
动物的共生菌固氮
2019-07-05 10:59金明霞嵇保中刘曙雯徐立军熊佳新喻爱林谢谷艾
生态学报 2019年10期
金明霞,王 怡,嵇保中,*,刘曙雯,徐立军,张 磊,熊佳新,喻爱林,谢谷艾
1 南京林业大学南方现代林业协同创新中心;南京林业大学林学院,南京 210037 2 江西省林业科学院,南昌 330013 3 南京中山陵园管理局,南京 210014
1888年,Beijerinck首次分离出具有固氮活性的微生物,1893年Glatz分离出厌氧型固氮梭菌,这些工作开创了研究生物固氮和固氮微生物的科学领域[1]。1912年Peklo从蚜虫体内分离获得固氮菌[2],此后不同作者陆续开展了白蚁[3- 5]、蚜虫[6]、小蠹虫[2]、药材甲、谷盗、衣蛾、果蝇[7]等共生菌固氮能力的研究。由于缺乏高精度检测仪器,这些早期研究并未提供动物共生菌固氮的有力证据。这种局面直到乙炔还原法的出现才得以改变,乙炔还原法为生物固氮研究提供了简便准确的检测方法,首先在白蚁共生菌固氮检测中获得成功[8- 9]。此后,针对不同昆虫的类似工作陆续展开[10- 11]。不仅测定固氮酶活性,还对消化道内的共生固氮菌进行分离鉴定[12- 13]。动物共生固氮研究的另一重要进展是分子生物学技术的应用,从而克服了许多固氮菌不能分离培养的局限。钼铁蛋白和铁蛋白固氮酶系统中铁蛋白由nifH基因编码,nifH基因高度保守,对其检测可用于固氮菌类群鉴定和系统发育研究[14]。虽然国外对动物共生菌固氮的研究已逾百载,但国内的相关研究尚处于起步阶段。黄小芳等[1]、王亚召等[14]曾分别撰文介绍海洋动物和白蚁共生菌固氮的研究情况。本文从动物共生菌固氮涉及的动物和微生物类群、动物共生菌固氮的性质和机理等角度,介绍动物共生菌固氮的研究进展。
1 共生菌固氮涉及的动物类群
一般认为,动物共生菌固氮现象主要存在于贫氮营养型动物,如珊瑚、白蚁、船蛆、食木鲶鱼等。但已报道具有固氮酶活性或具有固氮酶基因的动物已远超上述范畴,涉及的动物包括海绵、珊瑚、蚯蚓、磷虾、马陆、昆虫、蜗牛、船蛆、松鸡、奶牛、山羊、猪、河狸、沙鼠、豚鼠、人类等。据本文统计,已报道共生菌固氮动物有7门13纲23目50科99属174种(见表1) 。

表1 已报道的共生菌固氮动物
▲:乙炔还原法;◆:固氮酶基因检测;★:15N同位素检测;●:固氮菌分离鉴定
从海绵到人类,涉及低等到高等多种动物类群。其中有贫氮营养型动物,也有杂食性动物。表1所列174种动物中,多孔动物门2种(1.15%),刺胞动物门17种(9.77%),线虫动物门1种(0.58%),环节动物门9种(5.17%),节肢动物门113种(64.94%),软体动物门11种(6.32%),脊索动物门21种(12.07%)。节肢动物种类最多,其次为脊索动物、刺胞动物、软体动物和环节动物,多孔动物和线虫动物仅有少量研究。节肢动物113种中昆虫纲109种,占96.5%。昆虫纲中等翅目75种,占昆虫纲种类的68.8%;鞘翅目18种,占16.5%;膜翅目13种,占11.9%。脊索动物21种中,辐鳍鱼纲1种,鸟纲2种,哺乳纲18种。刺胞动物17种全部为珊瑚纲种类,其中石珊瑚目16种,柳珊瑚目1种。
2 动物共生固氮菌的类群特点
2.1 常见种类
表2所列52种动物所涉及固氮菌87属137种,大多数种类属于细菌,也有少数古菌,如Methanobacteriumivanovii[94]和鲁米叶甲烷马赛里球菌[93]。水生动物中还发现有藻类[24,49]。按出现频次,表2所列固氮菌中常见种类有慢生根瘤菌Bradyrhizobiumsp.(8次)、肠杆菌Enterobactersp.(7次)、肺炎克雷伯氏菌(6次)、葡萄球菌Staphylococcussp.(5次)、肠杆菌Enterobacteriaceaesp.(5次)、链球菌Streptococcussp.(4次)、法氏柠檬酸杆菌(4次)。出现3次的有11种,分别为圆褐固氮菌、拟杆菌Bacteroidessp.、芽孢杆菌Bacillussp.、巴氏固氮梭菌、催娩克雷白氏菌、Klebsiellasp.、甲基孢囊菌Methylocystissp.、根瘤菌Rhizobiumsp.、红球菌Rhodococcussp.、中华根瘤菌Sinorhizobiumsp.、密螺旋体Treponemasp.。出现2次的有23种。出现1次的96种,约占全部种类的70%。多数种类仅出现1次,表明研究的动物种类仍然较少,发现的固氮菌种类较为分散,实际蕴含的种类资源可能更为丰富(表2中固氮菌生态类型划分为:A:植物共生固氮菌;B:植物内生固氮菌;C:植物根际固氮菌;D:自生固氮菌;AB:植物共生/植物内生固氮菌;AD:植物共生/自生固氮菌;BC:植物内生/植物根际固氮菌;BD:植物内生/自生固氮菌)[102-116]。
表2 动物共生固氮菌种类与生态类型
Table 2 The Species and ecological types of animal symbiotic nitrogen xation bacteria

表2 动物共生固氮菌种类与生态类型
宿主/固氮菌种类/文献来源Host/Types of symbiotic nitrogen-fixing bacteria/Literature sources宿主/固氮菌种类/文献来源Host/Types of symbiotic nitrogen-fixing bacteria/Literature sources多孔动物门Porifera、寻常海绵纲 DemospongiaeNasutitermes graveolus/Enterobacter sp., Staphylococcus sp.[13]Ircinia strobilina, Mycale laxissima/DAzotobacter chroococum, DDesulfovibrio salexigens, Leptolyngbya sp., Methylocystis sp., Myxosarcina sp., Tolypothrix sp., Trichodesmium thiebautii[18]Nasutitermes nigriceps/Ensifer sp.[70]刺胞动物门Cnidaria、珊瑚纲AnthozoaNeotermes castaneus/Arthrobacter sp., Bacteroides. Enterobacteriaceae, Myxobacterales, CStaphylococcus sp.[90]Acropora millepora/Alphaproteobacteria sp., Bradyrhizobium sp.,Cyanobacteria sp., Deltaproteobacteria sp., Desulfuromonas sp.,Gammaproteobacteria sp., Myxosarcina sp., Sinorhizobium sp., Xenococcus sp.[23]Macrotermes sp./Azotobacter sp., Beijerinckia sp., DClostridium pasteur-ianumm, Klebsiella sp.[60]A. robusta/Bradyrhizobium sp., Paenibacillus durus, Rhizobium sp., Sinorhizobium sp.[27]Odontotermes formosanus/BDKlebsiella pneumoniae[95]; BKlebsiella variicola[80]Pocillopora damicornis, Acropora millepora, A. muricata/Bradyrhizobium sp., Cylindrospermopsisraciborskii, DDesulfovibrio salexigens, Dickeya dadantii, Methylobacter luteus, Methylocystis echinoides, Nodularia spumigena, Trichodesmium erythraeum, Wolinella succinogenes[25]Odontotaenius disjunctus/Paludibacter propionicigenes[88]Montipora capitata, M. flabellata/BAcetobacter diazotrophicus,DAzomonas agilis, BDA. macrocytogenes, BCAzospirillum lipoferum, DAzotobacter chroococcum, DA. vineladii, Bradyrhizobium sp.,Desulfobacter latus, Halorhodospira halophila, Marichromatium purpura-tum, AMesorhizobium loti[35]Conorhynchus pistor/BDKlebsiella pneumonia[17]Porites lutea/Acetobacterium woodii, ABradyrhizobium japonicum, ABBurkholderia cepacia, Chroococcidiopsis sp., CClostridium themocel-lum, Ideonella sp., Leptolyngbya minuta, L. nodulosa, Marichromatum purpuratum, CPseudomonas stutzeri[49]Dendroctonus frontalis, D. terebrans, Ips avulsus/Enterobacter aerogenes, BE. agglomerans, E. sp.[20]Pocillopora damicornis/BCAzospirillum brasilensis, ABradyrhizobium denitrificans, B. sp., Chlorobium limicola, C. phaeobacteroides,BKlebsiella pneumoniae, Klebsiella sp., ARhizobium rosettiformans, Sinorhizobium sp., Teredinibacter turnerae[49]; DAzotobacter chroococcum, BCAzospirillum halopraeferens, ABradyrhizobium denitrificans, Klebsiella sp., CPseudomonas stutzeri, Rhizobium sp.[96]Dendroctonus valens, D. rhizophagus /DRaoultella terrigena[21]环节动物门Annelida、寡毛纲 ClitellataBactrocera tryoni/BCKlebsiella oxytoca[31]Pheretima vittata/BCKlebsiella oxytoca[63]Ceratitis capitata/DCitrobacter freundii, Enterobacter sp., Erwinia amylo-vara, BCKlebsiella planticola, BDK. pneumoniae, BCK. oxytoca, Pectobacte-rium carotovora, P. cypripedii, BPantoea agglomerans[34]节肢动物门Arthropoda软甲纲Malacostraca、磷虾目Euphausiacea;甲壳纲Crustacea、桡足亚纲 CopepodaAcromyrmex octospinosus/Rhizobiales[39]Euphausia sp., Copepods/Chlorobium tepidum, DChromatium purpuratum, Desulfobacter curvatus, Desulfonema limicola[66]Tetraponera binghami/Burkholderia sp., Flavobacterium sp., Rhizobium sp., Methylobacterium sp., Pseudomonas sp.[51]昆虫纲Insecta软体动物门Mollusca、双壳纲BivalviaCryptocercus punctulatus/Desulfovibrio sp.[75]Lyrodus pedicellatus/Teredinibacter turnerae[97]Mastotermes darwiniensis/DCitrobacter freundii[12]; Enterobacter sp., Flavobacterium sp., Streptococcus sp.[13]; Desulfovibrio sp.[22]脊索动物门Chordata、辐鳍鱼纲ActinopterygiiZootermopsis angusticollis/Treponema sp.[98]Panaque nigrolineatus/BDAzomonas macrocytogenes, AAzorhizobium doebere-inerae, ABradyrhizobium japonicum, B. sp., Cellulosilyticum lentocellum, CClostridium arbusti, C. beijerinckii, CC. papyrosolvens, CC. tyrobutyricum, Desulfomonile tiedjei,Methylocystis parvus, M. sp., Opitutaceae, ASinorhizobium meliloti, CSphingomonas azotifigens[67]Cryptotermes primus/Enterobacter sp., Streptococcus sp.[13]脊索动物门Chordata、哺乳纲MammaliaCoptotermes formosanus/BEnterobacter agglomerans[99]; Acinetobacter sp., BDKlebsiella pneumonia[100]Bos tauru/Acinetobacter baumannii, BCBacillus methylotrophicus, BB. sonorensis, CStaphylococcus caprae[72]Coptotermes lacteus/DCitrobacter freundii[12];Enterobacter sp., Strepto-coccus sp.[13]Susscrofa domestica, Cavia porcellus/BCEnterobacter cloacae, Klebsiella aero-genes[79]

表2 动物共生固氮菌种类与生态类型
续表宿主/固氮菌种类/文献来源Host/Types of symbiotic nitrogen-fixing bacteria/Literature sources宿主/固氮菌种类/文献来源Host/Types of symbiotic nitrogen-fixing bacteria/Literature sourcesHeterotermes ferox/Enterobacter sp., Streptococcus sp.[13]Ochotona hyperborea/Bradyrhizobium sp., Bryantella formatexigens, Clos-tridium cellulovorans, DC. pasteurianum, Fibrobacter succinogenes subsp. succinogenes, Sphingomonas sp., Treponema sp.[101]H. indicola/Desulfovibrio termitidis[22]Ochotona dauurica, Ochotona hyperborea, Ochotona mantchurica/Bradyrhi-zobium sp., Bryantella formatexigens, Clostridium cellulovorans, Fibrobacter succinogenes, Sphingomonas sp., Treponema sp.[89]Reticulitermes lucifugus/Bacillus sp., Enterobacteriaceae, Erwinia sp., Rhodococcus sp., CStaphylococcussimulans, CS. haemolyticus[33]Meriones meridianus, M. tamariscinus/Bacillus sp., Corynebacterium sp., Enterobacteriaceae sp.[82]Reticulitermes flavipes/Desulfovibrio desulfuricans[22]Microtus rossiaemeridionalis/Arthrobacter sp., Bacillus cereus, BB. licheniformis, BCB. subtilis, Bacteroides sp., Curtobacterium sp., Mi-crococcus sp., Myxococcales, Rhodococcus sp., Staphylococcus sp. [84]Reticulitermes speratus/DClostridium pasteurianum, Desulfovibrio gigas, Methanobacterium ivanovii[94]Clethrionomys glareolus/BBacillus licheniformis, BCB. subtilis, Bacteroides sp., Enterobacteriaceae, Micrococcus sp., Myxococcales, Nocardia sp.[84]Nasutitermes exitiosus/DCitrobacter freundii, Staphylococcus sp.[13]Homo sapiens/BCEnterobacter cloacae, Klebsiella aerogenes[79]; Coprococcus catus, C. comes, Desulfitobacterium hafniense, Eubacterium hallii, E. sp., BDKlebsiella pneumoniae, Lachnospira multipara, Methylocystis sp., Megas-phaera elsdenii, Methanomassiliicoccus luminyensis, Ruminococcus obeum, Pyramidobactor piscolens, Roseburia sp.[93]
2.2 动物共生固氮菌的生态类型多样性
表2所列动物共生固氮菌种类,涵盖植物固氮菌和自生固氮菌等多种生态类型。其中40种曾作为植物共生固氮菌、植物内生固氮菌、植物根际固氮菌、自生固氮菌予以报道。植物共生固氮菌6种:德式固氮根瘤菌、Bradyrhizobiumdenitrificans、慢生大豆根瘤菌、百脉根中生根瘤菌、Rhizobiumrosettiformans、苜蓿中华根瘤菌。植物内生固氮菌7种:固氮醋杆菌、巨胞氮单胞菌、地衣芽孢杆菌、索诺拉沙漠芽孢杆菌、聚团肠杆菌、变栖克雷伯氏菌、成团泛菌。植物根际固氮菌8种:热纤梭菌、溶纸莎草梭菌、酪丁酸梭菌、斯氏假单胞菌、Sphingomonasazotifigens、山羊葡萄球菌、溶血性葡萄球菌、模仿葡萄球菌。自生固氮菌8种:敏捷氮单胞菌、圆褐固氮菌、棕色固氮菌、着色菌、弗氏柠檬酸杆菌、巴氏固氮梭菌、需盐脱硫弧菌、土生拉乌尔菌。植物共生/内生固氮菌1种,即洋葱伯克霍尔德菌。植物内生/根际固氮菌8种:巴西固氮螺菌、高盐固氮螺菌、生脂固氮螺菌、甲基营养芽孢杆菌、枯草芽胞杆菌、阴沟肠杆菌、 植生克雷白氏菌、催娩克雷伯氏菌。植物内生/自生固氮菌2种:巨胞氮单胞菌和肺炎克雷伯氏菌。
2.3 固氮菌的生态类型转换
在固氮菌中较为常见,兼性内生固氮菌,如一些固氮螺菌属细菌,既能在根内也能在根表和土壤中定殖[117]。表2中洋葱伯克霍尔德菌兼具植物共生固氮[102]和植物内生固氮能力[118]。巨胞氮单胞菌既是胡杨的内生固氮菌[114],又是自生固氮菌[115]。Frohlich等[70]用象白蚁Nasutitermesnigriceps肠道分离获得的剑菌培养物,对12种豆科植物进行接种试验,在豌豆、印度田菁、葫芦巴上均形成了根瘤,证实动物肠道固氮菌和植物共生固氮菌之间存在生态类型的转化。宾氏细长蚁中、后肠结合部有菌囊,菌囊外密布气管和马氏管。囊内含有伯克霍尔德菌、黄质菌、根瘤菌、甲基杆菌、假单胞菌[51]。伯克霍尔德菌属于植物共生和自生固氮菌,黄质菌可营植物内生固氮,根瘤菌则属于植物共生/植物内生和自生固氮菌,甲基杆菌是植物共生固氮菌常见类群,假单胞菌在植物共生/植物内生/植物根际固氮菌中常有发现。上述结果表明蚂蚁体内菌囊的内部环境似乎比固氮植物的根瘤更复杂,能包容许多固氮菌种类。一些不能形成植物根瘤的种类,如黄质菌,在菌囊内也能正常发育。
3 动物共生固氮菌的来源

但用荧光标记的聚团肠杆菌和肺炎克雷伯氏菌饲喂地中海果蝇成虫,在成虫消化道和卵表以及幼虫和蛹的消化道均检出上述细菌的存在[124]。说明肠道共生固氮菌也存在成虫-卵之间的垂直传递。Ayayee等在经过表面灭菌处理的光肩星天牛卵内检测到脲酶、固氮酶基因及其转录活性,表明脲分解细菌和固氮菌存在通过遗传进行垂直传递的可能性[28]。
4 动物共生菌固氮的性质
一般认为动物肠道共生菌固氮属于联合固氮,柯为称白蚁肠道内固氮菌是一种“联合共生固氮形式”在动物界的出现[125]。祁永青等则将联合固氮菌定义为“必须生活在植物根际、叶面或动物肠道等处才能固氮的原核生物”[126]。另外有珊瑚和固氮细菌的互作联合固氮,如石珊瑚和固氮菌属的联合固氮,海洋蓝藻与海绵属形成内共生关系,蛤类与蓝藻、固氮菌的联合固氮[127]。此外,动物共生固氮菌固氮活性容易受环境影响,也具有较明显的联合固氮特征。白蚁种类、品级以及不同季节等均影响固氮活性,其中食物含氮量为主要影响因子,其他因子在不同程度上也可溯及食物含氮量的关系[14]。食性对固氮活性的影响一般表现为低等食木白蚁固氮活性强,高等培菌白蚁和食土白蚁固氮活性弱或没有活性[14]。在食料中添加(NH4)2SO4或KNO3均能显著减低台湾乳白蚁工蚁肠道共生固氮菌的固氮能力[9]。已经分离获得的固氮菌中,许多种类属于兼性厌氧菌、生境较为广泛,似乎与白蚁构成专性共生的可能性不大[14]。Vecherskii等对欧洲散白蚁共生菌固氮研究表明:活体和蚁巢物质在好氧和厌氧条件下均有固氮酶活性,白蚁肠道内细菌群落与蚁巢物质的类似[33]。因此认为该种白蚁与共生固氮菌之间关系属于联合固氮作用。但也有研究表明动物肠道有特殊的含菌构造,如宾氏细长蚁中、后肠结合部有类似于根瘤的菌囊,囊内含有多种固氮菌[51]。此外,固氮酶基因检测表明,动物肠道固氮菌区系十分丰富,能够分离培养的仅为其极少部分,不能排除专性共生体系的存在。
5 动物共生菌固氮的微生境与共生复合体
5.1 动物共生菌固氮的微生境
氧气、氢气、pH和氧化还原电位等是肠道微生境的主要指标,在肠道不同部位这些指标存在明显差异,形成多样化的微生境,供不同生理需求的固氮菌生长发育。低等白蚁大多数肠道微生物位于后肠前段膨大的后肠囊内。高等白蚁肠道区室化有逐渐复杂化的趋势,食土白蚁后肠膨大形成连续的5个囊室,相邻囊室之间以肌肉阀瓣分隔。消化道不同部位以及不同的后肠囊内氧气、氢气、pH和氧化还原电位均有差异[128]。黄肢散白蚁和象白蚁的后肠囊氧气浓度分布呈现两个不同区域,外侧为微氧层,约占后肠囊体积的60%,其中生活的共生菌消耗掉向肠道内扩散的氧气,形成氧阱,造成后肠囊中心部分(40%)严格的厌氧环境。与此相反,后肠囊中心部位氢气浓度最高,并通过两个区域消耗氢气,抑制其向外扩散。一个区域是临近后肠囊中心部位的肠腔区域,另一个是后肠壁区域。氧气和氢气的上述分布特点使厌氧和微好氧微生物各得其所,中心部分严格的厌氧环境是发酵生成乙酸酯的场所,而乙酸酯是宿主昆虫的主要能源物质[129]。具角黑艳甲成虫的消化道分为前肠、中肠、前后肠和后后肠4段,每段中心部位均有一个厌氧核心区,由此向外氧气含量逐渐增高。放线菌和根瘤菌主要分布于前肠和后后肠。前后肠内表面肠壁细胞形态分化最大,形成密集的胞窝和棘刺,将空间进一步区室化供细菌、原生动物、线虫等定殖,其中拟杆菌和梭菌为主要类群。该部位还是产甲烷古菌的唯一分布区域和共生菌固氮核心部位[130]。
5.2 共生复合体
固氮菌一般与多种生物形成复合体,以满足固氮过程的能源和固氮产物同化等方面的需求。台湾乳白蚁与后肠的伪披发虫Pseudotrichonymphagrassii、伪披发虫内生菌Azobacteroidespseudotrichonymphae形成三重共生关系。食物通过口器和前肠粉碎、中肠消化吸收后,形成较为纯粹的木质纤维素微粒,这些微粒成为伪披发虫的食料,经伪披发虫分解后形成能量物质,为白蚁和固氮菌利用。A.pseudotrichonymphae则主要负责固氮和氮素营养的转化,为披发虫和白蚁提供氮素营养[16]。隐尾蠊后肠内原生动物披发虫Barbulanympha体表有许多外共生的拟杆菌,这些外共生菌具有固氮能力,与隐尾蠊、披发虫构成三重共生复合体[75]。猪毛菜属的Salsolainermis为旱生植物,与锥喙象以及锥喙象肠道固氮菌肺炎克雷伯氏菌形成三重共生。象甲形成紧密附着在植物根部的土室,以此获得碳素、水分等营养并躲避敌害,肺炎克雷伯氏菌通过共生固氮提供氮素营养,并促进植物生长[17]。固氮菌一般与纤维素分解菌协同作用,以满足各自对能量和氮素营养的需求。将黑翅土白蚁肠道分离的纤维素分解菌与固氮菌混合培养,纤维素酶活性与固氮酶活性均有显著提高[131]。从奶牛瘤胃中分离的纤维素分解菌和固氮菌进行混合体外发酵,混合菌株的添加对瘤胃的发酵也具有促进作用[132]。纤维素分解和固氮也可在同一种细菌体内完成。船蛆鳃上着生有Deshayes腺,腺体细胞内的船蛆杆菌既可分解木质纤维素,也可固氮[59,61]。
6 动物共生菌固氮产物的同化
6.1 动物体内氨同化途径
氨既是共生固氮的产物,对组织细胞也具有毒害作用,必须转化为有机氮素营养或及时清除。氨转化为含氮有机物主要有4种途径:(1)谷氨酰胺合成酶(GS)/谷氨酸合成酶(GOGAT)途径:氨与谷氨酸(Glu)在GS催化下生成谷氨酰胺(Gln),Gln在GOGAT催化下将氨基转移给α-酮戊二酸,生成2分子Glu。GS有3类:GSⅠ主要存在于细菌;GSⅡ主要存在于植物共生细菌以及真核细胞中;GSⅢ发现于专性厌氧杆菌、一些厌氧细菌和蓝细菌。GS/GOGAT对氨的亲和性高,主要在低氨浓度时发挥作用。(2)谷氨酸脱氢酶(GDH)途径:氨与α-酮戊二酸在GDH催化下生成Glu。GDH有两种,NAD-GDH降解谷氨酸,NADP-GDH用于谷氨酸的合成。GDH在高氨浓度时占主导作用。(3)丙氨酸脱氢酶(ADH)途径:ADH催化丙酮酸和铵生成丙氨酸;(4)天冬氨酸酶途径:催化延胡索酸和铵生成天冬氨酸[133]。
6.2 反刍动物瘤胃氨的同化
主要由细菌和真菌完成,瘤胃细菌可分为发酵结构性碳水化合物和发酵非结构性碳水化合物两类,前者仅能利用氨作为氮源,后者则可利用氨、氨基酸和肽作为氮源。结构性碳水化合物主要包括纤维素、半纤维素和木质素。瘤胃纤维分解菌对氨氮的利用主要是通过GDH和ADH途径。非结构性碳水化合物包括水溶性碳水化合物,如单糖、双糖、低聚糖和一些多糖,以及不溶于水的大分子多糖。非纤维分解菌对氨氮的利用主要是通过GDH路径和GS路径,可以从头合成多种自身所需氨基酸,且大多数不存在GOGAT路径。瘤胃厌氧真菌缺乏蛋白质降解能力,可以以氨为唯一氮源合成自身所需氨基酸满足生长需求。GDH和GS-GOGAT途径是瘤胃真菌同化氨的2条主要路径[133]。
6.3 珊瑚体内氨的同化
珊瑚对氨的同化由肠壁细胞与虫黄藻协同完成,虫黄藻位于珊瑚肠壁细胞细胞质内,能通过光合作用为珊瑚提供能量,还能通过GS/GOGAT途径将氨转化为氨基酸(见图1)。环境中的氨经珊瑚肠腔进入肠壁细胞质,再进入虫黄藻细胞质,通过GS/GOGAT途径转化为Gln和Glu。所形成的Gln、Glu可以直接转移进入珊瑚肠壁细胞质,进而转化为其他氨基酸,合成蛋白质。蛋白质代谢形成的氨又可扩散进入肠腔,再次吸收利用或排出体外。肠壁细胞质也存在GS/GOGAT途径和GDH途径,但转化速率明显低于虫黄藻共生体[135]。
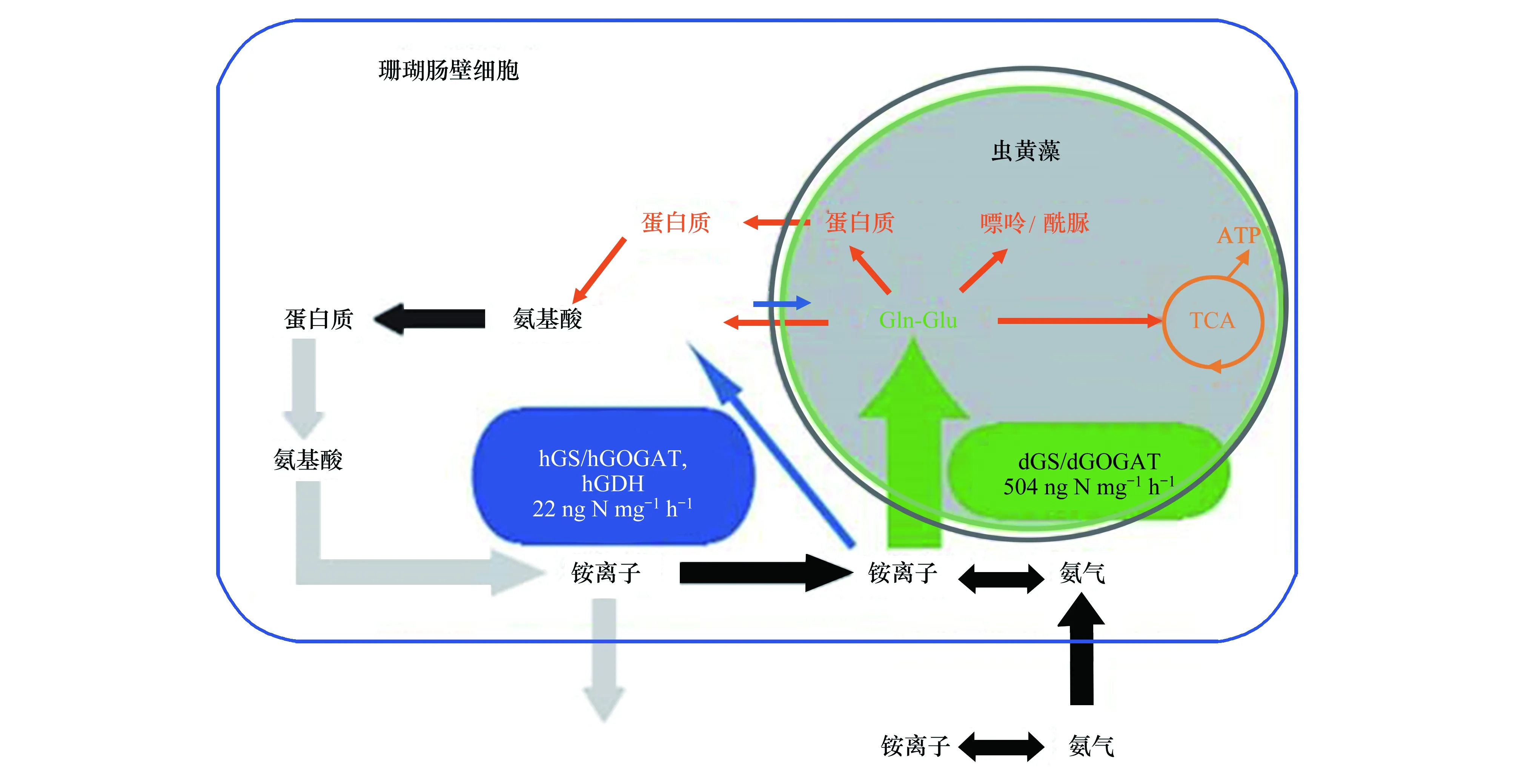
图1 珊瑚-虫黄藻共生体氨同化吸收模型[135]Fig.1 Model of ammonium assimilation in the coral-dinoflagellate symbiosis
6.4 白蚁体内纤维素分解、生物固氮与氨同化
6.4.1台湾乳白蚁-伪披发虫-内生菌复合体
台湾乳白蚁后肠有原生动物伪披发虫Pseudotrichonymphagrassii,伪披发虫有内生菌Azobacteroidespseudotrichonymphae。后肠内的木质纤维素颗粒被伪披发虫吞噬,降解为葡萄糖、木糖、糖醛酸等,进一步发酵形成CO2和H2,降解产物从批发虫细胞进入肠腔,在产酸菌作用下形成乙酸酯,成为能量物质。内生菌则发挥固氮和氮转化功能。披发虫细胞本身缺乏脲酶和GS,固氮形成的氨必须在内生菌体内GS催化下生成Gln,进而转化为其他氨基酸和辅因子。菌体内形成的氨基酸进入披发虫细胞质内进行含氮有机物转化,所形成的脲和氨再进入菌体内进行Gln和氨基酸合成(图2)[16]

图2 台湾乳白蚁-伪披发虫-内生菌复合体共生作用模型[134]Fig.2 Model of symbiosis among Coptotermes formosanus, Pseudotrichonympha grassii and its endosymbiotic bacteria
6.4.2栖北散白蚁-固氮菌-披发虫-白蚁菌群-1(TG1)复合体
栖北散白蚁后肠内有原生动物披发虫Trichonymphaagilis,披发虫内生菌为白蚁菌群1(TG1),后肠肠腔内还有共生固氮菌。木质纤维素颗粒在披发虫细胞质内分解形成葡萄糖- 6-磷酸等碳水化合物,并形成CO2、H2、乙酸酯等代谢产物。代谢产物进入肠腔,在产酸菌作用下形成乙酸酯。肠腔内固氮菌形成的氨,可以被披发虫吸收,经过GS途径合成Gln后进入TG1菌体内,转化为氨基酸和辅因子(见图3)。后肠肠腔内的营养物、披发虫、细菌组成的营养液,可能通过肛交哺和粪食性,被蚁巢内不同个体分享[136]。
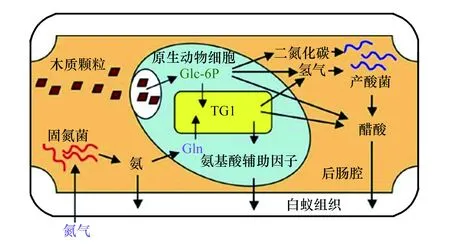
图3 栖北散白蚁-固氮菌-披发虫-白蚁菌群- 1(TG1)复合体共生作用模型[136]Fig.3 Model of symbiosis among Reticulitermes speratus, symbiotic nitrogen-fixing bacteria, Trichonympha agilis and TG1
6.4.3白蚁共生菌固氮产物的吸收路径
Bentley用15N2作为氮源培养Nusutitermescorniger工蚁和兵蚁,24h后分别检测肠道、头部、虫体其他部分固氮产物含量,发现三类样品中均有固氮产物检出,证实了共生菌固氮与产物同化现象[64]。不同部位固氮产物含量:工蚁为肠道>虫体其他部分>头部,兵蚁为肠道>头部>虫体其他部分。肠道为固氮产物的最初形成场所,固氮产物可以通过交哺在工蚁和兵蚁个体间转移,从工蚁向兵蚁的转移为主流方向。Fujita 等对栖北散白蚁的研究表明:溶菌酶活性主要分布于唾腺,前肠的溶菌酶来源于唾腺[137]。蛋白酶活性和游离氨基酸含量中肠最高。推测栖北散白蚁通过唾腺分泌的溶菌酶分解肛交哺获得的含菌后肠液,而溶菌酶进入中肠后被中肠蛋白酶分解失活,在后肠中活性很低,对后肠共生菌群较安全。此后,Fujita 等采用15N2示踪对栖北散白蚁生物固氮产物同化途径进行研究,固氮产物主要出现在后肠,其次为嗉囊与中肠,在头部以及虫体其他部分也有检出[36]。后肠、嗉囊与中肠内固氮产物首先出现,一段时间后,头部和虫体其他部分也出现固氮产物的踪迹。据此认为,大部分固氮产物的同化吸收并不是在后肠完成的,而是通过肛交哺转移到前肠,在前肠和中肠完成酶解和吸收过程。
7 共生菌固氮在动物氮素营养中的地位
动物氮素营养可以来自于食物、非蛋白氮以及共生菌固氮,其中非蛋白氮指氮素营养代谢形成的产物,如尿素及其衍生物、氨及铵盐、酰胺化合物等,非蛋白氮动物不能直接利用,但可作为共生菌的氮源,经共生菌转化后再次为动物利用。反刍动物的尿素在瘤胃中被细菌脲酶迅速水解成氨,部分氨参与瘤胃内微生物蛋白的合成,最终成为动物的氨基酸营养。另部分氨经由瘤胃扩散进入静脉,随血液在肝脏处经鸟氨酸循环再次合成尿素。肝脏合成的尿素被称作内源尿素,部分内源尿素可经由唾液或者血液再次返回到瘤胃内,被瘤胃细菌重新利用[139-140]。
鳞翅目、双翅目、脉翅目等全变态昆虫,在老熟幼虫、预蛹或预成虫阶段,则通过临时贮存非蛋白氮,将其转化为氨,合成新的氨基酸和蛋白质以满足结茧等特殊行为的需求[140-141]。栖北散白蚁将尿酸贮存在脂肪体尿盐细胞内,需要动用时,尿盐释入血淋巴,被马氏管摄取后排泄进入后肠腔,在后肠共生细菌作用下分解为氨,通过氨的同化进入氮素营养循环[142]。
氨不仅对生物固氮具有氨阻遏效应,对细胞还具有毒性,动物体内氮素代谢涉及营养和解毒等许多方面。通过食物氮、非蛋白氮以及生物固氮相互协调,形成营养和解毒的代谢网络,共同维持动物体内氮素营养的动态平衡。当食物中蛋白质营养丰富,非蛋白氮代谢产物含量高,动物体以解毒排泄为主,非蛋白氮的再利用以及生物固氮处于抑制状态;贫氮营养型动物,则通过非蛋白氮的循环利用和生物固氮,补充食物中氮素营养的不足。这些贫氮营养型动物,如白蚁、珊瑚、食草动物等,一般具有较高的共生菌固氮活性。生物固氮需要消耗很多能量,大量能源物质应用于氮素营养的合成,势必影响动物体内的能量权衡。因此,即使是具有共生菌固氮能力的白蚁,也依然对富氮营养食物表现出较强的趋性。取食含氮量高的食物时,体内共生固氮量下降[65]。
8 结语
一般认为,动物的共生菌固氮主要存在于贫氮营养型动物,但现有报道涉及的动物达到7门13纲23目50科99属174种,已远超上述范围。从海绵到人类,涉及低等到高等多种动物类群。虽然其中有些种类仅为固氮酶基因检出的结果,这些基因表达情况还有待进一步确认,但共生菌固氮可能是动物中普遍存在的现象。所涉及的固氮菌类群也十分丰富,其中大多数种类为细菌,也有少数古菌和藻类。已经发现的动物共生固氮菌种类与植物固氮菌具有明显的交叉性,植物共生固氮菌、植物内生固氮菌、植物根际固氮菌和自生固氮菌在不同动物肠道内均有发现。这些情况似乎表明生物固氮可能具有统一的固氮菌资源库,同一种固氮菌与植物共生、与动物共生或者自由生活,只是固氮菌对生境的选择,取决于环境条件是否能满足特定固氮菌固氮活性表达的需要。如电子和质子供体、能量供给、固氮酶活性保护以及解除氨阻遏等。这也表明动物肠道具有丰富的微生境,能满足不同固氮菌的生理需求。动物共生固氮菌将会成为生物固氮资源开发的富矿,在生物固氮研究、发酵工业、动物营养等领域展示其重要价值。
分子生物学技术的应用,对动物共生固氮研究发挥了很大的促进作用。但所发现的固氮菌资源以及固氮机理的理论模型,与具体情况的解释验证还有相当距离。动物共生菌固氮过程一般由多种动物和细菌组成的复合体,分别完成电子和质子提供、能量保障、氧阱形成以及氨的同化等任务。目前对这些过程基本上还不了解。动物共生固氮、植物共生固氮、自生固氮在固氮产物氨的同化方面均采用GS/GOGAT、GDH和ADH等途径,但不同生物体内相关途径的酶系来源存在较大差异。动物体内氨同化酶系是否完全源于共生菌?酶系来源差异影响动物营养代谢的机制如何等等,也是动物共生固氮机理研究的基本问题。此外,动物共生菌固氮的性质也需要研究确认。总体上,动物通过食物摄取、非蛋白氮排泄或循环利用以及共生菌固氮等途径,维持体内氮素营养的平衡和氮素代谢末端产物的降解排泄,随着研究的深入,将进一步加深人们对氮素资源利用和动物营养生理等方面的理解。
猜你喜欢
生物灾害科学(2022年2期)2022-10-15
农业科技与信息(2021年16期)2021-12-07
生态科学(2021年5期)2021-11-13
文萃报·周二版(2018年22期)2018-09-18
中学生数理化·高一版(2018年6期)2018-07-09
中国农资(2018年4期)2018-02-10
故事作文·高年级(2017年11期)2017-11-15
科学启蒙(2015年8期)2015-08-07
江西农业大学学报(2013年3期)2013-11-05
植物营养与肥料学报(2011年5期)2011-11-06
