
高温胁迫对洋竹草生长及部分生理指标的影响
2019-06-26 12:24张秀梅许建新何新杰张朝铖沈彦会张竞元
江西农业学报 2019年6期
张秀梅,许建新,何新杰,张朝铖,沈彦会,张竞元
(深圳市铁汉生态环境股份有限公司,广东 深圳 518000)
洋竹草(CallisiarepensJacq. L.)是鸭跖草科、锦竹草属多年生草本植物,原产于美洲,适用于韶关以南大部分区域,植株低矮,茎肉质,呈匍匐生长,节处生根;单叶互生,基部鞘状,长卵形或卵形,薄肉质,表面具蜡质光泽,翠绿色,时有紫色斑点;茎叶颜色随土壤水分和季节变化而变色,春夏叶片为翠绿色,秋冬季或茎叶老时变为酒红色,性喜温暖、湿润的环境条件,对土壤要求不严,抗逆性强,抗病虫害;株型紧凑,覆盖度高,绿期长,冬季4 ℃仍可存活,常应用于屋顶绿化、垂直绿化、地被等,具有较大的应用价值与应用前景。目前国内外对洋竹草的研究报道较少;曾宪锋等[1]对洋竹草属中国大陆新归化属植物进行了研究,介绍了洋竹草在中国大陆的分布信息;曾红等[2]对洋竹草的抗旱能力与其他3种屋顶绿化植物做了对比研究,研究认为4种植物的抗旱性排序为洋竹草>玉吊钟>藓状景天>锦绣苋。本试验主要通过设置不同的高温胁迫,研究了洋竹草对不同高温胁迫的适应性,为洋竹草的栽培提供理论依据。
1 试验材料与方法
1.1 试验材料
洋竹草为多年生草本,鸭跖草科洋竹草属植物,与铺地锦竹同属。
1.2 试验方法
试验从2018年7月12日开始,地点在东莞市铁汉生态环境股份有限公司实验室,从苗圃选择生长稳定、长势相对一致、健壮无病虫害的幼苗,定植于内口径15 cm、高8.5 cm的塑料盆中,盆栽基质为泥炭土∶蛭石=1∶1(m/m),盆底铺两层细密纱布以防止浇水时基质外渗。试验于人工气候箱中进行,采用22 ℃(昼/夜)为对照,30 ℃(昼/夜)为轻度高温胁迫、38 ℃(昼/夜)为重度高温胁迫3个温度梯度处理,人工培养箱空气相对湿度75%,光照时间12 h/d,光照度为1200 lx。盆底设托盘(水深2 cm)补充基质水分,以防止高温引发的水分胁迫伤害;每处理设3次重复,每重复10株,分别持续7、10、15 d,观测洋竹草在不同温度设置下的表现。各高温处理完成后,于09:00~10:30分别取样测定植株叶片相对电导率,并取样待测叶片酶活性。以洋竹草置于适宜气候条件下正常生长作为对照组,分别于处理完成后同时测定、取样待测各项指标。
1.3 测定指标及方法
1.3.1 叶宽、叶长、叶厚 在每个处理完成后,随机选取10片洋竹草叶子并做好标记,用电子游标卡尺测定叶长、叶宽与叶厚;为了保证实验精准性,叶长、叶宽均选取最长、最宽处,做好标记,确保每次都是固定叶片。
1.3.2 生理指标测定 各指标测定方法:丙二醛含量采用巴比妥酸(TBA)显色法[3],SOD活性采用氮蓝四氮(NBT)法[3],POD活性测定参考愈创木酚法[3],CAT活性的测定参考紫外吸收法[3],相对电导率采用浸泡法[4]。
1.4 数据处理
采用Excel 2003对所有数据进行初步的整理,用SPSS 19.0进行one-way ANOVA单因素方差和显著性分析,绘图在origin 9.0软件中进行。
2 结果与分析
2.1 高温胁迫对洋竹草叶宽的影响
由图1可知,同一天不同温度处理下,洋竹草的叶宽随着温度的增加而逐渐减小;同一温度不同天数处理下,以22 ℃温度(CK)胁迫处理15 d的洋竹草叶宽最宽,在高温38 ℃胁迫15 d时洋竹草的叶宽最小,说明在对照适宜温度处理下,洋竹草叶片随着时间的增长正常生长,温度的升高使洋竹草叶片宽度变窄。由显著性差异分析得出,与对照相比,30 ℃处理下洋竹草叶宽在胁迫10 d时开始差异显著(P<0.05),叶宽与对照相比减幅在第10、15 天分别为16.354%、27.78%;38 ℃处理下洋竹草叶宽均与对照差异显著(P<0.05),叶宽与对照相比减幅在第10、15天分别为21.50%、31.85%,说明38 ℃干旱15 d时对洋竹草叶宽影响较大。
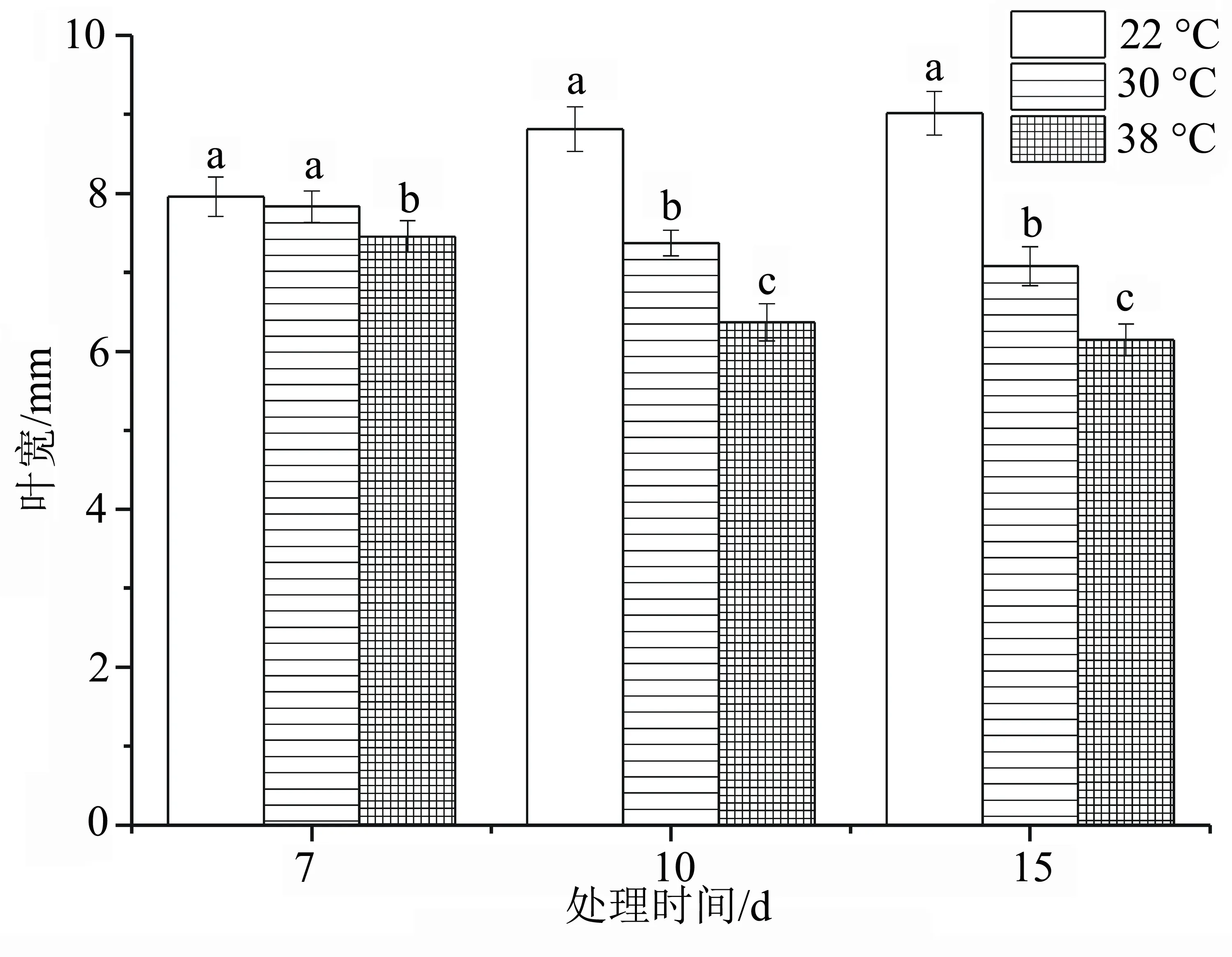
图1 高温胁迫下洋竹草叶宽的变化
2.2 高温胁迫对洋竹草叶长的影响
由图2可知,洋竹草叶长在10.64~16.32 mm。同一天数不同温度处理下,叶长随着温度的增加而逐渐下降,温度为38 ℃干旱15 d时叶长最短,为10.64 mm,相比对照减少了5.68 mm。在高温胁迫处理7 d时,洋竹草22 ℃处理与30 ℃处理差异不显著(P>0.05),而与38 ℃差异显著(P<0.05)。干旱处理10、15 d时不同温度处理均与对照差异显著(P<0.05)。随着天数的增加,叶长变小,同时与对照差异显著(P<0.05),说明洋竹草叶片长度随着温度的升高、胁迫天数的增加而变小。
2.3 高温胁迫对洋竹草叶片厚度的影响
由图3可知,各处理中以38 ℃胁迫15 d时叶片厚度最厚,达到0.84 mm,与对照差异显著(P<0.05)。高温30、38 ℃胁迫7、10 d时差异不显著(P>0.05),与对照差异显著(P<0.05),表明高温胁迫能增加洋竹草叶片的厚度。
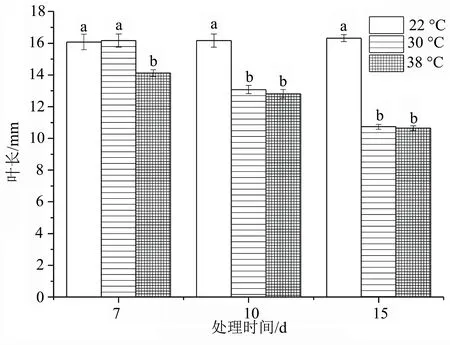
图2 高温胁迫下洋竹草叶长的变化
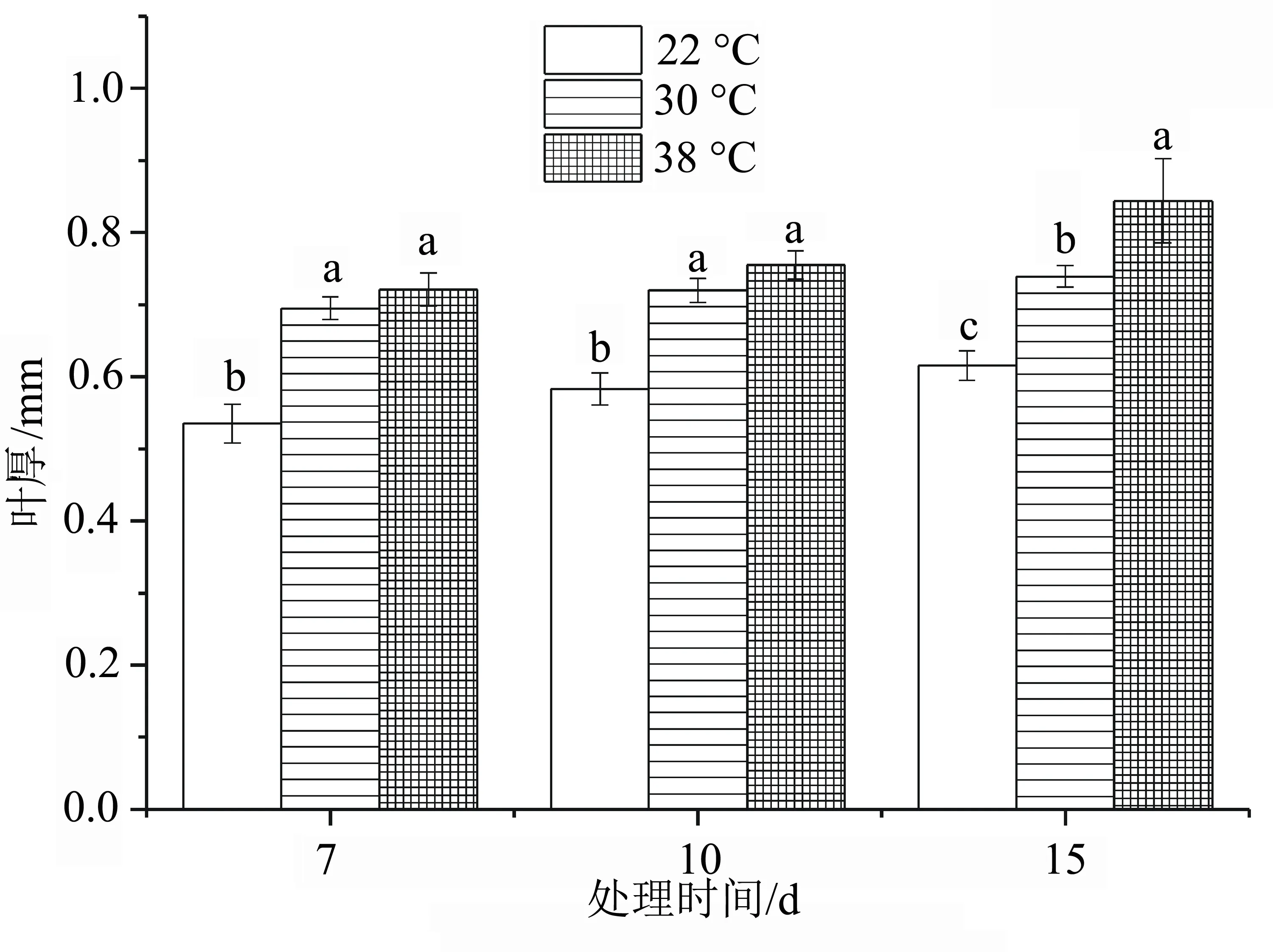
图3 高温胁迫下洋竹草叶厚的变化
2.4 高温胁迫对洋竹草相对电导率的影响
不同高温胁迫下,相对电导率在14.31%~48.53%。随着胁迫温度的升高,洋竹草相对电导率变化表现为持续上升的趋势,在高温胁迫7 d时,各温度处理差异均不显著(P>0.05);随着胁迫天数的增加,在高温胁迫15 d时,各温度处理的洋竹草相对电导率均差异显著(P<0.05),以高温38 ℃下最大。
2.5 高温胁迫对洋竹草丙二醛(MDA)含量的影响
植物在逆境条件下,会发生脂膜的过氧化,产生丙二醛,其浓度是判断脂质过氧化程度和膜系统伤害大小的指标[5]。除对照外,随着高温胁迫时间的延长,洋竹草叶片中MDA含量呈现持续增加的趋势,且在胁迫15 d时达到最大值,为3.11 mmol/g,为对照的1.31倍;高温30 ℃处理7、10 d时,MDA的含量与对照相比差异不显著(P>0.05);高温38 ℃处理10、15 d时,与对照差异显著(P<0.05),叶片中的MDA含量增加幅度较大,说明此时的高温胁迫对细胞膜造成了一定程度的伤害。
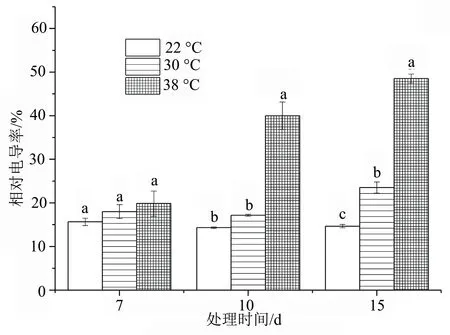
图4 高温胁迫下洋竹草相对电导率的变化
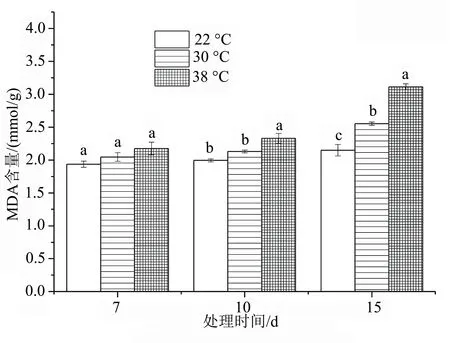
图5 高温胁迫下洋竹草丙二醛含量的变化
2.6 高温胁迫对洋竹草保护酶(SOD、POD、CAT)活性的影响
植物受到环境胁迫时,SOD酶是保护植物细胞免受自由基伤害的第一道防线,是清除活性氧的重要保护酶[6]。随着胁迫温度的升高,洋竹草叶片SOD活性呈逐渐上升的趋势(图6),在30 ℃高温胁迫处理下,SOD活性随着胁迫天数的延长轻微上升,且分别比CK上升了6.157%、5.843%和19.898%,在高温胁迫7、10 d处理下与CK差异不显著(P>0.05);在高温胁迫15 d处理下与CK差异显著(P<0.05);在38 ℃高温胁迫处理下随着胁迫时间的延长,SOD活性变幅不大,但与22、30 ℃均差异显著(P<0.05),且分别比CK上升了68.241%、68.928%和82.653%。
由图7可以看出,高温胁迫下不同天数处理对洋竹草叶片POD活性有影响。洋竹草的POD活性随着高温胁迫时间的增加呈缓慢上升趋势,且始终高于对照组。显著性分析表明,30 ℃和38 ℃处理下的洋竹草的POD活性在高温胁迫7、10 d时均差异不显著(P>0.05),在胁迫15 d时差异显著(P<0.05)。
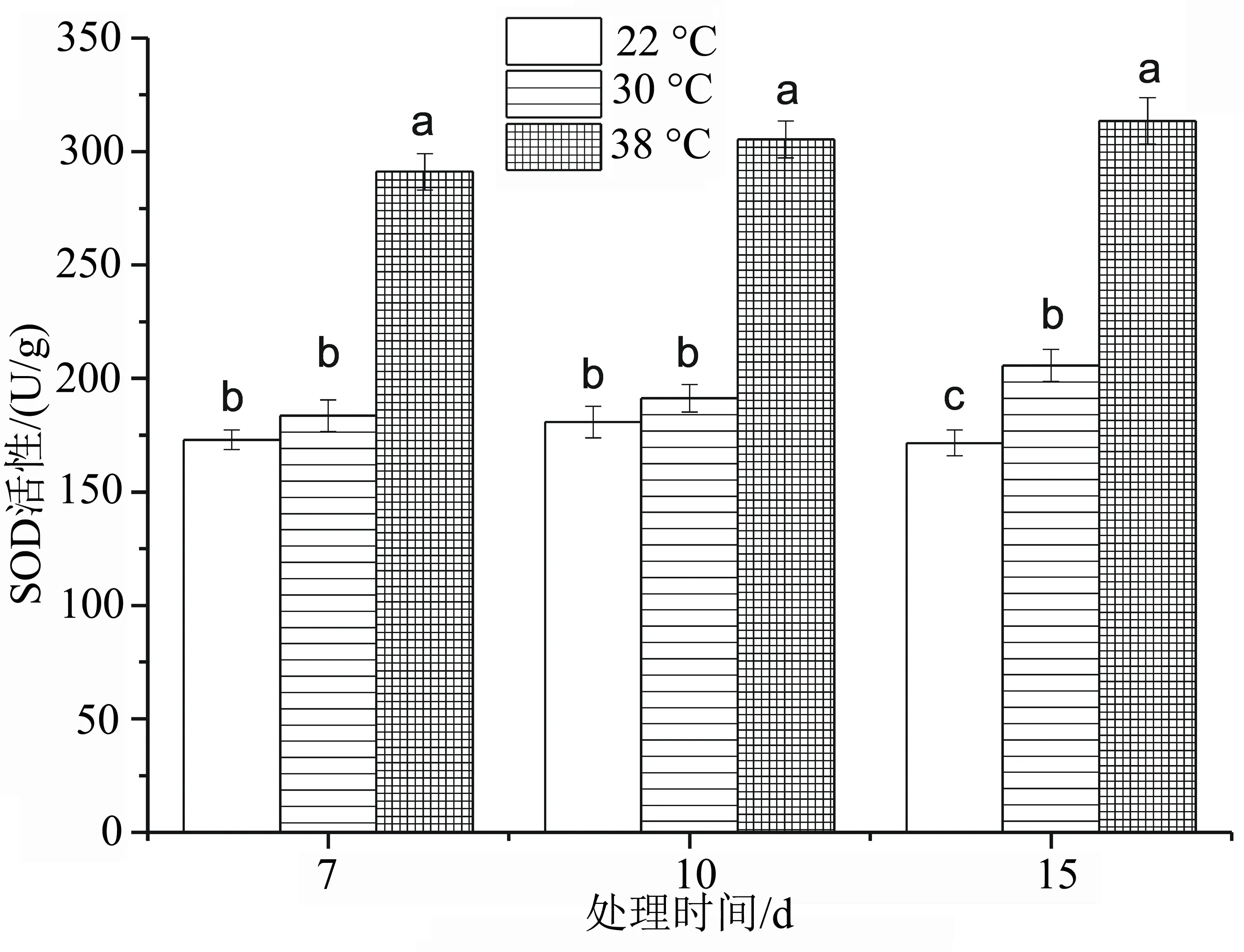
图6 高温胁迫下洋竹草超氧化物歧化酶SOD含量的变化
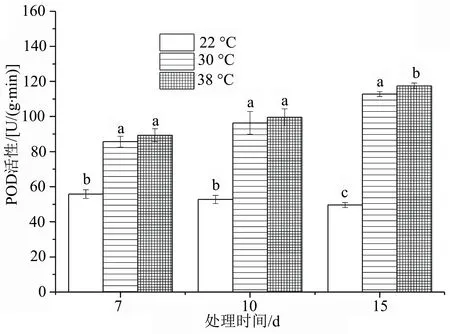
图7 高温胁迫下洋竹草过氧化物酶POD活性的变化
与CK相比,高温30 ℃胁迫处理下,CAT活性缓慢增加(图8),且随着胁迫天数的增加,CAT活性上升。高温38 ℃处理15 d时,与处理10 d相比发生了小幅下降;高温30、38 ℃两个温度处理,在高温胁迫15 d时差异不显著(P>0.05),在高温胁迫7、10 d处理下,两个温度处理差异显著(P<0.05)。
3 讨论与结论
3.1 高温胁迫对洋竹草叶片形态的影响
作为拥有最大接触面积和环境刺激最敏感的器官之一,叶的功能性状与资源的利用能力和效率密切相关[7-8];对植物叶片功能性状的研究可以更加深入了解植物对其储存资源的分配形式,探索出植物对生境的适应策略[9]。叶片厚度能够指示植物的适应对策,通常叶片较薄的植物属于开拓性策略,主要用于投资生长速率和资源获取能力;叶片较厚的植物属于保守性策略,主要用于投资养分储存效率以获得竞争优势[10-11],比如西南地区野生马蹄金叶片厚度会随着干旱胁迫的加深而变薄[12];黑麦草通过增加叶片的厚度来提高抗旱性[13]。本研究发现,洋竹草叶片应对高温反应时表现为叶片厚度加厚,叶片长度、宽度减小,叶长和叶宽的变化趋势大致相同,变化大小幅度不同。高温30 ℃处理7 d时,叶长及叶宽均与对照差异不显著,而高温38 ℃在不同天数的处理下叶长、叶宽及叶厚均与对照差异显著,说明洋竹草在高温胁迫下叶片产生自我保护的行为来防止高温的伤害。
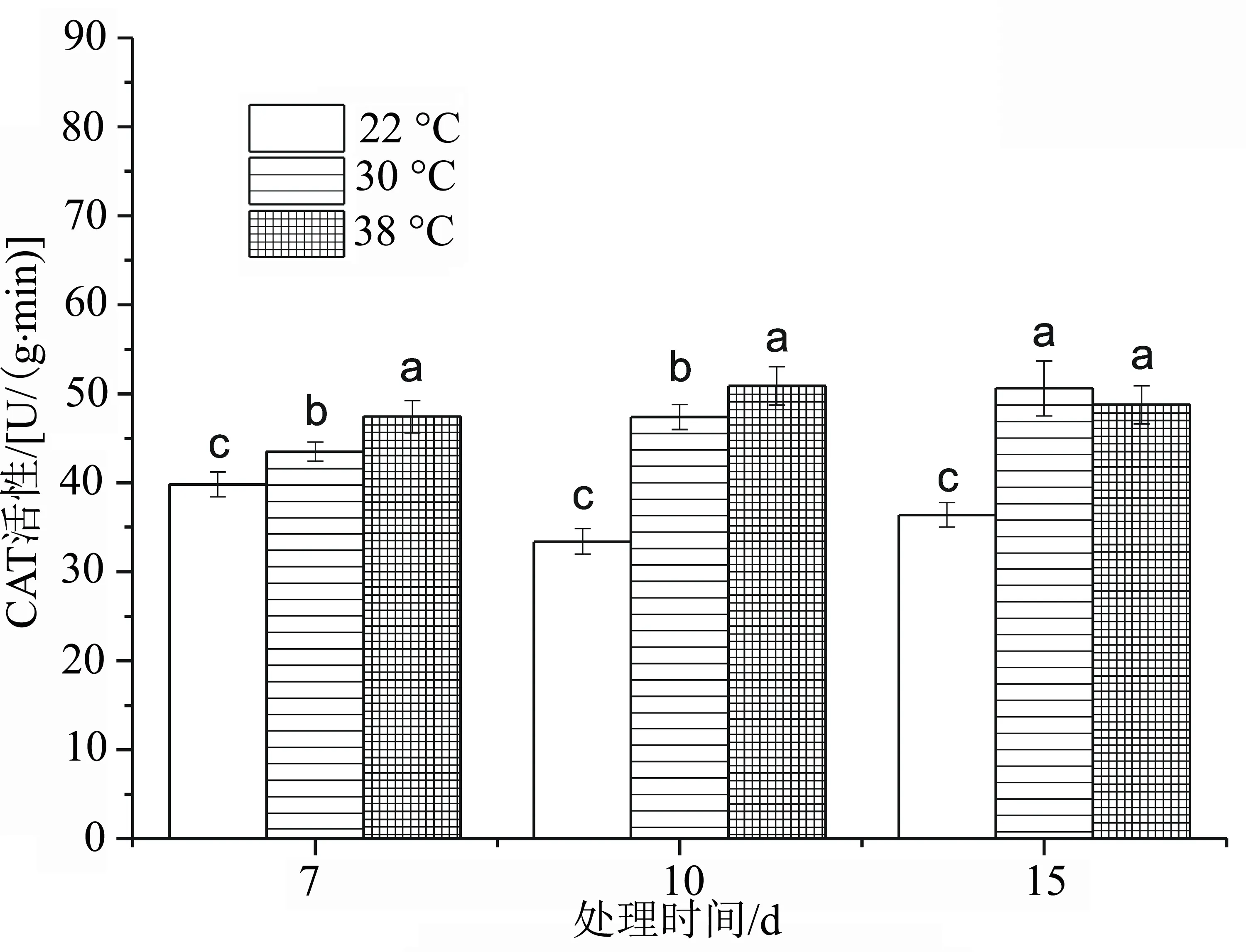
图8 高温胁迫下洋竹草过氧化氢酶CAT活性的变化
3.2 高温胁迫对洋竹草生理生化的影响
高温对植物的伤害最先表现是细胞膜系统损伤,膜的稳定性可作为植物抗热性的指标,丙二醛是细胞膜脂化产物,其含量的高低间接反映了植物组织的抗氧化能力[14-16];处于逆境中的植物会出现主动的适应调节,植物为缓解高温逆境造成的伤害,会激活体内保护酶活性的提高,减少细胞膜的脂质过氧化作用的程度,从而提高细胞膜的稳定性。在高温胁迫的条件下,植株体内活性氧的产生和清除动态平衡的机制遭到破坏,致使相对电导率和丙二醛含量增加[17]。在本试验中,对照处理下的丙二醛含量相对稳定,而不同高温胁迫下洋竹草叶片的丙二醛质量摩尔浓度与对照相比则显著增加,且其质量摩尔浓度大体上表现出随高温胁迫时间的延长而增加的趋势(图5)。本研究中,洋竹草叶片中的丙二醛含量和相对电导率变化趋势相同,均表现为随着胁迫天数的增加而上升的趋势,30、38 ℃胁迫7、10 d时与对照差异不显著(P>0.05),胁迫15 d时与对照差异显著(P<0.05),说明洋竹草在高温初期具有一定的耐热性,但持续的高温对洋竹草造成了一定的伤害。
逆境胁迫会破坏植物内生产和消除活性氧的动态平衡,活性氧物质在细胞中的积累导致细胞受到氧胁迫[18-19];超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)等是酶反应系统里重要的保护酶[20-21],因此可以通过高温胁迫下的抗氧化物质的活性来判断植物的耐热性,从图6~图8可知,洋竹草叶片中的抗氧化酶随着高温胁迫时间的延长均表现出较高的活性,来阻止高温逆境对自身造成的伤害,特别是高温38 ℃处理时,洋竹草叶片中SOD、POD活性均达到最大,在高温胁迫30 ℃时,SOD、POD活性均较对照高。在胁迫初期时,SOD活性与对照差异不显著,说明洋竹草对30 ℃反应不是太敏感,具有一定的耐热性;而POD活性一直较高,CAT活性在高温胁迫15 d时发生了下降,可能是与CAT的抗修复能力具有一定的阈值有关[22]。
通过以上分析,本研究认为,洋竹草可以承受高温38 ℃、处理10 d的胁迫,主要是与洋竹草本身及具有较高的渗透调节能力和较强抗氧化能力有关;短期的高温胁迫(7 d),植株的生长状况和对照无明显差别。因此,洋竹草在管理养护时,可以进行短期的高温胁迫,在38 ℃持续时间超过15 d时,应及时降温。
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
农业工程学报(2022年1期)2022-03-25
中国核电(2021年3期)2021-08-13
英语文摘(2021年3期)2021-07-22
小星星·阅读100分(低年级)(2019年10期)2019-11-26
安徽文学(2019年6期)2019-06-28
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
