
吹绵蚧卵孵化期的生物学观察研究
2019-07-30 01:05吴维民王静盛辉葛银银李照坤
安徽林业科技 2019年3期
吴维民 ,王静 ,盛辉 ,葛银银 ,李照坤
(1.合肥野生动物园,合肥 230031;2.合肥植物园,合肥 230031;3.安徽农业大学林学与园林学院,合肥 230036)
吹绵蚧(Icerya purchasi)属同翅目、蚧壳虫总科、吹绵蚧属,在热带及温带地域广泛分布。该害虫是多食性昆虫,在我国危害80余科250多种植物,是柑橘等果树及樱花等园林植物上的重要害虫[1]。吹绵蚧刺吸危害植株枝叶,造成叶片发黄凋落,致使树势衰弱,危害严重时甚至导致整株植物死亡。同时,吹绵蚧分泌的蜜露还会诱发煤污病,既影响植株的光合作用,又降低园林植物的观赏价值[2]。吹绵蚧在长江流域1年发生2~3代,由于吹绵蚧繁殖力强,每只雌成虫可产卵为100~1 400粒不等[3],极易暴发,是园林植物与果园的重点防治对象之一。
与多数蚧壳虫害一样,化学防治吹绵蚧常常难以取得满意效果,因为吹绵蚧孵化后体背会逐渐生出蜡质物,形成的蜡质层可以有效保护虫体,降低化学农药的渗透,使得此类害虫的化学防治往往难以取得令人满意的效果,这也是植物保护一线工作者常常面临的难题。有不少研究考虑到大量化学农药的不科学使用会对环境、生态平衡造成负面影响[4-5],从而寻求生物防治与物理防治等方法控制蚧壳虫[6],但生物防治效果往往受环境因素影响很大且作用效果较为缓慢,而物理防治更是费时费工。研究表明,在蚧壳虫初孵的分布扩散期,初孵若虫还未形成蚧壳,此时若虫对药剂的触杀作用最为敏感,是蚧壳虫防治的最佳时期,只要药剂喷洒周到,杀虫效果可达95%以上[7]。因此,充分了解吹绵蚧卵孵化期的生物学特性是确定该害虫防治适期的关键。
1 材料与方法
1.1 供试昆虫
供试昆虫为产卵期的吹绵蚧雌成虫,采自合肥大蜀山森林公园及安徽农业大学植物园的日本晚樱 (Cerasus serrulata) 及绣球荚蒾(Viburnum macrocephalum)植株上。
1.2 试虫采集与卵的孵化观察
4月初起,在大蜀山森林公园踏察越冬代吹绵蚧的产卵情况;4月下旬,用剪刀剪取附着有吹绵蚧成虫及卵囊的日本晚樱及绣球荚蒾枝叶,枝长约30 cm;将新鲜的枝叶置于装有纯净水的250 mL三角瓶中,每个三角瓶放置1根植物枝条,每根枝条上的每片叶片最多保留1个带有卵囊的雌成虫。试验设3个重复。将处理好的枝条置于室内朝南有阳光照射的开放环境中,每日体视镜下观察吹绵蚧卵囊的孵化情况。在第一只初孵若虫爬出卵囊后,每天上午和下午各在体视镜下观测、统计孵出的若虫数量,并用湿棉签将统计过的若虫进行清理,以便后续的孵化观察。
1.3 初孵若虫的显微观察
部分枝叶上的吹绵蚧初孵若虫爬出卵囊后,不做若虫清除,每日体视镜下观察若虫的活动情况与形态变化,并在体视镜下拍照,观测吹绵蚧若虫体壁的蜡质形成过程。
1.4 数据处理
数据利用DPS软件进行平均数、标准差及差异性分析。
2 结果与分析
2.1 卵的孵化与若虫分布
2.1.1 初孵若虫形态学观察
吹绵蚧初孵若虫呈椭圆形,体橙红色,离开卵囊1~2 d内爬行十分活跃,快速分散于叶脉两侧。在体视镜下观察发现,刚孵化出的吹绵蚧若虫头胸腹区分明显,能够清晰看见若虫的眼、足及触角各器官,活跃且移动迅速,是吹绵蚧一生中最关键的分布时期。在初孵若虫离开卵囊2~3 d内,大多数会在叶脉,尤其是主脉附近固定危害,不再轻易移动;此时若虫黏着于叶片表面,体色偏向橙黄色,身形扁平,足与触角已不好辨识,眼睛尚模糊可见。在初孵若虫固定危害1~2 d内,即开始分泌蜡质物;此时若虫体黄色,身体变得十分扁平,其体背隐约可见透明沟痕。若虫固定3~4 d后清晰可见黏附在叶表的若虫体背具透明“十字”型沟纹,虫体变得晶莹透亮,此时分布于叶脉两侧的吹绵蚧体壁开始全面覆盖薄层的蜡质,体视镜下虫体折光率与质感的变化可以清晰辨识。见图1。

图1吹绵蚧初孵若虫形态学观察
2.1.2 不同植物叶片对吹绵蚧若虫分布的影响
对寄生于日本晚樱与绣球荚蒾上吹绵蚧的分布进行体视显微观察后发现,吹绵蚧若虫在两种植物上固定危害的分布存在差异。吹绵蚧若虫在日本晚樱上固定危害时,若虫密集于叶脉两侧;在绣球荚蒾上固定危害时,若虫的分布则并无明显特征,它们较为均匀的分布在叶片表面。若虫活跃的分散期,在日本晚樱上多集中分散于叶背,在绣球荚蒾上它们则更多向与密披绒毛的叶背相反的叶面移动。吹绵蚧在这两种植株上的不同分布表现,应该是取决于这两种植物叶片上形态特征差异。绣球荚蒾叶背密被的绒毛对具刺吸式口器的吹绵蚧吮吸汁液的习性具有一定阻碍作用,由此它们趋向于移动至更易取食的叶面。所以,为了更有效进行吹绵蚧防治,应根据它所依附的植物的形态特征以及主要危害部位制定对应的防治措施。
2.2 卵的孵化进程观察
2.2.1不同寄主植物上吹绵蚧卵量的变化
通过对日本晚樱和绣球荚蒾寄主植物上吹绵蚧产卵量与孵化率的观察比较,吹绵蚧在这两种不同植物寄主上的卵孵化数量有着明显差异(图2)。经统计,危害日本晚樱的吹绵蚧,每只雌虫卵囊的若虫孵化数量在239~422头,平均数量为334头;危害绣球荚蒾的吹绵蚧,每只雌虫卵囊的若虫孵化数量在132~250头,平均数量仅190头;两者之间的产卵量有着明显的差异。导致同一地区不同植物上吹绵蚧产卵及孵化数量具明显差异的原因不仅与不同寄主植物对吹绵蚧的营养供给有关,也与不同寄主植物的抗虫性差异有关。
2.2.2 吹绵蚧卵的孵化进程
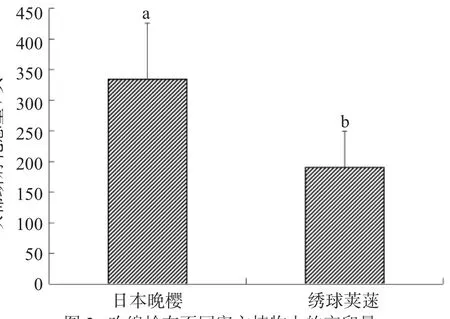
图2吹绵蚧在不同寄主植物上的产卵量
每日观察吹绵蚧在两种不同寄主植物上的孵化进程结果见图3。吹绵蚧在日本晚樱上的孵化期为7~10 d,吹绵蚧若虫的孵化比较集中,在孵化第2天到达孵化高峰期,孵化5~6 d后若虫孵化量明显降低,第9天若虫基本孵化完毕。吹绵蚧在绣球荚蒾上的孵化期则相对更为集中,第3天左右达到单日孵化峰值。吹绵蚧在绣球荚蒾上的卵孵化曲线呈明显的单峰形态,孵化虫口数量在逐步上升达到峰值后,又逐渐下降,第7天时若虫就基本孵化完毕。
吹绵蚧孵化高峰期在两种植株上也有所不同,在日本晚樱上达到孵化峰值的日期为5月9日,孵化虫口数为153头,占孵化虫口数总值的36.26%;在绣球荚蒾上达到孵化峰值的日期为5月12日,孵化虫口数为95头,占孵化虫口数总值的38.00%。尽管吹绵蚧在两种植物上的孵化峰值期不同,但到达孵化高峰期的时间基本是在若虫开始孵化1~2 d后发生,发生期限2~3 d。因此,在5月份发现植株枝叶上附着吹绵蚧若虫时,应尽快用化学农药喷洒植株,此时期孵化不久的吹绵蚧尚未分泌蜡质物,化学农药可以轻易杀死它们。
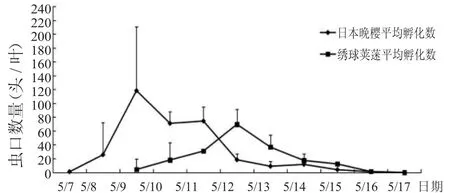
图3吹绵蚧在不同寄主植物上孵化数量动态
3 讨论与结论
5月初,合肥地区吹绵蚧卵从初孵期到孵化终期,整个孵化期为7~10 d;开始孵化后的2~4 d为孵化高峰期。对吹绵蚧初孵若虫的显微观察显示,若虫离开卵囊3~4 d内开始明显分泌蜡质物,一周就可在虫体表面形成薄层蜡质。同时,吹绵蚧在不同植物上的产卵量、空间分布及孵化特性均存在一定差异。结合吹绵蚧卵的孵化特点与若虫生物学特性,吹绵蚧的最佳防治时期为其若虫爬出卵囊的2~4 d,此时吹绵蚧为孵化高峰,同时体壁蜡质覆盖较少。因此,最迟防治时期不要超过若虫出现后的7 d。
蚧壳虫的最佳防治期就是蚧壳虫个体对农药最为敏感、抗药性弱、作用面大、防治效果最好的时期。吹绵蚧的最佳防治期就是在卵的孵化高峰且初孵若虫还未在整个体壁形成蜡层的时期,此时不仅其体壁薄而且没有或稀有蜡质覆盖,农药可以有效通过体壁渗透进入血腔,从而杀死害虫。本研究对吹绵蚧的显微观察确定了吹绵蚧卵的孵化比较集中,且在孵化一周内体壁上可形成明显蜡质层覆盖,这说明对吹绵蚧等蚧壳虫的防治最佳时期很短。从研究结果来看,吹绵蚧若虫孵化量高峰期为若虫爬出卵囊后2~4 d内,在此高峰期内喷洒化学药剂才能起到高效灭虫作用。
蚧壳虫与植物之间存在着复杂的生物学关系,有着协同进化的现象,近年来发现同种蚧壳虫因寄生不同植物而产生多型或双型现象[8]。对于吹绵蚧来说,它是一种多食性的害虫,危害樱花、柿树、绣球荚蒾、石榴等数十种常见植物,尤其对柑橘危害最盛。本研究证实吹绵蚧虽然能在日本樱花和绣球荚蒾上良好地生存,但不同寄主植物对其产卵量的影响还是显著的。因此对于吹绵蚧危害严重的区域,可以通过植物种群结构的调整来达到对吹绵蚧种群长期控制的目的。
猜你喜欢
辽宁农业科学(2021年6期)2021-12-02
少年漫画(艺术创想)(2020年2期)2020-06-15
少年漫画(艺术创想)(2020年1期)2020-05-20
歌海(2019年6期)2019-02-22
江苏农业科学(2017年7期)2017-05-23
中国蔬菜(2015年11期)2015-12-21
山东农业科学(2014年9期)2015-01-07
中国粮油学报(2014年8期)2014-02-06
中国果业信息(2014年9期)2014-01-23
作文周刊·小学六年级版(2009年50期)2009-08-27
