,分别比 RP、RW 和 RC-G降低42.6%—59.4%、40.7%—52.1%和36.0%— 52.1%。
随着氮肥的增加,RP、RW和RF处理下稻季CH4累积排放量均有不同程度的增加。N2水平下,RP、RW 和 RF的 CH4累积排放量分别为 51.2—55.8、45.3—51.5 和 25.0—30.5 g·m-2,比 N0水平提高23.0%—38.4%、26.7%—33.7%和 35.3%—43.5%;而与 N1相比,则提高 9.9%—19.7%、20.8%—23.1%和17.4%—18.8%(2016,RF模式下,N1和N2无显著差异)。而RC-G稻季CH4累积排放量则随着施氮量的增加而显著降低。与N0相比,N1和N2下稻季CH4累积排放量分别降低20.7%—42.4%和10.6%—16.6%。不同轮作间比较,N1和N2水平下,RF和RC-G的稻季CH4累积排放量无显著差异,但显著低于RW和RP。
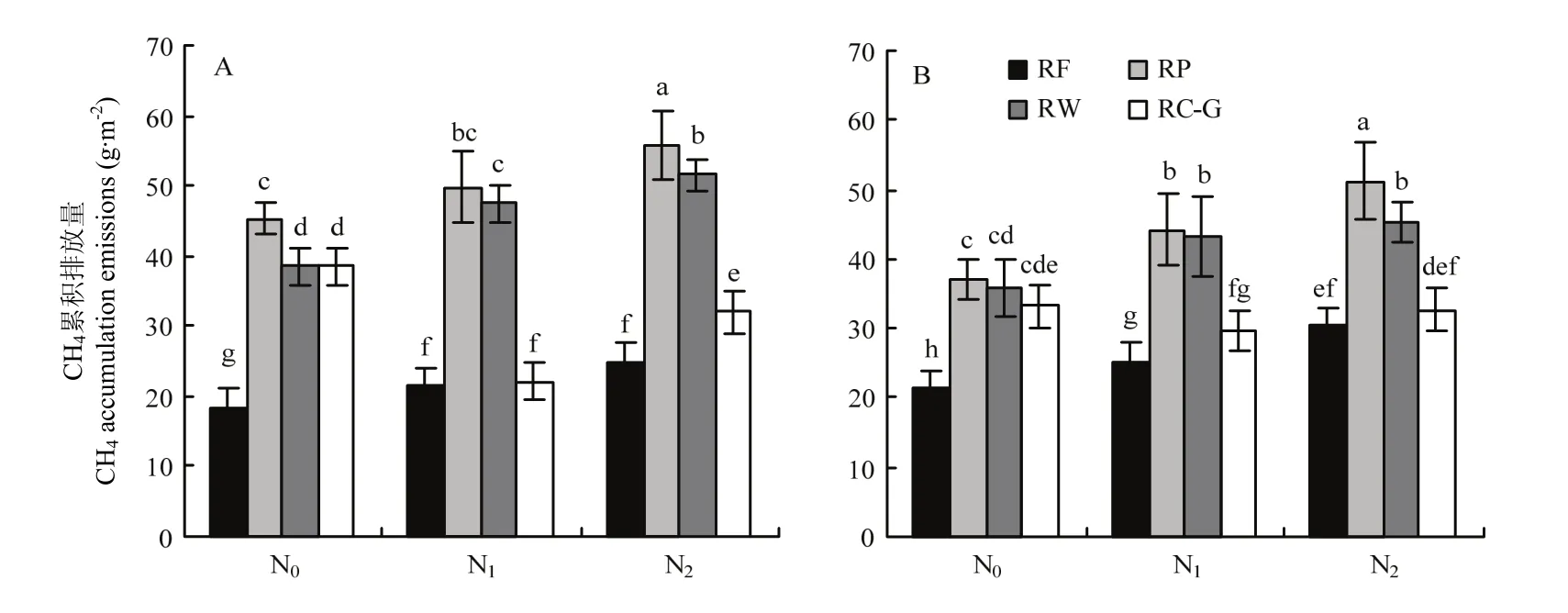
图3 2016(A)和2017年(B)不同轮作模式及肥料梯度下稻季CH4累积排放量Fig.3 CH4 accumulation emissions in rice season under different rotations and N fertilizer rates in 2016 (A) and 2017 (B)
稻季CH4排放通量还与水稻生育阶段相关。为了进一步明确不同轮作模式与氮肥水平下稻季CH4的排放规律,将水稻全生育期分为分蘖期、孕穗期和灌浆期3个主要生育阶段。总体而言,稻季CH4的累积排放量分蘖期>孕穗期>灌浆期,其中分蘖期、孕穗期和灌浆期占比分别为51.85%—72.31%、22.9%—31.3%和6.0%—20.7%,这表明CH4排放主要集中在生育前中期,而后期占比较小(图4)。轮作模式和氮肥水平对水稻不同生育阶段CH4累积排放量影响显著,差异主要集中体现在分蘖期和孕穗期,而灌浆期则无显著差异。N0水平下,与RP、RW和RC-G相比,RF的分蘖期 CH4累积排放量降低 95.9%—171.1%、70.7%—144.1%和74.2%—165.8%;随着氮肥的施用,各轮作分蘖期CH4累积排放量占比均显著提高(2016年RC-G除外),增幅为7.0%—48.3%。孕穗期CH4累积排放量,RP和RW模式显著高于RC-G和RF,而RC-G和RF无显著差异。灌浆期,CH4排放量较小,各处理之间无显著差异。
2.3 产甲烷菌(mcrA)和甲烷氧化菌(pmoA)基因丰度
土壤中产甲烷菌(mcrA)和甲烷氧化菌(pmoA)是影响稻季CH4排放的重要微生物。为进一步明确不同轮作模式下稻季CH4排放微生物差异,于2018年水稻移栽后3、14和28 d采集并测定土壤中mcrA和pmoA基因丰度(图5)。不同轮作模式与氮肥水平下,mcrA和pmoA基因丰度差异显著。N0水平下,移栽后3 d,mcrA基因丰度量较少,而移栽14 d和28 d后丰度显著增加。移栽后28 d mcrA基因丰度大小顺序为 RP>RW>RC-G>RF,相较于RF,RP、RW 和 RC-G模式下mcrA基因丰度增加195.7%、121.3%和45.7%,这说明秸秆还田有利于提高土壤中产甲烷菌丰度。施氮后,各轮作模式mcrA基因丰度均显著增加,相较于N0,N1水平下RP、RW、RC-G和RF的mcrA基因丰度分别增加191.4%、135.6%、43.1%和105.7%(移栽后28 d),其中以RC-G增加最少。
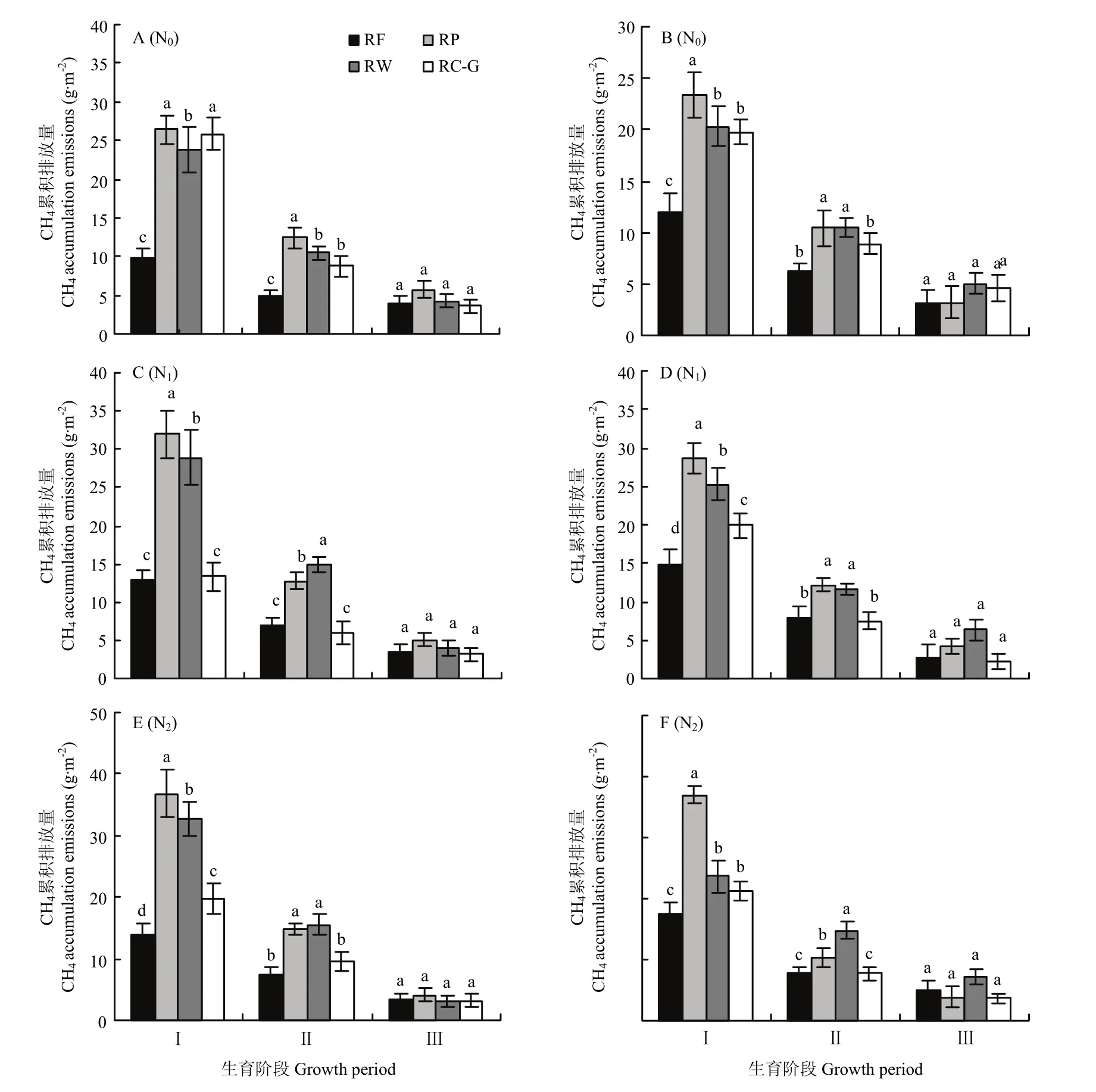
图4 2016(A、C和E)和2017(B、D和F)年度不同轮作模式及氮肥梯度下水稻不同生育阶段CH4累积排放量Fig.4 CH4 accumulation emissions at different rice growth periods under different rotation systems and N fertilizer rates in 2016(A, C and E) and 2017 (B, D and F)
与mcrA不同,N0水平下,移栽后14和28 d,RP和RW的pmoA基因丰度显著高于RF和RC-G。增氮后,RF、RW和RP的pmoA基因丰度显著减少降幅分别为42.7%、14.1%和13.3%(移栽后28 d),说明氮肥一定程度上抑制了甲烷氧化菌的活性,而RC-G则刚好相反,增施氮肥后pmoA基因丰度增加72.8%(移栽后28 d)。
以上结果表明,随着氮肥的增施,RP、RW和RF的产甲烷菌显著提高,但RC-G增加较少;而甲烷氧化菌的丰度变化则与CH4排放刚好相反,表现为RP、RW和RF pmoA基因丰度减少,而RC-G增加。进一步分析mcrA/pmoA比值(图6-E和6-F),发现N0水平下,不同轮作模式下 mcrA/pmoA比值大小顺序为RP>RW、RC-G>RF(移栽后14 d和28 d),其变化规律与稻季不同轮作方式下CH4排放变化基本一致。增施氮肥后RF、RP和RW mcrA/pmoA基因丰度比值显著提高,其增加比例分别为 71.4%—141.1%、197.1%—258.2%和84.6%—165.5%;而RC-G则下降26.8%—42.3%。这与田间CH4的排放规律相似。
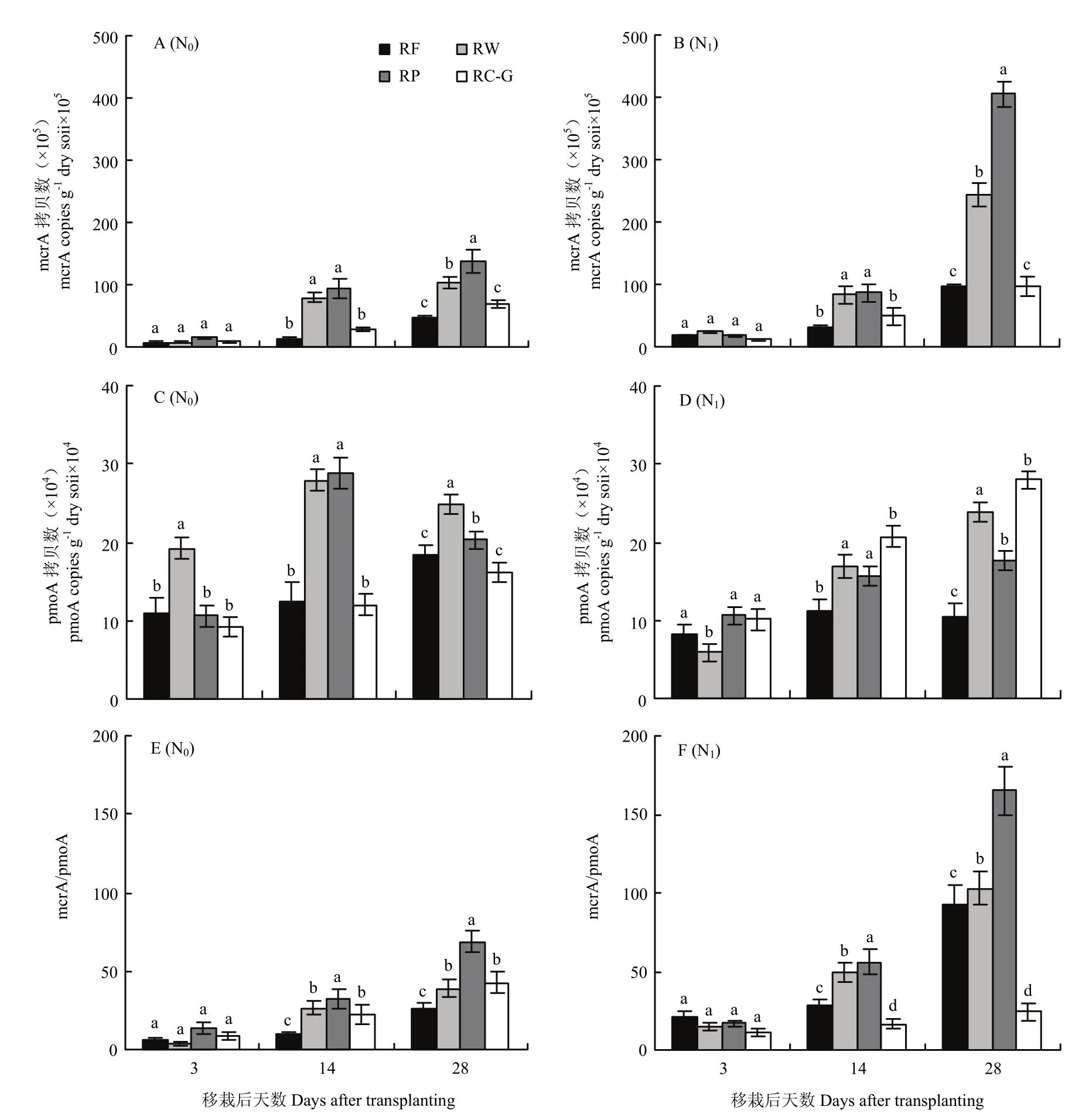
图5 2018年不同轮作模式及氮肥梯度下产甲烷菌基因(mcrA;A和B)和甲烷氧化菌基因(pmoA;C和D)拷贝数和mcrA/pmoA比值(E和F)Fig.5 Abundance of methanogenic gene mcrA (A and B) and methanotrophic gene pmoA (C and D) and mcrA/pmoA ratio (E and F) under different rotation systems and N rates in 2018
2.4 氧化还原电位差异性
不同轮作模式下氧化还原电位(Eh)变化规律在不同氮肥处理下基本一致(图6),搁田前(稻田一直处于淹水状态),各轮作模式 Eh持续下降,在移栽后18—25 d降到最低,然后开始回升,移栽30—35 d以后Eh>0,并呈波浪式变化。N0水平下,水稻移栽前期(3—7 d),此时各轮作模式 Eh大小顺序为RP>RW>RC-G>RF(轮作间 Eh差异不显著),可能是由于秸秆的添加与机械翻耕增加了土壤的通气性。移栽后22 d,RP、RW和RC-G模式之间无显著差异,但均显著低于 RF模式,降幅为 118.4%、102.6%和80.5%,说明随着淹水时间的延长,秸秆的降解消耗了土壤中的O2,加速了土壤中氧化还原电位的降低。增施氮肥后,Eh变化幅度因轮作模式而有所差异(移栽后22 d),N2水平下,RP、RW、RC-G和RF的Eh分别为-198.9、-188.5、-156.3和-80.3 mV(RP、RW 和 RC-G模式之间无显著差异,但均显著低于RF模式),与N0相比,分别降低33.1%、34.5%、29.7%和24.2%。而与N1相比,则降低7.2%、20.2%、38.4%和 15.9%。无机氮肥的投入,一定程度上促进秸秆的腐解速率,并进一步加速土壤中氧化还原电位的降低。
3 讨论
3.1 轮作模式对稻田CH4排放的影响
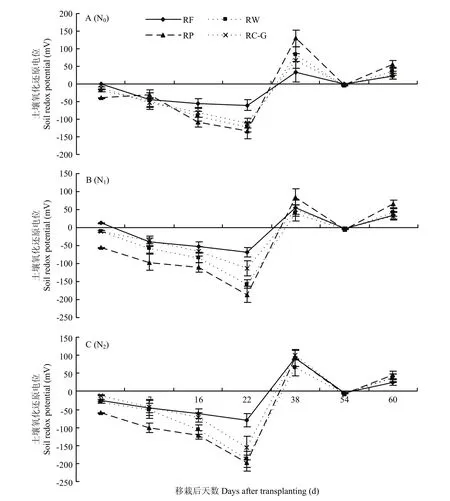
图6 不同轮作模式及氮肥梯度下稻季土壤氧化还原电位变化规律Fig.6 Variations of soil redox potential in rice season under different rotation systems and N fertilizer rates
轮作处理与氮肥水平并未改变稻季CH4总的排放规律。受田间水分管理和生长发育影响,稻季CH4排放主要集中在水稻分蘖期,其它时期(如拔节抽穗期、灌浆结实期)相对较少[28-29]。在水稻生长季,CH4排放表现为分蘖期前期升高,中后期特别是搁田开始后降低,搁田结束稻田复水后略有升高,并保持低排放量直至水稻收获,呈现波浪式变化[5]。
本研究发现轮作模式与氮肥水平对稻季CH4不同阶段排放通量及其累积排放量均有显著影响,其影响主要集中在分蘖期,这与张岳芳[30]和黄太庆[31]等的部分研究结论一致。另外,张啸林等[32]研究认为添加秸秆和绿肥作物均增加稻季CH4排放,而添加高生物量秸秆比添加低生物量绿肥作物有更多的CH4排放。这与本研究施氮(N1和 N2)处理下不同轮作模式 CH4排放规律基本一致;但在N0下,RP、RW和RC-G无显著差异。其变化规律还与产甲烷菌基因丰度密切联系,即添加秸秆生物量与CH4排放之间的关系受氮肥施用调控,并受到产甲烷菌活性的制约。微生物增殖与激活需要底物和能量,而土壤有效碳量是影响土壤微生物活性的限制因子[33]。稻田CH4排放主要受限于产甲烷菌和甲烷氧化菌的丰度及其活性。产甲烷菌是稻田土壤中甲烷产生的主要生物源[34]。而甲烷氧化菌,是以甲烷作为唯一碳源和能源进行同化和异化代谢的微生物[35]。李大明等[36]研究指出产甲烷菌丰度与稻田CH4排放量呈正相关,这与本研究结果一致。秸秆还田与氮肥处理影响了有机碳的转化和产甲烷菌与甲烷氧化菌可利用的有机碳及能量,进而影响到CH4排放[37]。在种植系统中,秸秆全量还田能够显著增加产甲烷菌丰度,进而导致稻田CH4排放量提高[38]。本研究发现N0下,产甲烷菌丰度变化规律与秸秆还田量有关,这与BAO[38]和ARIF[39]等关于大量秸秆碳的投入增加稻田产甲烷菌数量的结果一致。另一方面,稻田CH4排放量也因秸秆还田的有机物料种类而异。前人研究认为添加的有机物料C/N越高,能源物质越丰富,土壤有关微生物活性就越高,在环境适宜的条件下,CH4排放就越多[40]。本研究中RW、RP和RC-G还田秸秆的C/N比分别为70.39、29.25和19.26(图1 B),其中RP处理C/N较低,但其CH4排放通量显著高于RC-G,这与其秸秆还田量显著高于RC-G相符(图1-A)。显然,以还田秸秆 C/N比作为判断还田秸秆对 CH4排放的影响亦不全面。笔者认为与C/N相比,还田秸秆中有机碳含量可能与 CH4排放关系更加密切,但其效应受氮肥施用的影响。此外,长期定位轮作后,RP土壤中较高的有机质含量可能也是导致 RP轮作模式 CH4排放量最大的重要因素(表1)。
3.2 轮作模式和氮肥水平对稻田CH4排放的互作效应
氮肥能够影响稻田CH4排放,其效应受种类、施用量、施用方式和施用时间等影响[23]。长期还田秸秆的C/N比能干扰增施氮肥带来的CH4排放效应。在本研究的RP、RW和RF轮作模式中,我们发现,增施氮肥能够显著增加稻田CH4排放(图3),与前人研究印证[41]。但RC-G的表现则刚好相反。这与ZHU[42]和胡安永等[5]关于紫云英-水稻轮作模式不同施氮量下CH4排放研究结论一致。氮肥与有机物料的互作效应对稻季CH4排放的影响主要与有机物料分解及其后续有机物种类有关[43]。一般而言,具有较高C/N比的有机底物,其分解速率随着氮输入量的增加而增加[44]。另外秸秆的分解速率还与相关微生物丰度密切相关,后者则受到土壤中有效氮含量的制约[45],研究发现,在氮受限的环境中,增加氮的输入提高土壤中氮的有效性,这相应提高了土壤中产甲烷菌所需有机底物的有效性,促进了产甲烷微生物活性,增加CH4产生[46]。本研究进一步发现微生物对氮肥的响应机制因轮作模式而有所差异,增施氮肥促进产甲烷菌的增殖,却抑制了甲烷氧化菌的增殖(图5-A,5-B),但其变化幅度因轮作处理而有所不同。这与许欣等[25]关于土壤中可利用氮含量(铵态氮与硝态氮)与mcrA基因丰度正相关而与 pmoA基因丰度呈显著负相关的结论一致。笔者认为CH4排放对氮肥的响应,原因可能是:(1)产甲烷菌主要以NH4+-N为氮源,外源无机氮的输入提高了产甲烷菌繁殖所需底物的有效性[46],会加快产甲烷菌的生理活动[47],从而促进其群体的快速增长[48];(2)甲烷单氧酶(MMO)是甲烷氧化菌的关键功能酶,可以在氧气的作用下催化甲烷等低碳烷烃或烯烃羟基化或环氧化[49],NH4+是MMO的竞争性抑制剂,通过与MMO的竞争,减少了甲烷氧化,促进了甲烷排放[50];(3)氮素增加还会引起植物根系分泌量增加,进而为产甲烷菌带来更多生活底物[51];(4)由于产甲烷菌代谢过程严格厌氧,只能生活在氧化还原电位很低的环境中[52]。增加无机氮的投入,能够加速有机物料的腐解速率,腐解的同时消耗了水中的O2,降低了土壤中的氧化还原电位(图6-B和6-C),进一步增加了产甲烷菌的数量,进而促进CH4的排放。另外,WATANABE等[53]研究发现,稻田CH4排放与秸秆还田量显著正相关,当秸秆还田量为 2、4和 6 t·hm-2时,CH4排放量分别增加 19%、97%和 228%,其中秸秆中的碳对CH4排放的贡献分别为19%、51%和60%。本研究中,RW和RP中秸秆还田量达到15.15和 7.85 t·hm-2,属于高还田量栽培。微生物利用碳源进行繁殖增长的同时,需要外界提供较高的氮源[46]。而RC-G处理本身带入秸秆量较少(4.30 t·hm-2),含氮量较高。鉴此,笔者认为稻田系统中秸秆还田C/N的相对含量可能是干扰氮肥水平对稻田CH4排放作用的关键。另外,本研究发现,稻田土壤CH4排放量并不是取决于产甲烷菌或甲烷氧化菌中的某一个,而产甲烷菌与甲烷氧化菌基因丰度比值(mcrA/pmoA)能更客观的反映CH4产生、氧化及排放之间的数量关系,这与LIU[52]等研究结果一致。当土壤中生成的CH4通过扩散进入氧化区域时,大量CH4被甲烷氧化菌氧化。因此稻田土壤的CH4排放量不仅取决于CH4的生成能力,还决定于土壤对内源CH4的氧化能力[54]。本研究表明,相较N0水平下,增施氮肥(N1)显著增加RF、RW和RP轮作模式mcrA/pmoA比值;而RC-G轮作模式却正好相反。其变化规律与CH4排放量基本一致。表明紫云英绿肥还田配施氮肥(N1),对土壤中甲烷氧化菌的促进作用强于产甲烷菌,有助于减少稻田CH4排放。
3.3 水稻植株及冬季作物种植对稻季CH4排放的影响
稻田甲烷排放是稻田甲烷产生、氧化和传输综合作用的结果,而水稻的生长强烈影响这3个过程[55]。水稻根系作为重要的中间载体,其根系形态和生理特征严重影响CH4排放[56],而投入的有机物料及其分解后有机物种类对根系的影响程度不同,最终会导致甲烷排放的差异。王红妮等[57]发现水稻根系为了维持正常的呼吸作用,大量消耗根系附近的氧气,降低了根系土壤的氧化还原电位,促进了稻田土壤厌氧环境的形成,这也可能是导致不同轮作模式下稻田 Eh差异性的原因之一(图6);同时,水稻根系通过分泌物和凋落物的形式为产甲烷菌提供底物,促进产甲烷的增殖[58]。另外,水稻植株也会影响稻田CH4氧化作用,AULAKH等[59]研究指出大气中的氧气可以通过水稻植株的通气组织进入根系,形成一个好氧微区域,促进甲烷氧化菌的生长,氧化稻田土壤中产生的甲烷;稻田土壤中产生的甲烷大约有36%[55]到80%甚至更多[60]在根区微氧区域被甲烷氧化细菌消耗。本研究中紫云英绿肥还田配施氮肥后CH4排放相对较低,是否与水稻植株根系发育程度及其通气组织有关还有待更深入的研究。笔者认为大量有机物料及无机氮肥的投入对水稻植株及其根系影响程度存在差异,进而造成稻田甲烷产生、氧化和传输过程的不同,最终导致CH4排放存在差异。
本研究发现系统中C/N相对含量与CH4排放密切联系。冬季轮作及冬作后残留氮会影响土壤中有效氮含量[61-62],同时也会影响稻季秸秆还田和氮肥施用,这也可能是导致温室气体差异的原因之一。土壤中有效氮源(碳源)是微生物繁殖的限制因素,而不同冬季轮作种植能够影响土壤有机质含量、培肥地力及改善土壤结构[20],进而改变了微生物代谢的有效有机碳/氮的组成与数量[63],从而显著影响相关微生物的生存能力和活性[33]。本研究中,相较于 RF,RP、RW 和RC-G轮作系统下土壤全氮分别提高24.3%、14.0%和7.9%,有机碳分别提高24.2%、7.2%和6.3%(表1)。这与粱爱珍[64]和汪娟[65]等的部分结论一致。土壤全氮是能够反映土壤供氮能力的重要指标,而有机碳在一定程度上影响着土壤结构及其微生物多样性,同时又是土壤肥力和基础地力的最重要的物质基础[66]。尽管不同处理下,土壤属性有所差异,但轮作研究中土壤属性的变化缓慢而微量。而与秸秆还田和氮肥施用带来迅速且高通量的CH4排放相比,土壤地力的变化是否是影响当季CH4排放的重要因子,有待进一步研究。
4 结论
N0处理不同轮作模式下稻田CH4排放差异显著,具体表现为RP>RW>RC-G>RF;增施氮肥后,轮作模式与氮肥水平存在显著互作,RW、RP和RF的CH4排放显著增加,而RC-G则略有下降;产甲烷菌与甲烷氧化菌基因丰度比值(mcrA/pmoA)与CH4排放规律基本一致。鉴此,笔者认为稻田系统中秸秆还田C/N的相对含量可能是干扰氮肥水平对稻田CH4排放作用的关键,当系统中碳冗余时,相关微生物活性受到土壤中有效氮制约,投入无机氮可以减轻氮的限制作用从而显著提高CH4排放;而碳不足时,继续投入无机氮,相关微生物繁殖由于受到土壤中有限碳源的限制其活性也会受到抑制,CH4排放相对减少。
猜你喜欢
河南水利与南水北调(2022年7期)2022-08-18
海洋通报(2022年2期)2022-06-30
农业灾害研究(2022年1期)2022-05-07
今日农业(2021年11期)2021-11-27
吉林蔬菜(2021年2期)2021-07-19
暴雨灾害(2021年2期)2021-04-02
军民两用技术与产品(2021年10期)2021-03-16
水上消防(2020年1期)2020-07-24
今日农业(2019年12期)2019-08-15
疯狂英语·新读写(2018年3期)2018-11-29
