
林下凋落物对土壤种子萌发的作用机制
2019-07-08 03:55周清慧黄冠陈继豪王鹏程晏召贵
湖北林业科技 2019年3期
周清慧 黄冠 陈继豪 王鹏程 晏召贵
摘 要:林下土壤种子库是森林植被的物种组成、种群延续和种群演替的物质基础。林下凋落物影响土壤种子库种子的萌发,幼苗的生长,进而影响到种群更新及森林群落的演替。凋落物对种子萌发的影响主要有三大作用机制,即物理作用机制化学作用机制、和生物作用机制。其中化学作用机制特别是化感作用对土壤种子库的影响尤其重要。本文将从这三个作用机制的层面,分析总结前人有关土壤种子库种子的萌发及其与林下凋落物、林下植被多样性的研究,论述土壤种子库研究面临的挑战,并对土壤种子库与生物多样性的研究进行了相关展望。
关键词:土壤种子库;凋落物;林下植被多样性
中图分类号:S753 文献标识码:A 文章编号:1004-3020(2019)03-0001-06
Abstract:Soil seed bank of forest is the material basis of species composition, population growth and community succession for forest vegetation. Litter of forest understory affects seed germination, seedling growth, and population growth, thus affecting forest community succession. Litter affect soil seed bank through one of the three mechanisms, physical impact, chemical reaction, and biological interaction. Chemical reactions, especially allelopathy, play important roles in seed germination and seedling growth. In this paper, we analyzed and summarized previous studies on soil seed bank, litter and understory species diversity. We also discussed existing problems and future challenges on research of soil seed bank, litter and forest understory biodiversity.
Key words:soil seed bank;litter;biodiversity of undergrowth
1 引言
土壤種子库(soil seed bank)是指存在于土壤上层凋落物和土壤中全部种子的总和。本文讨论的森林土壤种子库即指森林土壤中存活的种子[1]。土壤种子库承接和储藏地表植被和周边植物种子,是植被天然更新和系统恢复的物质基础[2-3]。同时,土壤种子库时期也是植物种群生活史的一个潜种群阶段,研究种子库构成可以预测森林植物群落的再生潜力,反映植物群落动态演变特征土壤种子库目前的主要研究内容包括种子库的数量有效性及其动态、种子库与群落更新、种子库与生态演替、种子库与植被恢复重建、种子库的分布格局与其生物学特性的关系及种子库与其地上植被之间的关系[4-5]。种子库组成不仅取决于群落的物种组成和结构特征,还受群落环境特征影响。所以,种子库对生物多样性研究和生态恢复重建也具有重要意义[6]。
目前,中国有关森林凋落物对森林更新的研究起步于20世纪中叶,半个多世纪以来这一领域取得了长足的发展。国内最早的森林凋落物影响种子萌发与幼苗生长的报道始于20世纪60年代初。陶大立等(1985)在小兴安岭丰林自然保护区进行了有关死、活地被物(凋落物)对红松Pinus koraiensis伴生树种天然更新影响的研究,初步探讨了森林凋落物对种子萌发和幼苗生长的影响[7]。而后,又陆续有相关的科学研究证明不同森林生态系统森林凋落物的现存量和分解动态存在显著差异[8-9],从而导致土壤种子库的种子萌发与幼苗生长动态差异。国内的绝大部分研究还是从森林凋落物影响土壤种子库种子萌发与幼苗生长的单一机制入手,缺乏对于各机制协同影响森林更新的研究。本文将从物理、化学、生物及生物化学等方面,分析林下凋落物对于土壤种子库种子的萌发与更新的影响及作用机制。
2 凋落物对土壤种子库的影响
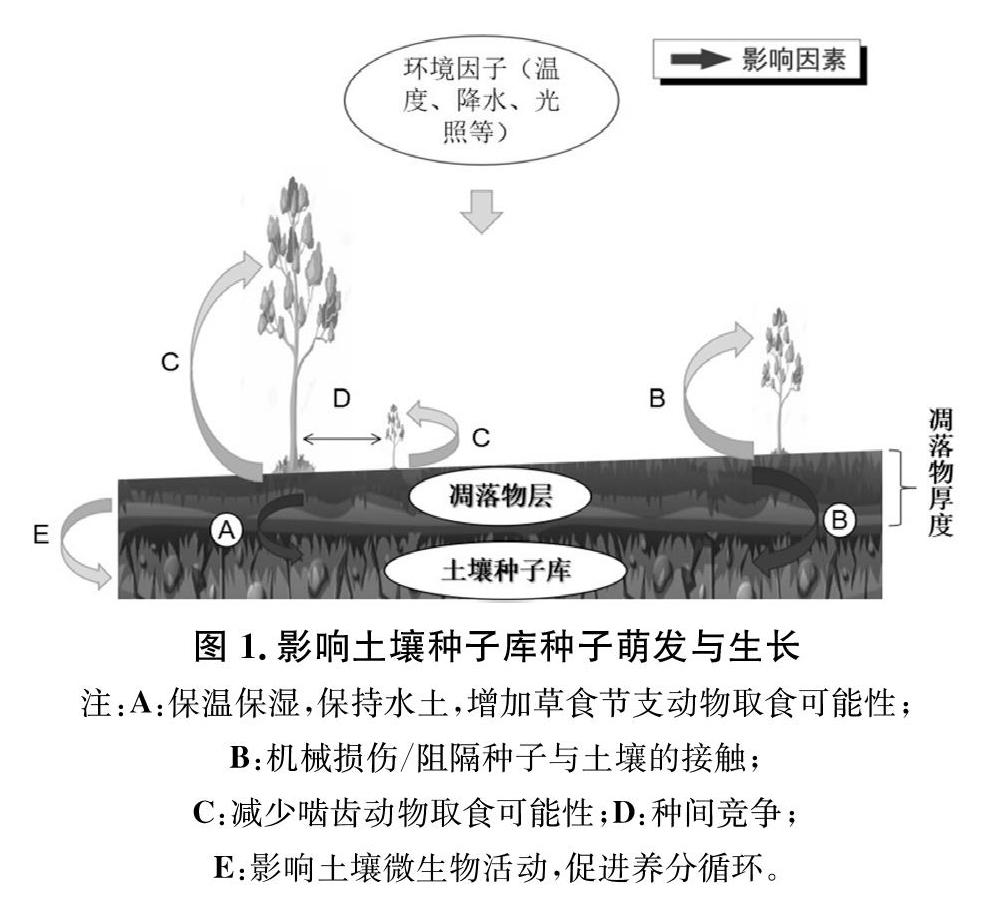
林下凋落物对种子库的影响主要包括影响土壤温湿度,水土保持,影响土壤中微生物活动,促进土壤养分的循环,进而影响土壤种子的萌发以及幼苗的生长(图1)。凋落物的分解过程,是一个包括土壤微生物、土壤物理、及土壤化学的综合过程。在凋落物分解过程中,各种营养元素,包括N,P,K及其它微量元素被释放,不仅改良了土壤的营养状况,同时,木质素,纤维素的分解,也直接改善了土壤的物理结构,从而为土壤种子库的萌发与生长奠定了基本的养分条件和相对完善的生存空间。
凋落物的覆盖,可使土壤种子库长期处于高湿、保温、荫蔽的环境下,从而影响到土壤种子库所处的光环境,对于一些需要在避光条件下才能萌发的种子来说,凋落物为其创造了一个荫蔽的环境,从而使其顺利萌发。凋落物在这些方面对土壤环境的改变,会对土壤种子库的萌发和幼苗的生长产生积极作用(图1)。
研究结果表明,凋落物量累积的过程就是土壤有机质和全氮积累的过程。凋落物改善了土壤的物理结构,土质疏松,利于幼苗扎根土壤,土壤孔隙度增加,根系的生长需氧就不会再受到限制,土壤保水能力增强,可使植物长期处于适宜的水环境当中,便于植物保持旺盛的生长状况。另外,凋落物的覆盖可在一定程度上起到保温的作用,在冬季和早春较为寒冷或者天气异常的状况下,凋落物可保证种子和萌发幼苗不被冻坏、冻死。凋落物的存在,还会影响到土壤种子库所处的光环境,对于一些需要在避光条件下才能萌发的种子来说,无疑凋落物为其创造了一个荫蔽的环境,从而使其顺利萌发。凋落物在这些方面对土壤环境的改变,会对土壤种子的萌发和幼苗的生长产生积极作用。
另一方面,凋落物的覆盖,可使土壤种子库长期处于高湿、保温、荫蔽的环境下,进而影响到土壤种子的萌发及幼苗的生长。一些休眠时间较长的植物种子会因为高湿,恒温条件而变质、腐烂。恒温的条件下,还会对需要在变温条件下才能破除休眠而萌发的一些植物种子产生不利影响。凋落物的厚度达到一定程度时,就会形成荫蔽的环境,刚萌发的植物幼苗叶片很难进行光合作用,当幼苗养分消耗殆尽也会导致幼苗死亡。
通过对种子萌发和幼苗生长的影响,凋落物对林下植物群落的组成及结构产生重要影响。例如,Persson(1987)报道了森林地表凋落物的组成成分在50年中的变化[10],试验表明植物群落从栎属Quercus和榛属Corylus为主变为以榆属Ulmus及水青冈属Fagus植物为主。由于减少了植林密度,凋落物使植株避免了竞争和提高了生长率,因此,对群落中种的多样性及种类组成产生一定的影响。凋落物的累积可降低种的丰富度。
3 凋落物作用机制
3.1 凋落物对土壤种子库的物理作用机制
凋落物对群落土壤的发育和改良有重要意义,凋落物在不断的积累与分解过程中,能够改善土壤结构,降低土壤容重,增大土壤孔隙度,增大土壤持水能力,增大降水入渗能力,使土壤具有良好的通透性,对保持水土、涵养水源有巨大的作用。首先,在对土壤物理性状的影响上,土壤结构和土壤温度是土壤物理性质的两个重要方面。土壤结构影响植物生长所需的水分与养分供应能力、通气和热量等环境条件,以及根系在土体中的穿透情况。
研究结论有相关报道,证实土壤持水能力的大小与地表覆盖的凋落物层厚度(或蓄积)密切相关[11]。国内对于森林凋落物对土壤种子库种子萌发更新的物理作用的机制研究一般集中在以下两种:一种是,他们分别从凋落物对于光照及水分的通过和渗透程度来表明凋落物通过物理机制的作用对于土壤种子库种子的正向影响。如郭伟等(2009)试验表明对未分解的森林凋落物作为地表的覆盖物是可以提高种子发芽率和幼苗成活率、增加植物的总生物量的,它通过截留降水增加土壤含水量、降低种间竞争以及草本层密度与生物量来实现[12];王俊等(2008)研究也表明了这一点,在种子质量和掩埋位置均相同的条件下,凋落物的覆盖物作用明显提高了幼苗的相对生长率。[JP+1]另一种研究则是说明了森林凋落物降低了幼苗成活率,而这一观点是通过凋落物带来的机械损伤及凋落物导致种子与土壤的接触程度不同来解释凋落物对土壤种子库种子萌发与更新的负向影响[13]。在一定条件下,凋落物不仅可以阻断种子与土壤的接触,还能对新生幼苗产生机械损伤[14-15]。Pierson和Mack(1990)在北美西部森林进行的研究也表明,Bromustectorum种子发芽率与幼苗生物量在凋落物厚度大(6 cm)地区均低于凋落物厚度小(1.5 cm)的地区[16]。刘尚华等(2008)对于京西百花山区的研究也表明了,幼苗的死亡率与凋落物厚度成正相关[17]。
这两种研究观点都表明了凋落物与土壤种子库之间的重要关系,但目前国内基本都为单一林分或单一种的研究,凋落物对土壤种子库的影响的研究当中也缺乏对种间比较的研究;同时,在凋落物对土壤种子库的物理作用机制里,由于生态研究的宏观性,忽略了时空异质性对于土壤种子库的影响。
3.2 化学作用机制
腐殖质层和土壤是植物赖以生存的养分库,在养分循环中起着十分重要的作用。凋落物掉至地表后形成腐殖质层,凋落物分解后将养分归还到土壤中,一方面改善土壤的物理结构,另一方面改善了土壤的肥力状况。其中的养分通过分解等过程释放归还至土壤,并被植物所利用。广义而言,凋落物是土壤有机质的一个组成部分,狭义的土壤有機质主要指土壤腐殖质,而它的主要来源还是植物的凋落物。凋落物的质和量,加上温度、雨量等外界环境因素共同决定了相应土壤中有机质的含量。在森林演替过程中,从先锋植物群落一直到顶极群落,土壤化学性质也在逐渐变化,从总的趋势上看,土壤有机质和全氮含量是不断增加的,这主要是植被演替过程中,凋落物不断积累和分解而对土壤长期作用的结果。已有报道表明[18],森林每年通过凋落物分解归还土壤的总氮量占森林生长所需总氮量的70%~80%,总磷量占65%~80%,总钾量占30%~40%。由此可见,凋落物及其分解在物质循环中占有极为重要的地位,是森林生态系统得以维持的重要因素。另外,土壤的酸碱度也是土壤的重要特性之一,土壤过酸或过碱对植物生长都不利。
3.3 生物作用机制
森林生态系统中凋落物的生物作用机制包含两个相对的过程:生物合成与生物分解。前者是无机物的有机化过程,主要由绿色植物和一些无机营养微生物来完成。后者是有机物的无机化过程,主要由森林微生物来完成,即森林凋落物的分解过程。森林凋落物的分解一般涉及三个步骤:①淋溶作用,通过降水将凋落物中可溶性物质带入土壤;②自然粉碎作用,通过腐食动物的啃食,土壤干湿交替、冻融等使凋落物破碎;③代谢作用,通过微生物的作用将复杂有机物转化为简单无机物。在森林凋落物分解过程中,这三个步骤交互作用,相互促进。
森林凋落物分解的微生物作用过程是一个基于分解者细胞水平的酶解过程,这一过程受环境因素和生物因素的相互作用影响[19]。微生物群落在凋落物残屑中的演替过程,其实质是一个以适应性微生物顺序出现而构成的腐食食物链从落地鲜叶到变色、组织崩溃、消解这一分解过程中,微生物类群的组成和数量基本呈现出由寄生到腐生的演替关系[20]。
3.4 生物化学作用机制
除纯粹的物理化学作用机制外,凋落物的生物化学作用机制,也是影响林下种子库及种子萌发的重要因子。生物化学机制又称化感效应,是指先锋树种分泌物对本身(自感)和其他物种(它感)的繁殖(如种子萌发)和生长的有利或不利的影响。1984年Rice将化感作用定义为"一种植物通过向环境释放化学物质而对另一植物(包括微生物)所产生的直接的或间接的,有利的或有害的作用",这是目前为止使用得最广泛的定义。
植物的化感作用在很多物种及生态系统中都得到验证。May & Ash (1990) 通过实验的方法[21],证明桉树化感物质对一些常见的林下植物(如Lolium,Lemna和Acacia spp.)的种子萌发和生长,有明显的抑制作用[22]。我国学者对天山云杉Picea schrenkiana的研究表明,凋落物的水浸提液对云杉自身的种子的萌发,幼苗的生长,都有极大的抑制作用[23]。Reigosa和González(2006)的研究认为,森林优势树种在种子萌发期间虽不能改变周边环境,但在幼苗生长时期可以通过自身的各种生物化学的作用机制适应并逐渐改变生活环境[25]。化感物质可以通过影响细胞膜透性、细胞分裂生长与分化、呼吸作用、蛋白质合成与基因表达以及改变激素合成与平衡等途径促进或抑制种子萌发[26-29]。大量实验已经揭示了化感物质对植物的伤害机制,但由于化感物质种类繁多,其不同浓度处理下不同植物的反应不同,所以作用机制也各不相同。
一般而言,植物受到高浓度化感物质伤害时往往表现出种子萌发率与速度降低,根和芽的生长速度降低,生物量的累积减少,器官形态也会改变,比如叶片发黄甚至萎蕉、死亡等。陈慧玲等(2013)研究发现低浓度油茶水浸液对桔梗幼苗的生长具有一定的促进作用[30]。而Jenson等使用黑胡桃juglans nigra果皮浸提液处理豌豆根,观察到了幼苗根茎有丝分裂受抑制的现象[31-32]。Muller(1965)报道了从Salvia leucophylla叶片中提取的挥发性物质对黄瓜根尖细胞有丝分裂的抑制,且对根和下化轴细胞的伸长也有明显的抑制作用[33]。蒋有绪(1981)研究发现,新凋落杜鹃叶、栎叶和竹叶浸出液能延长冷杉种子发芽时间,降低冷杉种子发芽势和发芽率[34]。也就是说,大多数化感物质在合适浓度下,是可提高植物对物理环境的适应性,抵御天敌的侵害,增强抗病性,提高物种间的竞争力,维系植物与其他生物的互惠关系,但在高浓度下,会抑制植物生长甚至造成植物死亡。
4 研究展望
当前种子库研究中存在的(也是突破传统研究)最重要的问题是:土壤种子库研究作为种群生态学研究的重要内容,不应仅仅从种群一个层次进行研究,应当从生态学的各个组织层次上进行[35]。微观层次的研究可从基因和个体方面进行,这对濒危植物和非濒危植物的比较生物学和比较生态学的研究和某种优良性状或遗传特征的遗传学研究都有重要意义[36]。另一方面,不同物种的种子萌发及幼苗生长对不同环境因子的响应不同,因而影响其自然更新和种群演替。影响种子萌发和幼苗的生长的直接因子包括光照、温度、土壤水分条件等以及对直接因子有影响的间接因子如坡向、坡位、林分郁闭度、林地凋落物等[37-40]。坡向、坡位通过影响光照条件、土壤水分而影响种子的萌发和幼苗的生长。而林分郁闭度直接影响林下光照,在高郁闭度林分内,由于光照不足,一些需光植物的种子的萌发受到影响甚至完全不能萌发,或萌发后幼苗不能正常生长(张敏等2012;颜安等2014),直接影响到这类植物的林下自然更新[41-42]。
通过大量研究发现,土壤种子库和地上植被的关系大致有2种情况,即不相似性和相似性。大多数研究认为,通常在演替初期,二者相似性较高,随着演替进行,到了演替后期,土壤种子库与地表植被之间存在的差异开始变大,大部分的演替晚期乔木和灌木种类会很少出现在持久种子库中,耐阴性树林的下层植被往往产生相对生命较短的少量的种子,而且不会积累在土壤里[43];如Wagner(2006)等人就发现在次生演替阶段的土壤种子库与地上植被随着演替时间长而相似性减小[44]。大多研究都认为:土壤种子库和地上植被间是具有相关性的。如Leck(1979)对湖水沼澤的研究,Peco(1998)、Manzano(2005)对一年生牧场的研究、Henderson(1988)等对沙漠断草群落、Roberts(1962)对农田、Roach(1983)对草地、刘明宏(2010)对森林的研究,均表明土壤种子库和地上植被间存在相关性[45-51]。其中部分研究者认为,相关性的主要原因是:干扰[52]。在干扰强烈的地方,尤其是一年生植物占优势的区域,土壤种子库与地上植被间的藕合相关性就越强,例如:耕地[53]。也有研究表明:土壤种子库与上植被间不存在显著相关性,如弃耕地土壤种子库和地上植被间随时间而逐渐不同。对常绿阔叶林的土壤种子库和地上植被间物种组成分析表明,演替初期其物种相似度为50%,而在演替其他阶段相似度仅为20%左右,其他对成熟林的研究也表明土壤种子库和地上植被组成成分存在显著差异[54-55]。到目前为止,土壤种库和地上植被间的稱合关系还没有统一的定论,然而土壤种子库和地上植被间的关系仍是土壤种子库的重要研究内容,仍是土壤种子库研究内容的热点之一[56]。
现今,土壤种子库大多采用幼苗出现法进行研究,这种方法在实际操作起来具困难且不可控因素较多,如何简便地开展土壤种子库研究这一问题,也是土壤种子库研究者今后考虑的重要方向。土壤种子库是植物演替的驱动因子却也是现存植物群落的产物,而目前的研究基本都着眼于驱动因子上,而对于两者之间的辩证关系缺乏相应的研究与解释;同时,也缺乏对于不同演替阶段的土壤种子库的差异性与地上植被多样性关系研究;涉及土壤种子库遗传结构的研究比较少的原因可能是所需时间长,或者是各方面的技术实力不足。今后应该加强土壤种子库的基因库和进化记忆功能的研究,为生物多样性的保护提供基础资料。
参 考 文 献
[1]张玲,李广贺,张旭.土壤种子库研究综述[J].生态学杂志,2004,23(2):114-120.
[2]Kebromt,Tesfaye B. The role of soil seed banks in the rehabilitation of degraded hillslopes in Southern Wello Ethiopia[J].Biotropiea,2000,32(1) :23-32.
[3]楊跃军,孙向阳,王保平.森林土壤种子库与天然更新[J].应用生态学报,2001,12(2):304-308.
[4]Bakker J P. Nature management by grazing and cutting[J]. Journal of Applied Ecology, 1990, 5(4):128.
[5]于顺利,蒋高明.土壤种子库的研究进展及若干研究热点[J].植物生态学报,2003,27(4):552-560.
[6]安村青,林向阳,洪必恭.宝华山主要植被类型土壤种子库初探[J].植物生态学报,1996,20(1):41-50.
[7]陶大立,徐振邦,李昕.死、活地被物对红松伴生树种天然更新影响的实验研究[J].植物生态学报,1985,9(1):47-58.
[8]齐泽民,王开运,宋光煜,等.川西亚高山箭竹群落枯枝落叶层生物化学特性[J].生态学报,2004,24(6):1230-1236.
[9]张东来,毛子军,朱胜英,等.黑龙江省帽儿山林区6种主要林分类型凋落物研究[J].植物研究,2008,28(1):104-108.
[10]Persson S, Malmer N, Bo W. Leaf litter fall and soil acidity during half a century of secondary succession in a temperate deciduous forest[J]. Vegetatio, 1987, 73(1):31-45.
[11]杨吉华,柳凯生.山丘地区森林保持水土效益的研究[J].水土保持学报,1993(3):47-52.
[12]郭伟,张健,黄玉梅,等.森林凋落物生态功能研究进展[J].安徽农业科学,2009,37(5):1984-1985.
[13]王俊,杨龙,李丹艳,等.凋落物覆盖及掩埋对不同质量的藜蒴种子萌发和幼苗生长的影响[J].生态环境学报,2008,17(5):1980-1985.
[14]Hitchmough J D . Urban landscape management.[J]. Urban Landscape Management, 1994.
[15]Gillman L N , Ogden J , Wright S D , et al. The influence of macro-itterfall and forest structure on litterfall damage to seedlings[J]. Austral Ecology, 2004, 29(3):305-312.
[16]Pierson E A , Mack R N . The population biology of Bromus tectorum in forests:effect of disturbance, grazing, and litter on seedling establishment and reproduction[J]. Oecologia, 1990, 84(4):526-533.
[17]刘尚华,石凤翎,吕世海,等.京西百花山区植物群落凋落物对土壤种子库的影响[J].水土保持通报,2008,28(2):41-47.
[18]周存宇.凋落物在森林生态系统中的作用及其研究进展[J].湖北农学院学报,2003,23(2):140-145.
[19]Manzoni S , Trofymow J A , Porporato J A . Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter[J]. Ecological Monographs, 2010, 80(1):89-106.
[20]陈辉,唐明,刘安全,等.人工竹林凋落物中真菌群落的研究[J].西北林学院学报,1993(4):41-45.
[21]May F E , Ash J E . An Assessment of the Allelopathic Potential of Eucalyptus[J]. Australian Journal of Botany, 1990, 38(3):245-254.
[22]Halvorson J J , Belesky D P , West M S . Inhibition of forage seed germination by leaf litter extracts of overstory hardwoods used in silvopastoral systems[J]. Agroforestry Systems, 2016, 91(1):1-15.
[23]罗侠.天山云杉自毒作用的生化机制研究[D].乌鲁木齐:新疆农业大学,2006.
[24]李兆慧.天山云杉自毒作用与自毒物质的鉴定[D].杭州:浙江大学,2011.
[25]Reigosa M J , Gonzalez L . Forest ecosystems and allelopathy[M]// Allelopathy. Springer Netherlands, 2006.
[26]Bu'Lock J D , Harbone J B . Phenolic compounds derived from shrikimate[J]. 1980.
[27]孔垂華.植物化感作用研究中应注意的问题[J].应用生态学报,1998,9(3):332-336.
[28]徐亮,包维楷,何永华.种子贮藏物质变化及其贮藏生理[J].种子,2003(5):60-63.
[29]杨期和,叶万辉,廖富林,等.植物化感物质对种子萌发的影响[J].生态学杂志,2005,24(12):1459-1465.
[30]陈慧玲,张新叶,杨彦伶.油茶水浸液对桔梗种子的化感效应[J].林业工程学报,2013,27(3):61-64.
[31]Jensen H A . Content of buried seeds in arable soil in Denmark and its relation to the weed population[J]. Dansk Bot Arkiv, 1969, 41(4):314-20.
[32]Rice E L . Allelopathy[J]. Encyclopedia of Entomology, 1984(1):292-293.
[33]Muller W H . Volatile Materials Produced by Salvia leucophylla:Effects on Seedling Growth and Soil Bacteria[J]. Botanical Gazette, 1965, 126(3):195-200.
[34]蒋有绪.川西亚高山冷杉林枯枝落叶层的群落学作用[J].植物生态学报,1981,5(2):4-13.
[35]李文华,赵景柱.生态学研究回顾与展望[M].北京:气象出版社,2004.
[36]盛海燕,常杰,殷现伟,等.濒危植物明党参种子散布和种子库动态研究[J].生物多样性,2002,10(3):269-273.
[37]Bewley,J.D.Seed Germination and Dormancy[J]. The Plant Cell Online, 1997, 9(7):1055-1066.
[38]Dechoum M S , Castellani T T , Zalba S M , et al. Community structure, succession and invasibility in a seasonal deciduous forest in southern Brazil.[J]. Biological Invasions, 2015, 17(6):1697-1712.
[39]Sevik, H., Cetin, M. Effects of water stress on seed germination for select landscape plants[J]. Pol. J. Environ. Stud, 2015,24(2), 689-693.
[40]Jimenezalfaro B , Silveira F O O , Fidelis A , et al. Seed germination traits can contribute better to plant community ecology[J]. Journal of Vegetation Science, 2016, 27(3):637-645.
[41]张敏,朱教君,闫巧玲.光对种子萌发的影响机理研究进展[J].植物生态学报,2012,36(8):899-908.
[42]颜安,蒋平安,盛建东,等.玛纳斯河流域表层土壤盐分空间变异特征研究[J].土壤学报,2014(2):410-414.
[43]Zobel M , Kalamees R , Kersti Püssa, et al. Soil seed bank and vegetation in mixed coniferous forest stands with different disturbance regimes[J]. Forest Ecology and Management, 2007, 250(1-2):0-76.
[44]Wagner M , Heinrich W , Jetschke G . Seed bank assembly in an unmanaged ruderal grassland recovering from long-term exposure to industrial emissions[J]. Acta Oecologica, 2006, 30(3):342-352.
[45]Leck M A,Graveline K J.The seed bank of a freshwater tibal marsh[J].American Journal of Botany,1979,66(9):1006-1015.
[46]Peco B , Ortega M , Levassor C . Similarity between seed bank and vegetation in Mediterranean grassland:a predictive model[J]. Journal of Vegetation Science, 1998, 9(6):815-828.
[47]Manzano P, Malo J E , et al. Sheep gut passage and survival of Mediterranean shrub seeds[J]. Seed Science Research, 2005, 15(1):21-28.
[48]Henderson C B,Petersen K E,Redak RA.Spatial and temporal patterns in the bank and vetgetation of a desert grassland community [J].Journal of Ecology,1988,76(3):717-728.
[49]Roberts H A . Studies on the Weeds of Vegetable Crops:II. Effect of Six Years of Cropping on the Weed Seeds in the Soil[J]. Journal of Ecology, 1962, 50(3):803-813.
[50]Roach D A . Buried seed and standing vegetation in two adjacent tundra habitats, northern Alaska[J]. Oecologia, 1983, 60(3):359-364.
[51]刘明洪.长白山地区不同林型土壤种子库组成特征及与地上植被的关系[D].长春:东北师范大学,2010.
[52]Oswald A,Ransom J K . Striga control and improved farm productivity using crop rotation[J]. Crop Protection, 2001, 20(2):113-120.
[53]Jensen H A . Content of buried seeds in arable soil in Denmark and its relation to the weed population[J]. Dansk Bot Arkiv, 1969, 41(4):314-20.
[54]Thompson. Small-Scale Heterogeneity in the Seed Bank of An Acidic Grassland[J]. Journal of Ecology, 1986, 74(3):733.
[55]Pickett S T A , Mcdonnell M J . CHAPTER 8–Seed Bank Dynamics in Temperate Deciduous Forest[J]. Ecology of Soil Seed Banks, 1989:123-147.
[56]尚占環,徐鹏彬,任国华,等.土壤种子库研究综述——植被系统中的作用及功能[J].草业学报,2009,18(2):175-183.
(责任编辑:夏剑萍)
