
Atsttrin reduces lipopolysaccharide-induced neuroinflammation by inhibiting the nuclear factor kappa B signaling pathway
2019-07-18 09:53:40LianLiuYuanQuYiLiuHuaZhaoHeChengMaAhmedFayyazNoorChangJiaoJiLinNieMengSiLeiCheng
中国神经再生研究(英文版) 2019年11期
Lian Liu , Yuan Qu , Yi Liu Hua Zhao He-Cheng Ma Ahmed Fayyaz Noor, Chang-Jiao Ji, Lin Nie Meng Si , Lei Cheng
1 Department of Orthopedics, Qilu Hospital of Shandong University, Jinan, Shandong Province, China
2 Department of Orthopedics, Qilu Children's Hospital of Shandong University, Jinan, Shandong Province, China
3 Department of Hand Surgery, the Second Hospital of Jilin University, Changchun, Jilin Province, China
4 Department of Chemistry, University of Massachusetts Lowell, Lowell, MA, USA
5 Department of Orthopedics, the Affiliated Hospital of Shandong University of Traditional Chinese Medicine, Jinan, Shandong Province, China
Abstract Progranulin is closely related to neuronal survival in a neuroinflammatory mouse model and attenuates inflammatory reactions. Atsttrin is an engineered protein composed of three progranulin fragments and has been shown to have an effect similar to that of progranulin.Atsttrin has anti-inflammatory actions in multiple arthritis mouse models, and it protects against further arthritis development. However,whether Atsttrin has a role in neuroinflammation remains to be elucidated. In this study, we produced a neuroinflammatory mouse model by intracerebroventricular injection of 1 µL lipopolysaccharide (10 µg/µL). Atsttrin (2.5 mg/kg) was administered via intraperitoneal injection every 3 days over a period of 7 days before intracerebroventricular injection of 1 µL lipopolysaccharide (10 µg/µL). In addition,astrocyte cultures were treated with 0, 100 or 300 ng/mL lipopolysaccharide, with 200 ng/mL Atsttrin simultaneously. Immunohistochemistry, enzyme-linked immunosorbent assay and real-time reverse transcription-polymerase chain reaction were performed to examine the protein and mRNA levels of inflammatory mediators and to assess activation of the nuclear factor kappa B signaling pathway. Progranulin expression in the brain of wild-type mice and in astrocyte cultures was increased after lipopolysaccharide administration. The protein and mRNA expression levels of tumor necrosis factor-α, interleukin-1β and inducible nitric oxide synthase were increased in the brain of progranulin knockout mice after lipopolysaccharide administration. Atsttrin treatment reduced the lipopolysaccharide-induced increase in the protein and mRNA levels of tumor necrosis factor-α, interleukin-1β, matrix metalloproteinase-3 and inducible nitric oxide synthase in the brain of progranulin knockout mice. Atsttrin also reduced the expression of cyclooxygenase-2, inducible nitric oxide synthase and matrix metalloproteinase 3 mRNA in lipopolysaccharide-treated astrocytes in vitro, and decreased the concentration of tumor necrosis factor α and interleukin-1β in the supernatant. Furthermore, Atsttrin significantly reduced the levels of phospho-nuclear factor kappa B inhibitor α in the brain of lipopolysaccharide-treated progranulin knockout mice and astrocytes, and it decreased the expression of nuclear factor kappa B2 in astrocytes. Collectively, our findings show that the anti-neuroinflammatory effect of Atsttrin involves inhibiton of the nuclear factor kappa B signaling pathway, and they suggest that Atsttrin may have clinical potential in neuroinflammatory therapy. The study was approved by the Animal Ethics Committee of Qilu Hospital of Shandong University, China (approval No. KYLL-2015(KS)-088) on February 10, 2015.
Key Words: nerve regeneration; progranulin; Atsttrin; neuroinflammation; inflammatory cytokines; lipopolysaccharide; intracerebroventricular injection; astrocyte; nuclear factor kappa B signaling pathway; progranulin knockout mouse; cerebrospinal fluid; neural regeneration
Graphical Abstract

Atsttrin is a potential target for neuroinflammation therapy
Introduction
Neuroinflammation plays a key role in the pathogenesis of neurodegeneration and neuronal loss (Fan et al., 2015) in a variety of neurological diseases, including Alzheimer's disease (AD) (Taylor et al., 2018) and Parkinson's disease (PD)(Festoff et al., 2016). Neuroinflammation also contributes to tissue damage following traumatic injury, and behavioral deficits (Caffes et al., 2015; Chiu et al., 2016). Progranulin has anti-inflammatory effects in diverse physiological processes (Wei et al., 2014; Cerezo et al., 2015; Zhao et al.,2015). It is also linked to a number of neurological disorders,including AD (Carecchio et al., 2009; Jing et al., 2016), PD(Carecchio et al., 2014), frontotemporal lobar degeneration(FTLD) (Karch et al., 2016) and Gaucher disease (Jian et al., 2016). For example, low serum levels of progranulin are found in patients with Gaucher disease (Jian et al., 2016),and progranulin deficiency is considered a major cause of FTLD (Karch et al., 2016). Plasma progranulin level was shown to be a useful marker for the diagnosis of AD (Carecchio et al., 2009). Furthermore, mutations in the progranulin gene (GRN) are associated with a wide variety of clinical phenotypes, including Parkinsonism (Carecchio et al., 2014).Recently, Atsttrin, an engineered protein composed of three progranulin fragments, has been shown to have anti-inflammatory actions in many disease models (Jian et al., 2016).Most importantly, progranulin and Atsttrin were found to have similar effects on the progression of inflammatory diseases (Liu, 2011; Tang et al., 2011; Wang et al., 2015).Atsttrin was shown to significantly attenuate dermatitis in a mouse model (Zhao et al., 2013) and curtail disease development in a mouse model of arthritis (Wang et al., 2015;Wei et al., 2017). Additionally, various studies have indicated that Atsttrin interacts with the nuclear factor kappa B (NFκB) signaling pathway (Zhao et al., 2013; Wei et al., 2017).However, whether Atsttrin suppresses the development of neuroinflammation remains unknown.
The NF-κB family includes important transcription factors involved in diverse cellular processes (Yan et al., 2016). NFκB plays key roles in neurological diseases as well. For example, NF-κB is upregulated and controls the abundance and variety of microRNAs in AD brain tissue (Zhao et al., 2014).Interestingly, progranulin deficiency results in hyperactivation of the NF-κB signaling pathway in lymphoblasts from patients with FTLD (Alquézar et al., 2016). Furthermore,in mouse models of osteoarthritis and dermatitis, Atsttrin suppresses the NF-κB signaling pathway, delaying disease progression (Zhao et al., 2013; Wei et al., 2017). In the present study, we use a lipopolysaccharide (LPS)-induced neuroinflammatory mouse model as well as cultured astrocytes stimulated with LPS to examine the effects of Atsttrin on neuroinflammation and to clarify the underlying molecular mechanisms.
Materials and Methods Animals
The animal experiments were performed in compliance with the institutional guidelines issued by the Animal Care and Use Committee of Shandong University, China. The studies followed the policies of the Animal Management Rules of the Chinese Ministry of Health (document No. 55, 2001).This study was approved by the Animal Ethics Committee of Qilu Hospital of Shandong University, China (approval No.KYLL-2015(KS)-088) on February 10, 2015.
Twenty-eight C57BJ/6 male wild-type (WT) clean mice(20-25 g, 8 weeks old) were purchased from the Animal Center of Shandong University, China (license No. SYXK(Lu) 2019-0005) and 25 male clean progranulin knockout(Grn-/-) mice, of similar weight and age, were obtained from Jackson Laboratories (West Grove, PA, USA). Moreover, 20 newborn mice were purchased from the Animal Center of Shandong University, China (license No. SYXK (Lu) 2019-0005). Brain tissues from these newborn mice were used for the in vitro experiment. All animals were housed in an environment with the same light, temperature (20 ± 2°C) and humidity.
LPS-induced neuroinflammation mouse model
First, to detect levels of progranulin in mouse brain, 18 WT mice were randomly divided into the following two experimental groups: (1) intracerebroventricular (ICV) injection of normal saline (NS) (n = 9) and (2) ICV injection of LPS (n =9). A 1-µL volume of NS or LPS (10 µg/µL; Solarbio, Beijing,China) was administered via ICV injection according to a published protocol with minor modification (Lawson et al.,2013). Vital signs were measured every 12 hours, one day after injection to ensure that all mice survived. Animals in each group (n = 3) were euthanized 0, 24 and 72 hours after injection, and progranulin was detected by immunohistochemistry. To assess the levels of progranulin-related cytokines in the brain, WT (n = 10) and Grn-/-(n = 10) mice were randomly divided into two groups (n = 5 each) and given the same ICV injections. Animals were euthanized 24 hours after injection (Cheng et al., 2017). A total of 15 Grn-/-mice were used to examine the therapeutic effect of Atsttrin in neuroinflammation. These mice were randomly divided into three groups: normal control, LPS treatment, and LPS with Atsttrin treatment (n = 5 each). Mice in the LPS treatment group were administered 1 µL LPS (10 µg/µL) alone. In the LPS with Atsttrin treatment group, Atsttrin (2.5 mg/kg; Sangon Biotech,Shanghai, China) was administered via intraperitoneal injection every 3 days for 7 days (Zhao et al., 2013; Schaafsma et al., 2015) before ICV injection of 1 µL LPS (10 µg/µL). All 15 knockout mice were euthanized 1 day after LPS administration. Brain tissue samples were promptly collected and stored at -80°C until analysis.
Murine cerebrospinal fluid collection
Cerebrospinal fluid was collected from Grn-/-mice according to previously published protocols (Barten et al., 2011; Liu et al., 2012; Lim et al., 2018) 1 day after LPS ICV injection.Briefly, a midline sagittal incision on the skull surface was made after anesthesia to expose the cisterna magna, followed by slow insertion of a microneedle. Finally, the needle was removed, and approximately 8-10 µL of cerebrospinal fluid was collected with a pipette within 6 minutes and diluted 1:50 before assay (Liu et al., 2012; Tucker et al., 2015).
Immunohistochemistry
Immunohistochemistry was performed according to a previously published protocol, with modification (Jian et al.,2016). Mouse brain tissue was cut into 5-µm-thick paraffin sections and incubated with primary antibodies (rabbit anti-progranulin and goat anti-tumor necrosis factor-α (TNF-α),1:100, Santa Cruz Biotechnology, Dallas, TX, USA; rabbit anti-inducible nitric oxide synthase (iNOS) and rabbit anti-interleukin-1β (IL-1β), 1:150, Abcam, Cambridge, MA, USA;rabbit anti-matrix metalloproteinase-3 (MMP-3), 1:100, Abcam; rabbit anti-phospho-NF-κB inhibitor α (p-IκBα), 1:100,Abcam) at 4°C for 12 hours. After incubating with secondary antibodies (goat anti-rabbit, 1:200, Abcam; rabbit anti-goat,1:150, Jackson ImmunoResearch, PA, USA) at 37°C for 1 hour, the sections were stained with a diaminobenzidine kit(ZSGB-Bio, Beijing, China), and observed with an IX-81 microscope (Olympus, Beijing, China) and quantified using Image-Pro Plus 6.0 (Media Cybernetics, Rockville, MD, USA).
Isolation and culture of astrocytes
Primary astrocyte cultures were prepared from cortices of newborn mice as previously described with minor modification (Tarassishin et al., 2014). Dulbecco's modified Eagle medium/F-12 (HyClone, Northbrook, IL, USA), containing 1% penicillin-streptomycin solution (Solarbio) and 10% fetal bovine serum (Gibco, Grand Island, NY, USA), was used to culture primary cells. When adherent cells reached 90% confluence, the cultures were shaken in an incubator (250 r/min)overnight to minimize microglial contamination. After three passages, the purity of the astrocyte cultures was 95%.
Drug treatment of astrocytes
To observe how the expression of progranulin were affected by LPS, the astrocytes were stimulated with LPS (0, 100, and 300 ng/mL) for 48 hours. To assess whether inflammatory cytokine levels were affected by Atsttrin, cells were treated with LPS (100 ng/mL), with or without Atsttrin (200 ng/mL) for 24 hours simultaneously. All samples were stored at -20°C and tested within 1 month.
Western blot assay
Protein was extracted from astrocytes according to a previously published protocol (Tarassishin et al., 2014) using a protein extraction kit (Boster, Wuhan, China). Sodium dodecyl sulfate-polyacrylamide gel (10%) electrophoresis was used to separate proteins. Membranes were incubated with polyclonal primary antibodies (rabbit anti-progranulin,1:1000, Santa Cruz Biotechnology; rabbit anti-IκBα, 1:1000,Abcam; rabbit anti-phospho-IκBα (p-IκBα), 1:1000, Abcam)overnight at 4°C. Glyceraldehyde-3-phosphate dehydrogenase (rabbit, 1:5000; Proteintech, Chicago, IL, USA) and LaminB1 (rabbit, 1:3000; Proteintech) were used as internal controls. After incubation with secondary antibody (goat anti-rabbit, 1:2000; Abcam) at room temperature for 1 hour,the membranes were observed on a GE Amersham Imager 600 (General Electric, Boston, MA, USA) and quantified using Image-Pro plus 6.0 (Media Cybernetics).
Immunofluorescence staining
Astrocytes were cultured on coverslips in 24-well plates, and the cells were stimulated with 100 ng/mL LPS. After LPS stimulation, 50% of the cultures were treated with 200 ng/mL Atsttrin for 24 hours (Wei et al., 2017). Immunofluorescence was performed following a published protocol with minor modification (Tarassishin et al., 2014). Primary antibodies(rabbit anti-cyclooxygenase-2 (COX-2) and rabbit anti-i-NOS, 1:100, Abcam) were added to the cells and incubated overnight at 4°C. The cells were then incubated with Alexa Fluor 488-conjugated goat anti-rabbit secondary antibody(1:300, Cell Signaling Technology, Danvers, MA, USA) at 37°C for 1 hour, followed by incubation with 4′,6-diamidino-2-phenylindole for 5 minutes. An IX-81 fluorescence microscope was used to capture images.
Enzyme-linked immunosorbent assay
The cell culture supernatants were collected after stimulation with LPS for 24 hours. The levels of TNF-α and IL-1β in the cell culture supernatant and cerebrospinal fluid were measured with mouse TNF-α and IL-1β enzyme-linked immunosorbent assay (ELISA) kits (Bluegene, Shanghai, China)following the manufacturer's instructions.
Real-time reverse transcription-polymerase chain reaction
Astrocytes and mouse brains were collected in TRIzol reagent(Takara Bio, Shiga-ken, Japan), and total RNA was prepared according to the manufacturer's protocol. Total RNA was used for reverse transcription using a reverse transcription kit (Toyobo Life Science, Osaka, Japan). Primers used for RTPCR were designed to generate products between 100 and 200 bp in length. Sequence-specific primers were as follows:5′-GCA GGG AGG AGA GTG ATT TGA-3′ and 5′-CCG CCT ACC TCA GTT TCC ATT-3′ for progranulin; 5′-TAC AAG CTG GCT GGT GGG GA-3′ and 5′-GTC GCG GGT CTC AGG ACC TT-3′ for NF-κB2; 5′-GGT TCC TTT GTG GCA CTT G-3′ and 5′-TTC TCT TGG TGA CCG GGA G-3′for TNF-α; 5′-AAT CTC ACA GCA GCA CAT CA-3′ and 5′-AAG GTG CTC ATG TCC TCA TC-3′ for IL-1β; 5′-AAA ACT GAT GCG TGA AGT GCT G-3′ and 5′-AAA ACT GAT GCG TGA AGT GCT G-3′ for COX-2; 5′-GAT GAA CGA TGG ACA GAG GAT G-3′ and 5′-TGT GGA GGA CTT GTA GAC TGG-3′ for MMP-3; 5′-ACC CCT GTC TTC CAC CAG GAG TTG AA-3′ and 5′-TGC AGC CAT GAC CTT TCG CAT TAG CAT GG-3′ for i-Nos. reverse transcription-polymerase chain reaction was performed using SYBR Green PCR Master Mix (Toyobo Life Science). Glyceraldehyde-3-phosphate dehydrogenase was used as an internal control to normalize target gene expression. The relative levels of the target gene were calculated using the 2-ΔΔCTmethod.
Statistical analysis
The data are presented as the mean ± standard deviation(SD). One-way analysis of variance (where appropriate) and Dunnett's test were used to compare data across all groups.All statistical analyses were performed using GraphPad Prism Software (v6.01, GraphPad Software, La Jolla, CA,USA) and Image-Pro Plus 6.0. A P-value < 0.05 was regarded as significant.
Results
Protein and mRNA levels of progranulin are increased in LPS-induced neuroinflammation both in vivo and in vitro
Immunohistochemistry was performed to assess whether progranulin expression was increased in mice with LPS-induced neuroinflammation. Figure 1A-C shows that progranulin expression was increased after ICV LPS injection compared with NS injection. A significant increase in progranulin expression was found at 24 hours, compared with 0 and 72 hours (P < 0.01). Figure 1D shows that the relative levels of progranulin mRNA were upregulated after LPS injection. Additionally, as shown in Figure 2A, greater than 95% of the cultured cells were astrocytes as assessed by glial fibrillary acidic protein (GFAP) labeling. Figure 2B and C shows that the expression of progranulin increased markedly after stimulation with 100 and 300 ng/mL LPS for 48 hours,compared with control (P < 0.001). A significant increase in mRNA expression of progranulin in astrocytes was observed after administration of different concentrations of LPS(Figure 2D). These results indicate that treatment with LPS increases progranulin expression both in brain tissues and in cultured astrocytes.
LPS increases expression of pro-inflammatory factors in progranulin knockout mice
Accumulating studies show that progranulin suppresses inflammation in osteoarthritis and inflammatory bowel diseases (Wei et al., 2014; Zhao et al., 2015). A previous report revealed that some pro-inflammatory cytokines are increased 24 hours after LPS ICV administration (Cheng et al., 2017). Therefore, in this study, immunohistochemistry was used to detect pro-inflammatory factors 24 hours after LPS or NS ICV injection (Figure 3A-F). As shown in Figure 3A and B, in the Grn-/-mice, the levels of TNF-α were higher after LPS treatment, compared with NS (P < 0.001).Furthermore, TNF-α expression was significantly increased in Grn-/-mice compared with WT mice 24 hours after LPS injection (P < 0.001) (Figure 3B). There was a significant increase in IL-1β in Grn-/-mice compared with WT mice 24 hours after LPS injection (P < 0.01) (Figure 3C and D).Similarly, iNOS expression was higher in Grn-/-mice than in WT mice 24 hours after LPS administration (P < 0.01) (Figure 3E and F). Expression levels of TNF-α, IL-1β and iNOS at 24 hours after NS administration were measured, revealing no significant differences between WT and Grn-/-mice (P> 0.05) (Figure 3B, D and F). Next, RT-PCR was carried out to measure mRNA levels of TNF-α, IL-1β, iNOS and COX-2. As shown in Figure 3G-J, compared with NS, LPS significantly increased the expression of these molecules in brain tissues from Grn-/-mice (P < 0.001). Together, these findings demonstrate that the expression levels of key pro-inflammatory factors are significantly higher in Grn-/-mice than in WT mice 24 hours after LPS administration.
Atsttrin reduces LPS-induced neuroinflammation in progranulin knockout mice
Atsttrin has been shown to attenuate the severity of dermatitis in mice (Zhao et al., 2013). Because a more severe inflammatory response was found in the Grn-/-mice (Figure 3), we used these mice to assess if Atsttrin plays a protective role in the progression of neuroinflammation (n = 5 for each group). Immunohistochemistry, real-time RT-PCR and ELISA were performed to assess cytokine changes. Figure 4A, C, E and G shows the TNF-α, IL-1β, MMP-3 and i-NOS immunoreactivities in the normal control, LPS treatment and LPS with Atsttrin treatment groups. As shown in Figure 4B, D, F and H, compared with the LPS treatment group,the immunoreactivities of TNF-α, IL-1β, MMP-3 and i-NOS were significantly decreased by Atsttrin injection (TNF-α: P< 0.01; IL-1β: P < 0.001; MMP-3: P < 0.01; i-NOS: P < 0.001).As shown in Figure 4I-L, the mRNA levels of the inflammation-associated molecules were also markedly reduced by Atsttrin injection in the Grn-/-mouse [TNF-α (P < 0.001),IL-1β (P < 0.01), MMP-3 (P < 0.05), i-NOS (P < 0.01)]. Figure 4M and N shows that, compared with the LPS treatment group, the expression levels of TNF-α and IL-1β were lower in the LPS with Atsttrin treatment group (TNF-α: P < 0.001;IL-1β: P < 0.001). These results suggest that Atsttrin reduces LPS-mediated neuroinflammation in this mouse model.
Atsttrin suppresses inflammatory reactions induced by LPS in vitro
Atsttrin was previously demonstrated to suppress TNF-αmediated inflammation in chondrocytes (Wei et al., 2017).Here, we assessed whether Atsttrin antagonizes the effects of LPS in vitro. Immunofluorescence revealed that COX-2 and i-NOS production induced by 100 ng/mL LPS was downregulated by the addition of 200 ng/mL Atsttrin (Figure 5A and B). The addition of Atsttrin also lowered TNF-α levels in the LPS-treated cultures (P < 0.001) (Figure 5C).Figure 5D shows that the supernatant concentrations of IL-1β were also lowered by Atsttrin, compared with LPS treatment alone (P < 0.05). Furthermore, as Figure 5E-G shows, the addition of Atsttrin significantly diminished the LPS-induced increases in MMP-3 (P < 0.01), COX-2 (P <0.05) and iNOS (P < 0.001). These results suggest that Atsttrin suppresses the secretion of pro-inflammatory cytokines in LPS-stimulated astrocytes.
Atsttrin suppresses the LPS-induced activation of the NF-κB signaling pathway both in progranulin knockout mice and in astrocyte cultures
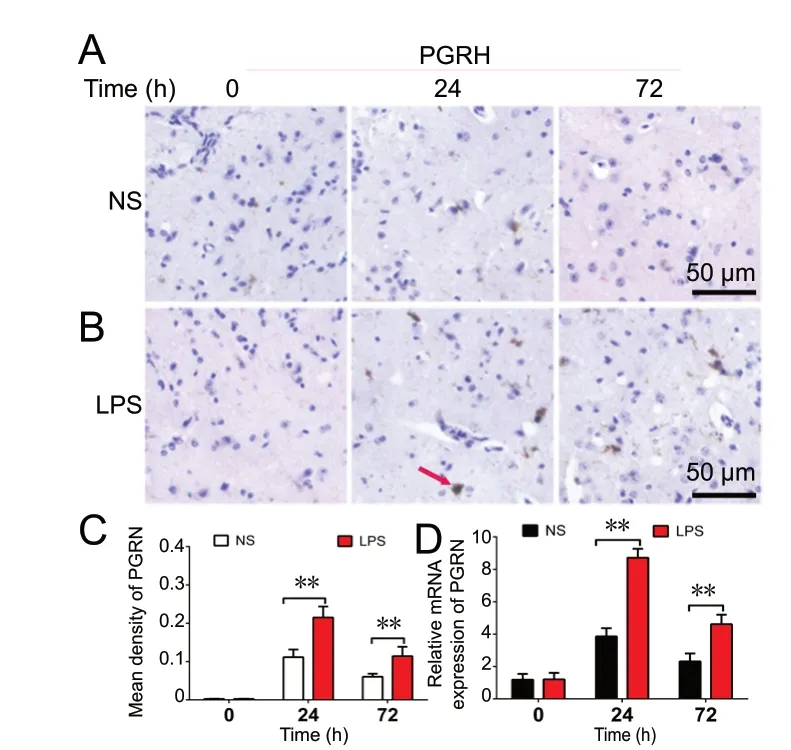
Figure 1 Progranulin (PGRN) is upregulated in neuroinflammation in vivo.(A-C) Immunoreactivity of PGRN (arrow) in brain tissues. Eighteen mice were sacrificed, and a lipopolysaccharide (LPS)-induced mouse model of neuroinflammation was produced. PGRN was detected by immunohistochemistry at 0, 24 and 72 hours (h) after intracerebroventricular injection of 1 µL of LPS (10 µg/µL)or normal saline (NS) (n = 3 for each group). Original magnification, 200×. Scale bars: 50 µm. (D) Real-time reverse transcription-polymerase chain reaction analysis of PGRN mRNA expression in brain tissue after LPS injection. Data are expressed as the mean ± SD. **P < 0.01(one-way analysis of variance followed by Dunnett's test).LPS: Lipopolysaccharide.
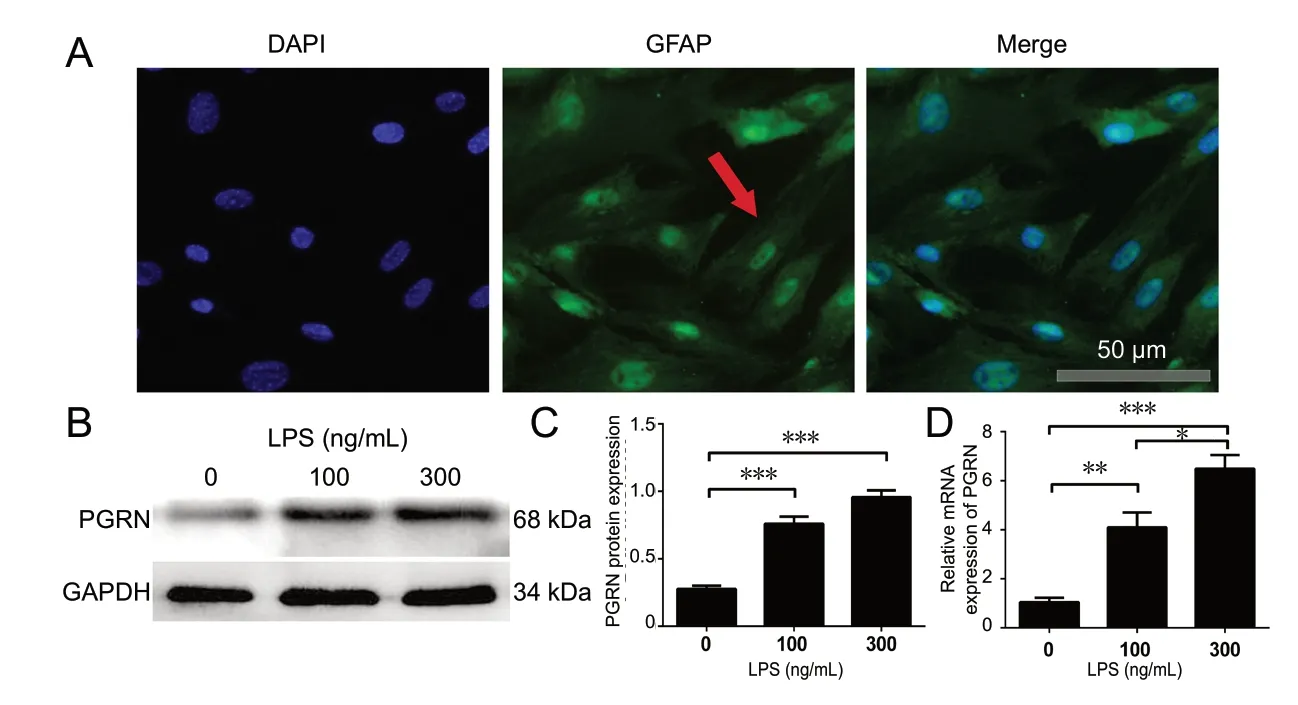
Figure 2 Protein and mRNA expression levels of progranulin (PGRN) are increased in astrocytes by LPS.(A) Immunofluorescence was used to identify astrocytes (arrow). Astrocytes were stained for glial fibrillary acidic protein (GFAP), and nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI). Scale bar: 20 µm. (B, C) Western blot assay for PGRN in astrocytes after LPS treatment (0, 100 or 300 ng/mL) for 48 hours. (D) Real-time reverse transcription-polymerase chain reaction was used to measure mRNA levels of PGRN in astrocytes. The relative expression levels of the target gene were calculated using the 2-ΔΔCT method. Data are expressed as the mean ± SD. *P < 0.05, **P < 0.01,***P < 0.001 (one-way analysis of variance followed by Dunnett's test). GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; LPS: lipopolysaccharide.

Figure 3 Expression levels of pro-inflammatory biomarkers are markedly increased in progranulin knockout (PGRN-/-) mice.Brain tissues were collected 24 hours after intracerebroventricular injection of 1 µL normal saline (NS) or 1 µL LPS (10 µg/µL) for immunohistochemistry and real-time reverse transcription-polymerase chain reaction (n = 5 for each group). (A-F) Immunohistochemistry was used to detect LPS-induced inflammatory cytokines (TNF-α, IL-1β and iNOS) (arrows). Scale bars: 50 µm. (G-J) The mRNA levels of TNF-α, IL-1β, i-NOS and COX-2 in the brain tissues of NS- and LPS-treated mice, assayed by real-time reverse transcription-polymerase chain reaction. The relative levels of the target gene were calculated using the 2-ΔΔCT method. Data are expressed as the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 (one-way analysis of variance followed by Dunnett's test). PGRN: Progranulin; TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β; iNOS: inducible nitric oxide synthase; COX-2: cyclooxygenase-2; LPS: lipopolysaccharide.
As previously reported, Atsttrin suppresses inflammation in a dermatitis mouse model through the inhibition of the NF-κB signaling pathway (Zhao et al., 2013), leading us to evaluate here whether Atsttrin and the NF-κB signaling pathway interact during the neuroinflammatory process.As Figure 6A and B show, Atsttrin significantly decreased the levels of p-IκBα in the brain tissue of LPS-treated Grn-/-mice. Additionally, primary astrocytes were stimulated with LPS (100 ng/mL) for 2 hours, with or without pretreatment with 200 ng/mL Atsttrin simultaneously. Figure 6C shows that Atsttrin markedly decreased p-IκBα levels. As shown in Figure 6D, real-time RT-PCR showed that, in astrocytes, the increased NF-κB2 expression caused by LPS was significantly suppressed by Atsttrin (P < 0.01). In summary, Atsttrin may inhibit inflammatory reactions by repressing the NF-κB signaling pathway both in vivo and in vitro.

Figure 4 Atsttrin suppresses the production of inflammatory mediators in the brain and cerebrospinal fluid in an inflammatory mouse model.(A-H) Atsttrin reduces expression of TNF-α, IL-1β, MMP-3 and iNOS (arrows) in the brain of LPS-injected progranulin knockout (PGRN-/-)mice, as indicated by immunohistochemistry.Representative images from normal control(Cont), LPS treatment (LPS) and LPS with Atsttrin (LPS + Atsttrin) treatment groups. Mice were administered 1 µL LPS (10 µg/µL), with or without Atsttrin (2.5 mg/kg), by intraperitoneal injection every 3 days over a period of 7 days. Brain tissues were harvested for immunohistochemistry(n = 5 for each group). Scale bars: 50 µm. (I-L)Real-time RT-PCR shows that Atsttrin treatment reduced TNF-α, IL-1β, MMP-3 and iNOS mRNA levels in LPS-treated PGRN-/- mice. The relative levels of the target gene were calculated using the 2-ΔΔCT method. (M, N) Atsttrin decreased circulating levels of TNF-α and IL-1β in LPS-treated PGRN-/- mice, as detected by enzyme-linked immunosorbent assay (ELISA). Cerebrospinal fluid was collected from normal control, LPS treatment and LPS with Atsttrin treatment groups, and ELISA was performed for TNF-α and IL-1β. Data are expressed as the mean ± SD. *P < 0.05, **P < 0.01,***P < 0.001 (one-way analysis of variance followed by Dunnett's test). TNF-α: Tumor necrosis factor-α; IL-1β: interleukin-1β; MMP-3: matrix metalloproteinase-3; i-NOS: inducible nitric oxide synthase; LPS: lipopolysaccharide.
Discussion
Neuroinflammation is a protective response to acute brain injury; however, an excessive inflammatory response can promote the development of neurological diseases (Ellwardt and Zipp, 2014; Chiu et al., 2016; Russo and McGavern,2016). For example, neuroinflammation plays a key role in the development of multiple sclerosis through immune effector mechanisms (Ellwardt and Zipp, 2014). It also plays a critical pathogenetic role in AD and PD by altering the expression of protease-activated receptor-2 (Hurley et al.,2015). LPS induces neuroinflammation in the central nervous system and is extensively used to model neuroinflammation (Lawson et al., 2013; Russo and McGavern, 2016).For example, after the administration of LPS, notably higher expression levels of multiple pro-inflammatory cytokines are found in rodent models of PD (Ramsey and Tansey, 2014).These cytokines, in turn, promote the development of neurological diseases.
Progranulin is a multi-functional molecule. In arthritic diseases such as osteoarthritis, progranulin attenuates the breakdown of physiological cartilage structures and slows disease progression in an osteoarthritis mouse model (Zhao et al., 2015). Progranulin has been demonstrated to exert protective functions in neurological diseases as well. Previous research has shown that recombinant progranulin is therapeutic against Gaucher disease (Jian et al., 2016). Here,we found high expression levels of progranulin in vivo in an LPS-induced neuroinflammatory mouse model (Cheng et al., 2017). Moreover, we found a similar increase in the expression of progranulin in LPS-stimulated astrocyte cultures. These findings led us to suspect that progranulin plays a protective role in neuroinflammation. Pro-inflammatory cytokines are expressed most highly 24 hours after LPS administration. In the current study, the levels of inflammatory molecules were examined 24 hours after LPS stimulation in progranulin knockout mice. We found that TNF-α, IL-1β,iNOS and COX-2 were markedly increased in these Grn-/-mice, suggesting that progranulin deficiency exacerbates neuroinflammation.
Recombinant progranulin has been shown to protect against the deterioration of brain tissue in Grn-/-mice (Menzel et al., 2017), and was found to be therapeutic in a mouse model of Gaucher disease (Jian et al., 2016). Atsttrin is an engineered protein derived from progranulin that has been extensively investigated (Liu, 2011; Tang et al., 2011; Zhao et al., 2013; Liu et al., 2014; Wang et al., 2015). In mouse and rat models, Atsttrin protects against early osteoarthritis by protecting cartilage tissues against injury, and alleviates articular pain caused by osteoarthritis (Wei et al., 2017). Atsttrin has an anti-inflammatory effect similar to that of progranulin in diverse diseases. For example, progranulin and Atsttrin both effectively ameliorate inflammation in a mouse model of colitis (Liu et al., 2014; Wang et al., 2015). Furthermore,progranulin and Atsttrin reportedly antagonize TNF-α and protect against inflammatory arthritis (Liu, 2011; Tang et al.,2011; Wang et al., 2015). In the present study, we found that in Grn-/-mice, LPS stimulation increased pro-inflammatory cytokine levels. Furthermore, Atsttrin treatment significantly decreased expression of these cytokines.
Astrocytes are major players in the development of diverse neurodegenerative diseases (Cekanaviciute and Buckwalter,2016). The pathological response of astrocytes includes reactive astrogliosis and the release of interleukins and nitric oxide (Taylor et al., 2018). Mouse astrocytes respond to LPS by secreting pro-inflammatory cytokines (Tarassishin et al.,2014). Astrocyte activation can be induced by LPS (Lawson et al., 2013; Ramsey and Tansey, 2014). Here, we found that Atsttrin dramatically suppressed the expression of inflammatory mediators in astrocytes, consistent with the in vivo findings. These results suggest that Atsttrin may have therapeutic potential in the treatment of neuroinflammation.
The NF-κB signaling pathway is pivotal during the inflammatory process (Yan et al., 2016). NF-κB signaling causes mitochondrial dysfunction by repressing adenine nucleotide translocase type 1 gene transcription in neurodegenerative diseases such as AD and PD. LPS activates the NF-κB signaling pathway in astrocytes to produce pro-inflammatory mediators. Both progranulin and Atsttrin play an anti-inflammatory role in many diseases, including osteoarthritis,rheumatoid arthritis and dermatitis, by inhibiting NF-κB signaling (Liu, 2011; Tang et al., 2011; Zhao et al., 2013). This prompted us to examine whether Atsttrin affects the NFκB signaling pathway in LPS-induced neuroinflammation.We observed here that p-IκBα levels are reduced by Atsttrin treatment. Together, these findings show that Atsttrin significantly suppresses the NF-κB signaling pathway and impacts the levels of its downstream effectors.
Progranulin alleviates inflammatory arthritis and colitis in mice by binding to TNF receptors (TNFRs) (Tang et al.,2011; Wei et al., 2014; Wang et al., 2015; Zhao et al., 2015).TNF-α mediates a variety of inflammatory responses, and in neurodegenerative disorders such as AD, amyotrophic lateral sclerosis and multiple sclerosis, TNFR1 is activated upon binding soluble TNF (Tang et al., 2011; Wei et al.,2014; Wang et al., 2015; Zhao et al., 2015). Atsttrin binds TNFR (Liu, 2011; Tang et al., 2011; Wang et al., 2015) and inhibits inflammation through its interaction with TNFR1 and TNFR2 in an arthritis mouse model (Liu, 2011; Tang et al., 2011; Wei et al., 2017). Further study is needed to investigate whether Atsttrin also alleviates inflammation in humans. Nonetheless, our findings suggest that Atsttrin may have therapeutic potential for the treatment of neurological diseases associated with neuroinflammation.
Acknowledgments: The authors would like to greatly thank the Cardiovascular Laboratory of Shandong University, China for their technical support, which was fundamental to the realization of this work.
Author contributions:Experiment design: LL, LC; study authorization and instruction: LC; experiment implementation and data analysis: LL,YQ, YL, HZ, HCM, CJJ, LN; manuscript writing: LL; manuscript revising: AFN, MS. All authors approved the final version of the manuscript.
Conflicts of interest:None declared.
Financial support:This work was supported by the National Natural Science Foundation of China, Nos. 81572191 (to LC) and 81601067 (to HZ). The funders did not participate in data collection and analysis, article writing or submission.
Institutional review board statement:This study was approved by the Animal Ethics Committee of Qilu Hospital of Shandong University of Science and Technology, China (approval No. KYLL-2015(KS)-088) on February 10, 2015.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.

Figure 5 Atsttrin inhibits the LPS-induced inflammatory reaction in astrocytes.Astrocyte cultures were prepared from newborn wild-type mouse brain and cultured with 100 ng/mL LPS for 24 hours, in the presence or absence of 200 ng/mL Atsttrin. (A, B) Immunostaining was performed for COX-2 and i-NOS (arrows; Alexa Fluor 488), and 4′,6-diamidino-2-phenylindole was used to stain nuclei. Scale bars: 20 µm. (C, D) Supernatant levels of TNF-α and IL-1β in LPS-treated astrocytes, measured by ELISA. (E-G) The LPS-induced increase in pro-inflammatory factors, including MMP-3, COX-2 and i-NOS, was attenuated by Atsttrin, as analyzed by real-time RTPCR. The relative expression level of the target gene was calculated using the 2-ΔΔCT method. Data are expressed as the mean ± SD. *P < 0.05, **P< 0.01, ***P < 0.001 (one-way analysis of variance followed by Dunnett's test). COX-2: Cyclooxygenase-2; i-NOS: inducible nitric oxide synthase;TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β; MMP-3: matrix metalloproteinase-3; LPS: lipopolysaccharide.
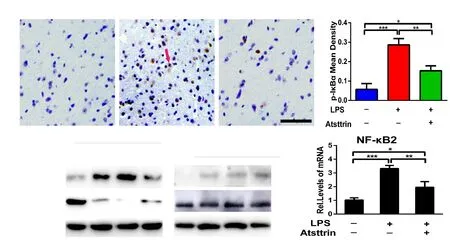
Figure 6 Atsttrin inhibits the LPS-induced activation of the nuclear factor kappa B (NF-κB) signaling pathway.(A, B) Atsttrin attenuated the increase in the expression of p-IκBα (arrow) in the brain tissues of LPS-treated progranulin knockout mice, as assessed by immunohistochemistry. Mice received the same treatment as in Figure 3. Scale bar: 50 µm. (C) Western blot assay showing that Atsttrin inhibits the LPS-induced activation of the NF-κB signaling pathway. Astrocytes were cultured with 100 ng/mL LPS, with or without 200 ng/mL Atsttrin simultaneously, for 2 hours. Lamin B1 served as an internal control. (D) Cultured astrocytes were treated with 100 ng/mL LPS for 24 hours, with or without 200 ng/mL Atsttrin, and NF-κB2 levels were measured with real-time RT-PCR. The LPS-induced increase in NF-κB2 levels was significantly diminished by Atsttrin. The relative expression level of the target gene was calculated using the 2-ΔΔCT method. Data are expressed as the mean ± SD. *P < 0.05, **P <0.01, ***P < 0.001 (one-way analysis of variance followed by Dunnett's test). IκBα: NF-κB inhibitor α; p-IκBα: phospho-IκBα; LPS: lipopolysaccharide.
Open peer reviewer:Cristoforo Comi, University of Piemonte Orientale,Italy.
Additional file:Open peer review report 1.

- 中国神经再生研究(英文版)的其它文章
- Etomidate affects the anti-oxidant pathway to protect retinal ganglion cells after optic nerve transection
- Pain inhibition through transplantation of fetal neuronal progenitors into the injured spinal cord in rats
- MicroRNA expression in the hippocampal CA1 region under deep hypothermic circulatory arrest
- Association between PPARG genetic polymorphisms and ischemic stroke risk in a northern Chinese Han population: a case-control study
- Silencing Huwe1 reduces apoptosis of cortical neurons exposed to oxygen-glucose deprivation and reperfusion
- Paired associative stimulation improves synaptic plasticity and functional outcomes after cerebral ischemia