
莱州湾金城海域网采浮游植物年际变化及与环境因子的关系
2019-05-31 00:51宋秀凯何健龙李佳蕙于广磊刘爱英
生态学报 2019年8期
宋秀凯,程 玲,付 萍,何健龙,李佳蕙,于广磊,刘爱英
山东省海洋资源与环境研究院山东省海洋生态修复重点实验室, 烟台 264006
浮游植物是海洋重要的初级生产力,海洋浮游植物群落结构的转变将改变浮游植物的生物量及生产力,继而对整个海洋生态系统的物质循环和能量传递产生重要影响[1]。浮游植物群落对环境变化响应敏感,因此群落的变动可以反映水质健康和气候变化。近年来,越来越多的研究表明[2- 4],受近海环境变化的胁迫,浮游植物群落结构正处于不稳定状态。
莱州湾地处渤海南岸,是渤海三大海湾之一,入海径流丰富,营养盐充足,是传统鱼、虾、蟹的产卵场和索饵场[5]。随着近几年养殖业的发展,莱州湾金城海域已发展成为山东省重要海湾扇贝养殖基地之一,但由于片面追求高产量,忽视长远的生态和环境效益,缺乏对水域生态养殖容量的科学了解和统一的规划管理,致使局部海域开发过度;另一方面由于养殖密度过高,造成生态环境质量下降,浮游植物的群落结构也随之发生了较大变化,严重影响海水养殖业的持续稳定发展。浮游植物作为双壳贝类重要的饵料生物,其种类和数量多寡将直接影响养殖贝类的产量。因此,研究该海域浮游植物种类和数量的年际变动,有助于了解养殖海域生态环境和养殖业的现状,解决养殖贝类死亡率高和出肉率低等问题。本文分析了2009—2010年连续两年24个月莱州湾金城海域贝类养殖区主要理化因子及网采浮游植物种类组成、数量分布和生物多样性变化规律,并通过典范对应分析结合多个环境因子探讨浮游植物与环境间的复杂关系,旨在较为系统地阐述近岸浮游植物的群落结构特征及演替规律,进一步深入研究莱州湾浮游植物的长期变化规律及海洋生态系统动态变化机制,为莱州湾浅海养殖业的合理开发、环境治理和修复提供基础数据。
1 材料与方法
1.1 样品采集
调查海域为莱州湾金城海域(119.9°—120.2°E,37.3°—37.6°N),共设15个采样站位(图1),该海域水深范围3.5—13.0 m,由南向北逐渐加深。于2009—2010年逐月进行共24个航次水质及浮游植物调查。采样船只为当地渔民渔船。
浮游植物采集及样品处理等均按《海洋调查规范》[6]进行,使用浅Ⅲ浮游生物网自底层至表层作垂直拖网采集,样品经鲁格氏溶液固定后实验室进行分类、鉴定、计数和统计。原则上每个样品鉴定到种的比例占总种类数的80%以上,每个样品镜检3次,每次镜检计数细胞个数不少于200个。各采样站位同步调查水温(WT)、盐度(Salinity)、透明度(Transparency)、化学需氧量(COD)、pH值、叶绿素a(Chl a)、总氮(TN)及总磷(TP)等环境参数,所有操作均按照《海洋监测规范》进行[7]。
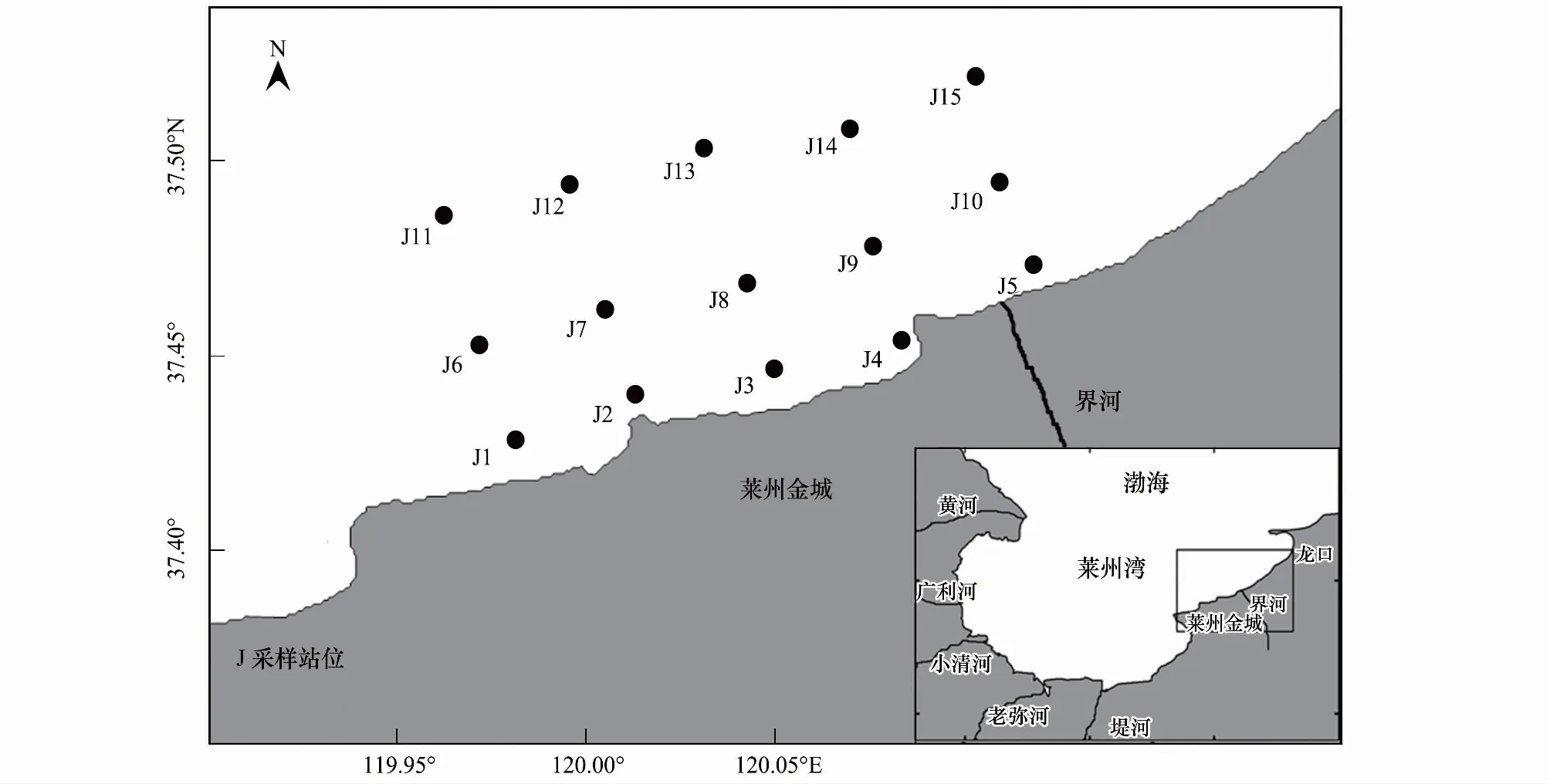
图1 莱州湾金城海域采样站位图Fig.1 Sampling sites in Jincheng area of Laizhou Bay
1.2 数据分析
1.2.1优势种、多样性指数、丰富度、均匀度分析
浮游植物丰度以每立方米出现的细胞数表示(个/m3)。
优势种的优势度:Y=(ni/N′)×fi
式中,ni为第i种的丰度,fi为该种在各站位中出现的频率,N′为总丰度。根据种类优势度公式计算各种生物的优势度,将Y>0.02的生物定为优势种。
物种丰富度指数采用Margalef指数(dMa)计算公式[9]:dMa=(S-1)/log2N
均匀度指数采用Pielou指数计算公式[10]:E=H′/lnS
式中,N为采集样品中所有物种的总个体数,S为样品中的种类总数,Pi为第i种的个体数与样品中的总个体数的比值。
1.2.2群落结构分析
多元统计方法包括聚类分析和典范对应分析(CCA)等,均是基于种类数据和环境数据集合的排序方法,已广泛应用于海洋生态群落结构分析中[11- 13]。本文选取水温、盐度、pH、透明度、化学需氧量、总氮、总磷和叶绿素a 8个指标分析其对浮游植物群落结构的影响。CCA分析中,浮游植物主要选取海区常见种,剔除出现频率小于15%的物种,物种及环境数据均采用对数log(x+1)转换。
全部数据的统计分析用SPSS 19.0软件完成,群落结构聚类分析使用PRIMER 6.0软件完成,典范对应分析采用CANOCO 4.5软件完成。
2 结果
2.1 浮游植物群落组成
本调查共鉴定出浮游植物81种,隶属于硅藻门、甲藻门、黄藻门和金藻门4门25科43属。其中硅藻门最多,共16科33属67种,占总种数的82.7%;甲藻门次之,共6科7属11种,占总种数的13.6%;黄藻门2科2属2种,占总种数的2.5%;金藻门1科1属1种。2009年和2010年金城海域浮游植物种类数基本一致,硅藻优势种主要以角毛藻属、丹麦细柱藻、菱形藻属为主,甲藻以夜光藻为主。2009年与2010年种类组成及比例基本一致,硅藻比例均在80%以上,甲藻为10%左右,其余为金藻和黄藻。与2009年相比,2010年硅藻所占比例略有升高,甲藻略有下降(图2)。
莱州湾调查海域浮游植物门类组成的周年变化如图3所示,除2009年5月及2010年5月、10月外,其余月份均为硅藻占绝对优势,比例均在80%以上。5月份甲藻比例明显升高,主要是夜光藻的大量出现;7、8、9月硅藻占绝对优势,几乎见不到其他藻类。周年变化显示调查海域浮游植物以硅藻为主要优势,季节演替不明显。
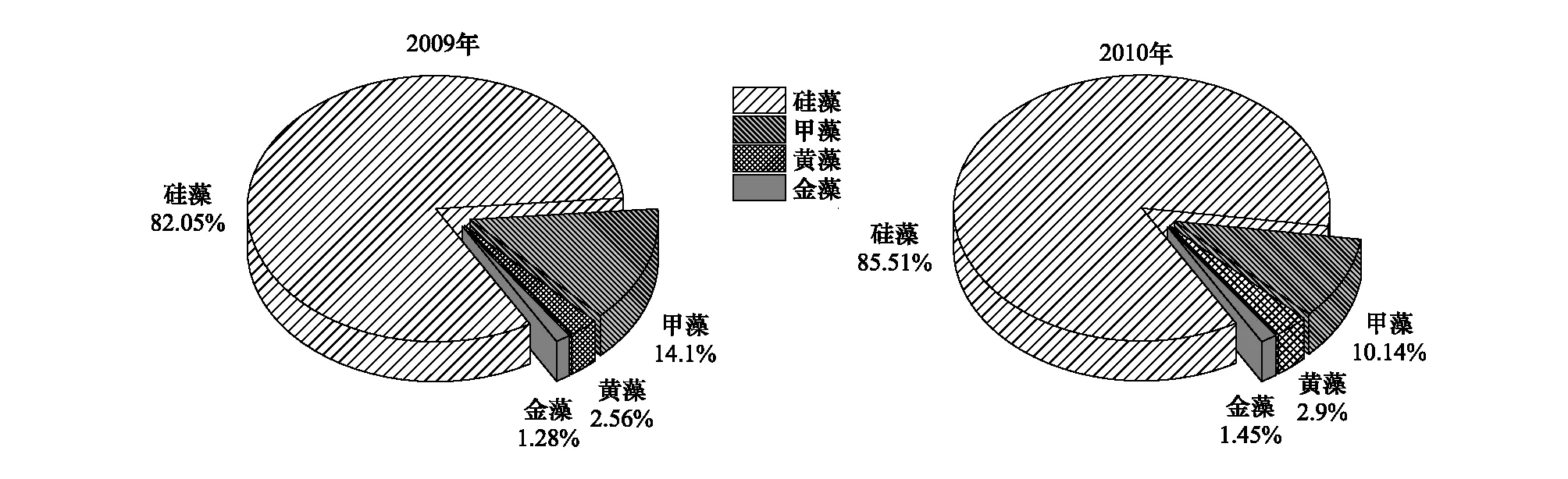
图2 2009—2010年浮游植物种类组成Fig.2 Specific composition of phytoplankton from 2009 to 2010

图3 2009—2010年浮游植物门类组成周年变化Fig.3 Annual variation in species composition of phytopiankton from 2009 to 2010
2.2 浮游植物群落结构
为直观展示各月份浮游植物丰度变化情况,浮游植物丰度周年变化采用常用对数形式表示。如图4所示,浮游植物丰度的年际变化呈现明显的双峰模型,2009年与2010年变化趋势基本一致,2月份达全年最高值,之后呈下降趋势,至5月为全年最低值,在9月到达到全年的第二个高峰。
浮游植物种类数年际变化与丰度相似亦呈双峰型变化趋势,2009年与2010年略有差别,除8月和10月外,其余月份2010年浮游植物种类数均小于2009年。
浮游植物多样性指数年际变化呈现明显的季节变化规律,高峰分别出现在3月和10月,2009年多样性指数最低值出现在7月,2010年最低值出现在9月。除2010年9月份外,生物多样性指数均在1.5—3.5的正常范围内。
浮游植物均匀度指数年际变化趋势显示:1—5月均匀度指数较为稳定,6—9月有所下降,10—12月有所升高并趋于稳定。除2010年8月和9月外,均匀度指数均大于0.3。
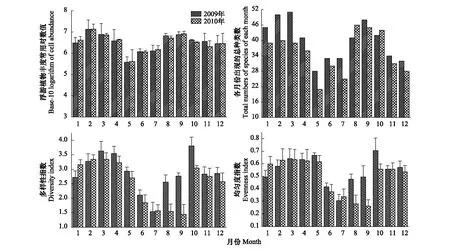
图4 2009—2010年浮游植物群落结构(细胞丰度、总种类数、多样性指数、均匀度指数)年际变化Fig.4 Phytoplankton community variation including cell abundance, species, diversity and evenness from 2009 to 2010
2.3 优势种群动态变化
海区主要优势种季节更替明显(表1)。春季(3—5月)主要优势种为尖刺拟菱形藻(Y=0.0228—0.217)、夜光藻(Y=0.0282—0.293)、旋链角毛藻(Y=0.0902—0.217)等;夏季(6—8月)主要优势种为洛氏角毛藻(Y=0.167—0.776)、尖刺拟菱形藻(Y=0.0233—0.203)、丹麦细柱藻(Y=0.108—0.276)等;秋季(9—11月)主要优势种为丹麦细柱藻(Y=0.0416—0.262)、洛氏角毛藻(Y=0.0559—0.739)、中肋骨条藻(Y=0.0285—0.436)等;冬季(12—2月)主要优势种为尖刺拟菱形藻(Y=0.0417—0.486)、中肋骨条藻(Y=0.0725—0.442)、旋链角毛藻(Y=0.0389—0.281)等。
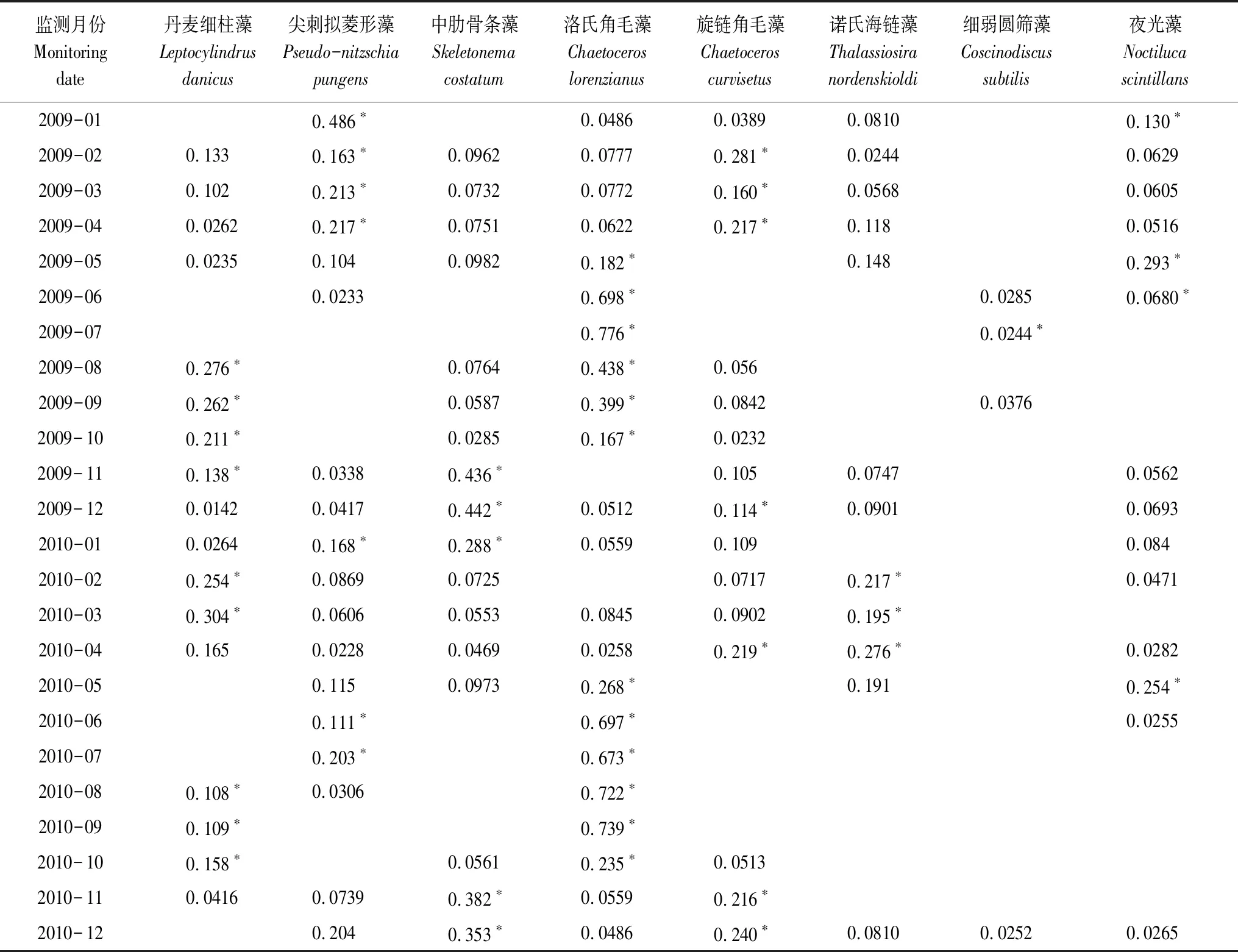
表1 主要优势种的优势度年际变化
优势度>0.02的物种为该航次的优势种,表格中带*的数字为该月份海区前两位优势种的优势度
除了主要优势种群随季节演替明显外,各主要优势种丰度随季节变化也十分明显,由图5可以看出,主要优势种(除洛氏角毛藻)均在2月达全年丰度的最高值,5月各优势种丰度均较低,与浮游植物丰度年际变化趋势较为一致;8、9月洛氏角毛藻、丹麦细柱藻丰度相对较高,是造成8、9月浮游植物丰度出现另一高峰的主要原因。2009年与2010年各优势种丰度年际变化基本一致。

图5 2009—2010年主要优势种的丰度动态变化Fig.5 Cell abundance variation of dominate species from 2009 to 2010
2.4 浮游植物群落聚类分析
对各个航次的浮游植物细胞丰度进行平方根转化,做出Bray-Curtis相似性矩阵,以此矩阵进行CLUSTER聚类和MDS标序,结果见图6。调查海域浮游植物群落在60%相似性水平大致可分为4个类群,同时使用SIMPER分析了各组分的优势物种。第Ⅰ类群以5月份样品为代表,第Ⅱ类群包括11、12、1、2、3、4月份样品,第Ⅲ类群包括8、9、10月份样品,第Ⅳ类群包括6、7月份样品。
类群Ⅰ:2009年5月和2010年5月浮游植物群落相似性约89.6%,此月份浮游植物丰度偏低,种类数较少,甲藻丰度占比较高,夜光藻、诺氏海链藻、洛氏角毛藻和丹麦细柱藻等为特征种,贡献率分别为15.7%、12.4%、9.6%和6.4%,累计贡献率为44.1%。此时,海区的水温(15.8±0.9)℃、盐度30.992±1.281、总氮(0.621±0.013) mg/L、总磷(0.0255±0.0083) mg/L。
类群Ⅱ:2009和2010 年每年11月至次年4月浮游植物群落相似性约52.7%,该时间内水温相对较低,硅藻为主要优势种,特征种主要为尖刺拟菱形藻、中肋骨条藻、旋链角毛藻等,贡献率分别为10.3%、7.4%和6.6%,累计贡献率24.3%。此时,海区的水温(5.5±2.7)℃、盐度30.354±0.607、总氮(0.802±0.272) mg/L、总磷(0.0265±0.0086) mg/L。
类群Ⅲ:2009和2010年每年8月至10月的浮游植物群落相似性约59.8%,此时间范围水温较高,特征种为丹麦细柱藻、洛氏角毛藻、掌状冠盖藻等,贡献率分别为10.6%、9.8%和6.6%,累计贡献率21.0%。此时,海区的水温(23.2±4.3)℃、盐度30.614±0.264、总氮(0.685±0.235) mg/L、总磷(0.0431±0.0143) mg/L。
类群Ⅳ:2009和2010年6月、7月浮游植物群落相似性约59.6%,特征种为尖刺拟菱形藻、洛氏角毛藻、圆柱角毛藻和大西洋角毛藻等硅藻,贡献率分别为9.9%、8.7%、8.4%和6.6%,累计贡献率为19.6%。此时,海区的水温(25.4±0.7)℃、盐度30.914±0.427、总氮(0.664±0.112) mg/L、总磷(0.0227±0.0142) mg/L。

图6 金城海域24个航次浮游植物群落的聚类树状图和多维尺度转换排序(MDS, non-metric Multidimensional Scaling)Fig.6 Cluster and MDS of phytoplankton community on 24 monitoring dates in Jincheng aera
2.5 浮游植物群落结构与环境因子关系
为探讨浮游植物与环境因子的关系,对浮游植物主要优势种与水温等8项环境指标做相关性分析,结果如表2所示。水温与洛氏角毛藻丰度呈极显著正相关(R=0.613,P<0.01),与旋链角毛藻、中肋骨条藻和夜光藻丰度均呈极显著负相关(R=-0.662, -0.649, -0.649,P<0.01);化学需氧量与旋链角毛藻和中肋骨条藻均呈显著负相关(R=-0.589, -0.480,P<0.05);盐度与中肋骨条藻呈极显著负相关(R=-0.585,P<0.01);其他环境因子与各优势种丰度相关性不明显。
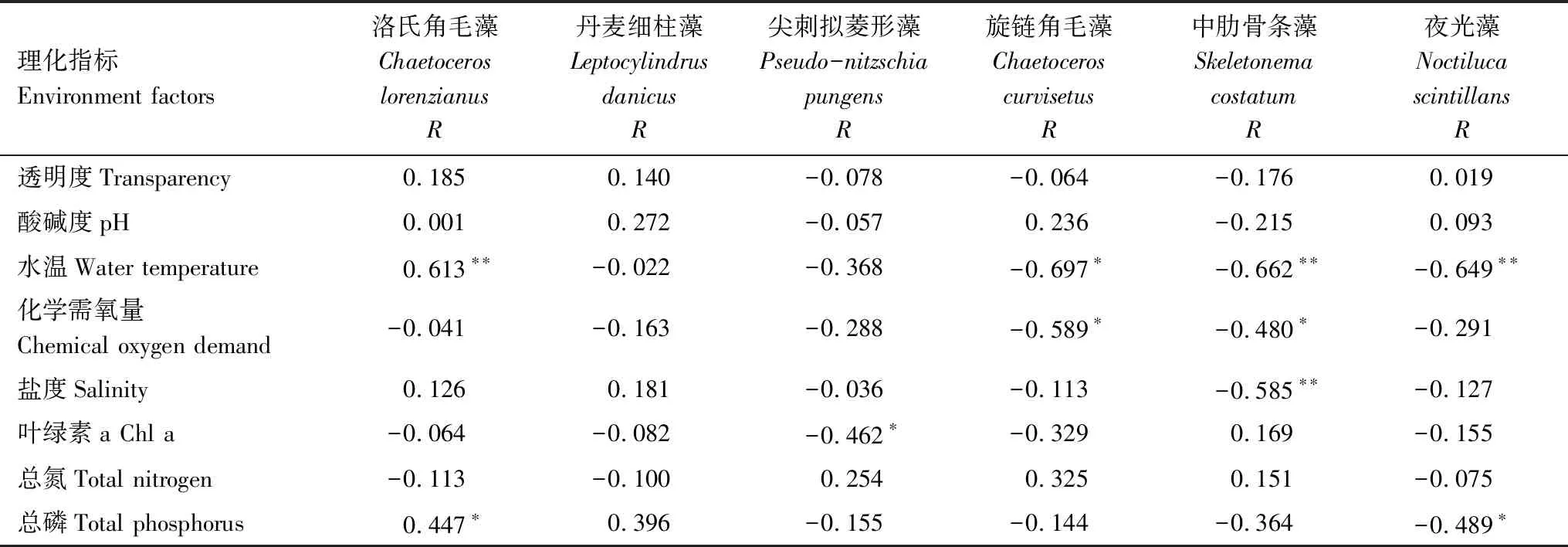
表2 主要优势种丰度与环境因子的相关性
*0.05显著水平(双尾),**0.01显著水平(双尾)
为了进一步探讨环境因子与浮游植物群落结构的关系,对主要优势种与8种环境因子进行CCA分析。由图7可以看出,影响浮游植物群落结构最主要的因素是水温,其次是盐度,化学需氧量、总氮、总磷、透明度及叶绿素a对浮游植物的影响较小。不同种类的浮游植物对环境因子的要求不同,夜光藻、旋链角毛藻等多为低温低盐种,洛氏角毛藻则为高温高盐种;故而在水温较高的8、9月份洛氏角毛藻丰度较高,而在1、2月份旋链角毛藻、夜光藻等丰度较高。丹麦细柱藻与盐度及pH均呈现较强的正相关。

图7 浮游植物优势种与环境因子典范对应分析 Fig.7 Canonical correspondence analysis (CCA) of dominant species and associated environmental factorsCLOR:洛氏角毛藻Chaetoceros lorenzianus;LDAN:丹麦细柱藻Leptocylindrus danicus;CCUR:旋链角毛藻Chaetoceros curvisetus;PPUN:尖刺拟菱形藻Pseudo-nitzschia pungens;SCOS:中肋骨条藻Skeletonema costatum;NSCI:夜光藻Noctiluca scintillans;TNOR:诺氏海链藻Thalassiosira nordenskioldii;HSUB:细弱圆筛藻Coscinodiscus subtilis;GSTR:斯氏几内亚藻Guinardia striata;透明度Transparency;pH:酸碱度pH value;WT:水温Water temperature;COD:化学需氧量Chemical oxygen demand;Chl a.:叶绿素a Chlorophyll a;TN:总氮Total nitrogen;TP:总磷Total phosphorus;盐度Salinity
3 讨论
3.1 浮游植物群落组成及参数变化
莱州湾浮游植物种类众多,有关其种类组成的研究曾有过记载[14-15]。本次莱州湾金城海域调查初步鉴定浮游植物(含变型及变种)共43属81种,隶属于4大门类,多于1997年的62种[5],其中硅藻门无论丰度还是种类数均占绝对优势,这与陈碧娟等[5]2001年的研究结果一致。2009年和2010年莱州湾调查海域浮游植物丰度变化范围分别为38.4×104—1351×104个/m3和41.8×104—1380×104个/m3,1992—1993年莱州湾浮游植物丰度变化范围28×104—1780×104个/m3,而1982—1983年莱州湾浮游植物丰度变化为27×104—3629×104个/m3,莱州湾浮游植物丰度较20世纪90年代有所降低[15-16]。
浮游植物丰度分布的季节变化明显,高峰分别出现在2月和9月,最低值出现在5月。王俊和康元德对1992—1993年渤海浮游植物的调查结果表明,浮游植物丰度最高峰分别出现在上半年的2月份和下半年的10月份,5月为全年最低值[15];刘慧等对莱州湾养殖海区2001—2002年浮游植物的研究显示早春2—3月是细胞丰度的高峰,最低值出现在4—5月[17];均与本次调查结果一致。莱州湾处于北半球中纬度温带海,浮游植物的季节变化应符合温带海域浮游植物两周期的季节变化类型,渤海季节变化特点是上、下半年各有一次数量高峰[18]。本次调查结果符合这一结论,说明莱州湾多年来浮游植物数量的年际变化规律基本一致,未发生较大波动。
浮游植物多样性指数和均匀度指数是其生物群落结构的重要参数,通常情况下多样性指数小于1、均匀度小于0.3可认为群落结构受到了其他环境因素的扰动[19],莱州湾金城海域浮游植物多样性指数均大于1,均匀度指数均大于0.3,该海域浮游植物群落结构相对稳定。
3.2 浮游植物群落聚类及主要优势种变化
2009—2010年莱州湾金城海域浮游植物群落聚类分为四大类:Ⅰ类为5月份样品,2009年和2010年5月样品相似度达80%以上,优势种为夜光藻和洛氏角毛藻,尤其夜光藻,在数量上占绝对优势,与刘慧等[17]2001年莱州湾东部养殖区浮游植物的调查结果一致;Ⅱ类11月—4月样品为冬春季样品,优势种有中肋骨条藻、旋链角毛藻和夜光藻;Ⅲ类8、9、10月样品主要为夏秋季样品,优势种主要包括洛氏角毛藻和丹麦细柱藻;Ⅳ类为6、7月样品,优势种为洛氏角毛藻和尖刺拟菱形藻。全年优势种与1982年的结果有较大不同,1982年莱州湾的主要优势种为中肋骨条藻、透明辐杆藻和日本星杆藻[20],2001—2003年度的主要优势种为中肋骨条藻、角毛藻、柔弱根管藻和斯氏几内亚藻[17]。
3.3 环境因子等对浮游植物群落的影响
莱州湾金城海域地处中纬度地区,季节分明,因此温度尤其是表层水温对生物的影响较大,典范对应分析显示,莱州湾金城海域影响浮游植物群落结构最主要的因素是水温,这与以往的研究结果一致[21-22]。此外,与温度呈正相关的优势种主要为洛氏角毛藻,而夜光藻、旋链角毛藻等与温度呈负相关,这与浮游植物优势种随季节变化的演替规律一致。此外,分析显示总氮、总磷对浮游植物的影响较小,调查海域主要为扇贝养殖区,不存在氮磷限制现象。有研究发现扇贝养殖促进了浮游植物特别是硅藻的繁盛,这可能是由于扇贝的滤食作用以及扇贝养殖区小生境中丰富的微生物,促进了浮游植物必须的营养盐及微量元素的循环供应[17,23]。另外,结合养殖的实际情况,5月为扇贝海上挂苗期,此时浮游植物丰度为全年最低,尤其作为扇贝饵料藻的硅藻类比例下降,会影响扇贝的产量和质量,因此要合理控制养殖规模,防止因饵料缺少导致贝类死亡。
猜你喜欢
英语文摘(2022年5期)2022-06-05
海洋通报(2021年1期)2021-07-23
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
石材(2020年6期)2020-08-24
流行色(2019年10期)2019-12-06
南水北调与水利科技(2018年2期)2018-07-05
安徽农业科学(2018年18期)2018-05-14
科学启蒙(2017年2期)2017-04-27
河北渔业(2016年9期)2016-11-05
