
基于中红外光谱法土壤理化性质与稳定性同位素剖面分布
2019-05-22 07:05:06马怀霞GbadamassiDossa许建初
水土保持研究 2019年3期
马怀霞, 阎 凯, Gbadamassi G.O.Dossa, 许建初
(1.中国科学院 昆明植物研究所东亚植物多样性与生物地理重点实验室, 昆明 650201;2.中国科学院 昆明植物研究所山地生态研究中心, 昆明 650201;3.中国科学院大学, 北京 100049; 4.世界农用林业中心, 昆明 650201)
随着人类不断扩大土地的利用范围,加剧对土地利用的强度,不同的土地利用方式也更显著的影响着土壤理化性质。在许多热带—亚热带发展中国家,土地利用方式正在快速变革,尤其是东南亚,大面积的热带被改造成农业用地,由此带来的土壤理化性质的变化是值得我们深入探究的。老挝是东南亚唯一的内陆国,农业人口占全国人口的90%左右,农业作为老挝的支柱型产业,占国内生产总值的51%[1]。其境内80%地区为山地和高原,潜在耕地面积约800万hm2,而实际耕地面积约80万hm2,总利用率非常低,仅约10%[2]。老挝北部多山,与中国云南接壤,人文差异小,土地资源丰富,但是受农业发展水平的限制,土地利用率低。了解老挝北部山区的土壤养分状况及其分布特点,对可持续开发和利用老挝的土地资源具有导向型作用[3]。
近年来,关于不同土地利用方式对土壤养分含量及其垂直分布的影响,不少学者分别基于不同研究区域探讨了土地利用方式对土壤养分的影响,结果表明土壤理化性状的变化与土地利用方式有密切关系[45]。土壤有机碳和氮素是土壤肥力的物质基础,有机碳和氮素作为土壤微生物能源和最主要营养元素,能够驱动土壤中碳氮磷硫等养分的转化和循环,影响土壤的理化性质以及养分供应能力[6]。并且土壤有机碳是控制土壤肥力和农业生产的关键因素,土壤有机碳的流失可以导致土壤肥力和土壤质量下降,影响陆地生态系统的稳定性[7]。土地利用变化是影响陆地生态系统碳氮循环的最大因素之一,也是仅次于化石燃烧而使大气CO2浓度急剧增加的最主要人为活动[8]。人为活动引起的土地利用变化可能会通过与生境扰动相结合改变进入土壤的植物残体数量与种类来间接影响土壤有机质的累积和分解速率,最终造成土壤总有机碳和全氮含量的变化[9],进而影响其他养分的循环和转化。另外,土壤有机质稳定碳氮同位素组成(δ13C 和δ15N 值)能有效地阐明土壤有机碳和氮的迁移与转化以及评估土壤有机质分解程度,是研究土壤碳氮循环的有力工具。有研究表明,土地利用变化还会影响不同粒径土壤颗粒之间的转化和再分布,而土壤有机碳和全氮含量的变化在不同粒径土壤中对土地利用变化的响应也会有所差异[10]。因此,仅对土壤中有机碳和全氮进行测定显然很难及时、准确反映不同土地利用类型下土壤质量的内在差异。
漫反射红外光谱是对固体样品粉末进行直接测量,操作简便、快速,能够非破坏的对各种样品进行快速、精确的分析,加上分析仪器的数字化和化学计量学方法的发展,运用化学计量学方法已能很好地进行光谱信息的提取及解决背景干扰方面的影响。其中中红外漫反射光谱比近红外漫反射光谱的谱带信息更强,更稳定,信息提取更容易。
目前受常规分析耗时费用昂贵的限制,鲜有研究针对较大研究区域进行多样本取样,同时对土壤的多项理化性质进行分析,本研究选择老挝北部的乌多姆赛省的孟本为研究区,利用中红外漫反射光谱法快速分析土壤理化性质,研究竹林、耕地、休耕地、橡胶林、森林和次生林6种土地利用方式对土壤理化性质的剖面分布的影响,以期为可持续开发利用合理管理老挝土地资源提供参考依据。
1 研究区概况
老挝北地势高低起伏,地貌类型多丘陵山地。海拔范围为196~2 266 m,坡度在40°以内范围,属热带季风气候,年平均温度高于20 ℃[11]。选择老挝北部的乌多姆赛省的孟本地区(东经101.79°—101.87°,北纬 20.41°—20.49°)作为监测和调查样区,选取一个10 km×10 km的区域作为试验调查样地,包含16个2.5 km×2.5 km的小区域,利用程序随机在每个小区域内随机选取一个圆心,以564 m为半径的圆圈范围作为一个群集(cluster),再次利用程序在每个群集中随机选取10个0.1 hm2(1 000 m2)的样地(plots)。本试验在孟本地区总共有160个双重随机嵌套分层抽样的样地,双重随机嵌套分层抽样的设计方法可以最小化由便利抽样所带来的偏差,可以多维度的了解不同空间尺度复杂的生态系统。这160个样地包含的土地利用类型有森林(61个),次生林(41个),橡胶林(4个),竹林(3个),耕地(41个),休耕地(10个)森林是30 a以上的热带雨林,主要树种有紫檀属(Pterocarpus)、石柯属(Lithocarpus)、栗属(Castanea)、龙船花属(Ixora)、茜草属(RadixRubiae)、木姜子属(Litsea)、火焰树属(Pathodea)、无患子属(Sapindus)、银柴属(Aporusa)等,当地村民只是在森林中进行薪材和蘑菇的收集,绝大多数森林没有任何人为活动。次生林是停止耕种4~10 a的退耕还林地,现在只是用于薪材和蘑菇的收集或者无人为干预,主要树种有黄牛木属(CratoxylumBI.)、紫薇属(Lagerstroemia)、血桐属(MacarangaThou.)、番荔枝属(Annona)、茜草属(RadixRubiae)、巴豆属(Croton)和椴树属(Tilia)等。橡胶林均是单一种植橡胶,耕种年份在3~7 a间,施用除草剂。竹林是受村民保护的原始森林,只是进行一些木材的收集,主要树种有无患子属(Sapindus)、牡荆属(Vitex)、榕属(Ficus)、茜草属(RadixRubiae)、紫薇属(Lagerstroemia)、楝属(Melia)、山矾属(SymplocosJacp)、倒吊笔属(Wrightia)、锦葵科(Malvaceae)、木姜子属(Litsea)、巴豆属(Croton)、龙船花属(Ixora)、栲属(Castanopsis)、番荔枝属(Annona)、叶下珠属(Phyllanthus)、柯属(Lithocarpus)和黄檀属(Dalbergia)等。耕地年限均小于50 a,属于刀耕火种的迁移农业,绝大多数耕地只种植了3~4 a玉米、水稻、西瓜和香蕉等农作物,还有一部分是刚刚进行了毁林开荒的刀耕火种作业。休耕地属于刀耕火种休憩地,主要种植玉米水稻2~4 a后停止耕种。
2 试验材料与方法
2.1 土壤样品采集与分析
于2015年9月在老挝孟本境内进行土壤样品采集。在去除土壤表面落叶等后,对6种土地利用方式土壤分4层进行采样:0—20 cm,20—50 cm,50—80 cm,80—110 cm。在20℃风干3 d,碾碎后过2 mm的聚乙烯筛以筛除碎石、卵石以及植物残骸,每份样品取15 g研磨后过0.5 mm 的聚乙烯筛。使用Bruker公司开发的ALPHA FT-IR 光谱分析仪进行土样的中红外漫反射光谱扫描,搭配使用光谱分析OPUS(Optics Users Software)软件进行光谱图的制作和基线校正导数转换,电脑运行R软件,通过R程序包和随机森林算法,进行光谱信息的识别及数字化转化、并与同一样品的常规检测结果进行比较,再通过机器学习建立数据库和建立模型后,“一键式”对待检土壤样品待测组分。土壤测定项目主要包括:土壤pH、有机碳(OC)、总氮(TN)、碳氮比、土壤全磷含量、土壤全钾含量、土壤不同颗粒(砂粒、粉粒、黏粒)的百分比组成、土壤稳定碳氮同位素(δ13C和δ15N)组成。
2.2 数理统计方法
样地数据采用R进行统计分析。采用单因素方差分析(One-Way ANOVA)对不同土地利用方式、不同土层之间土壤养分在α=0.05上的显著性差异进行分析和检验,不同处理间的多重比较采用scheffe成对检验法。
3 结果与分析
3.1 不同土地利用方式土壤pH剖面分异
所有用地类型中pH值均为酸性,与刘志强等人的研究一致[3]。所有土层中,耕地的pH都高于其他几个用地类型,并且在0—20 cm土层中,pH值大小为:耕地>竹林>休耕地>次生林>森林>橡胶林,耕地的pH值显著高于森林、橡胶林和次生林的(p<0.05);20—50 cm土层中,pH值大小为:耕地>竹林>休耕地>橡胶林>次生林>森林,耕地的pH值显著高于森林和次生林的(p<0.05),50—80 cm和80—110 cm土层中,pH值大小为:耕地>橡胶林>竹林>休耕地>次生林>森林,耕地的pH值显著高于森林和次生林的(p<0.05)。由于施肥的原因,导致耕地的pH高于其他用地类型。竹林、森林和次生林的pH较低,是因为土壤表层有大量的枯枝落叶,分解过程向土壤释放大量的有机酸,并且土壤腐殖质含量较高也导致土壤酸化,受热带地区水分温度的影响,土壤次生母质风化作用强烈,产生大量的铝离子,基本粒子的淋溶作用也加剧了土壤酸化。由图1可知,耕地的标准误差较大,这与耕地组内样地数量较多,并且各样地所耕作年份和种植作物不同有关。pH值在各土地利用方式下,随土层变化不大,差异并不显著(p<0.05)。
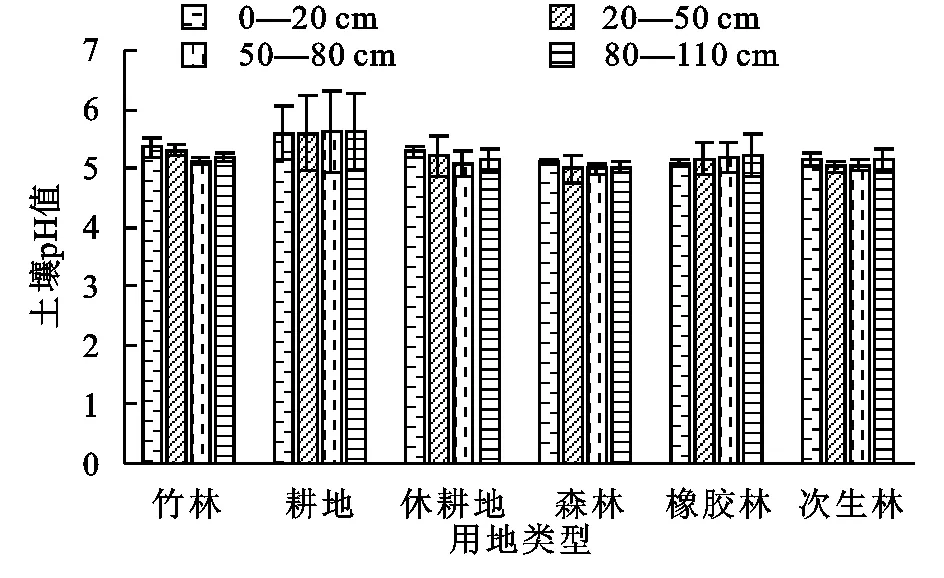
图1 不同土地利用方式下不同土层土壤pH值
3.2 不同土地利用方式土壤有机碳剖面分异
由图2所知,0—20 cm土层中,土壤有机碳含量的排序为:竹林(19.06 g/kg)>森林(16.46 g/kg)>次生林(14.70 g/kg)>耕地(14.48 g/kg)>休耕地(13.30 g/kg)>橡胶林(12.49 g/kg);20—50 cm土层中,土壤有机碳含量的排序为:竹林(12.55 g/kg)>森林(11.82 g/kg)>耕地(11.39 g/kg)>次生林(10.95 g/kg)>休耕地(10.15 g/kg)>橡胶林(10.01 g/kg);50—80 cm土层中,土壤有机碳含量的排序为:竹林(10.58 g/kg)>耕地(10.36 g/kg)>森林(10.35 g/kg)>次生林(10.04 g/kg)>休耕地(9.58 g/kg)>橡胶林(9.48 g/kg);80—110 cm土层中,土壤有机碳含量的排序为:竹林(12.55 g/kg)>耕地(11.82 g/kg)>次生林(11.39 g/kg)>休耕地(10.95 g/kg)>森林(10.15 g/kg)>橡胶林(10.01 g/kg)。可见4个土层中,橡胶林和休耕地的有机碳含量都较低,竹林有机碳含量都是最高的。因为休耕地经过多年的耕作,现在失去了耕作者施肥等管理措施,土壤养分没有得到及时的补充,土壤处于很贫瘠的状态。橡胶林因为使用除草剂,加上单一种植,也没有合理的进行施肥,故土壤养分非常缺乏。竹林是受到当地村民的保护的天然雨林,受人为干扰很少,生态系统没有受到破坏,大量的枯枝落叶,动植物残渣得以返回到土壤中,丰富的土壤根系和微生物群落可以加快土壤养分循环。随着土层加深土壤有机碳含量都在减少,但在不同土地利用类型中,各土层之间的显著性差异并不一致,但总体而言,表层土壤与底层土壤的有机碳含量都达到显著性差异(p<0.05),这与王洪杰等[11]研究一致。竹林、耕地、森林和次生林的表层土壤的有机碳含量组内标准差较大,相较于底层土壤,表层土壤的有机碳含量更易受表层动植物活动和地表降水径流的影响,从而带来较大的组内标准差。休耕地和橡胶林组内各样地的地表植被和地表土壤裸露情况较为一致,故组内标准差较小。
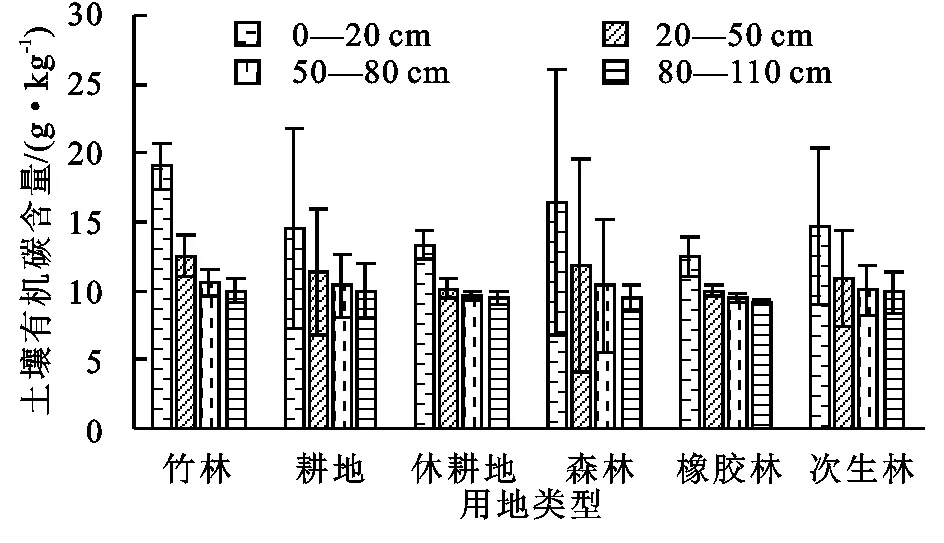
图2 不同土地利用方式下不同土层土壤有机碳含量
3.3 不同土地利用方式土壤总氮剖面分异
由图3可知,0—20 cm土层中,土壤总氮含量的排序为:竹林(1.62 g/kg)>森林(1.40 g/kg)>耕地(1.30 g/kg)>次生林(1.28 g/kg)>休耕地(1.18 g/kg)>橡胶林(1.14 g/kg);20—50 cm土层中,土壤总氮含量的排序为:竹林(1.20 g/kg)>森林(1.13 g/kg)>耕地(1.10 g/kg)>次生林(1.06 g/kg)>休耕地(1.03 g/kg)>橡胶林(1.02 g/kg);50—80 cm土层中,土壤总氮含量的排序为:竹林(1.10 g/kg)>耕地(1.06 g/kg)>橡胶林(1.02 g/kg)=次生林(1.02 g/kg)>森林(1.01 g/kg)=休耕地(1.01 g/kg);80—110 cm土层中,土壤总氮含量的排序为:耕地(1.04 g/kg)>竹林(1.03 g/kg)>次生林(1.02 g/kg)>橡胶林(1.00 g/kg)>休耕地(0.98 g/kg)>森林(0.96 g/kg)。由此可见,在0—110 cm内,土壤有机碳与总氮均值大小顺序及其在不同土地利用方式之间的显著性差异非常一致,这是因为土壤有机碳和总氮之间通常具有显著相关关系[12-13]。在各土层中均是休耕地与橡胶林的含量较低,竹林是总氮含量最高的土地利用方式,除了在80—110 cm土层中耕地略高于竹林,在各用地类型中,土壤总氮含量均随着土层加深而减少,且表层0—20 cm土层的总氮含量显著高于其他3个土层的总氮含量(p<0.05)。耕地的有机碳和总氮含量在50—110 cm土层中较高,这与耕作者施肥和秸秆还田等措施有关,施肥增加了土壤中氮素等养分输入,并显著提高土壤的有机质含量[14],大量养分下渗入深层土壤。因种植的玉米水稻等作物根系较短,无法吸收深层土壤中的养分,而表层土壤的养分被吸收的很充分,所以0—50 cm土层中,耕地碳氮含量低于竹林和森林。

图3 不同土地利用方式下不同土层土壤总氮含量
3.4 不同土地利用方式土壤碳氮比剖面分异
由图4可知,0—20 cm土层中,土壤碳氮比值的排序为:森林(12.11)>竹林(11.94)>次生林(11.91)>休耕地(11.64)>耕地(11.57)>橡胶林(11.43);20—50 cm土层中,土壤碳氮比值的排序为:森林(10.88)>竹林(10.80)>耕地(10.68)>次生林(10.59)>橡胶林(10.15)>休耕地(10.14);50—80 cm土层中,土壤碳氮比值的排序为:森林(10.43)>耕地(10.37)>次生林(10.19)>竹林(10.06)>橡胶林(9.91)>休耕地(9.84);80—110 cm土层中,土壤碳氮比值的排序为:耕地(10.23)>森林(10.19)>休耕地(10.13)>竹林(10.10)>次生林(10.04)>橡胶林(9.78)。土壤碳氮比受成土母质、地形因子及土壤形成后的土壤类型等结构性因素和施肥耕作种植方法等人为活动的随机性因素共同作用[15]。本研究主要探讨土地利用方式对土壤碳氮比空间变异的影响。综上可知,土壤碳氮比在各土层中均是休耕地和橡胶林的比值较低,森林、耕地和竹林的比值较高,所有用地类型的碳氮比值都在9~13,碳氮比不仅可以指示土壤有机质的降解程度,同时也是影响土壤有机质降解的重要因素,土壤碳氮比小于25时,微生物活动增强,易于有机质的降解[16]。通常碳氮比值大于25时,土壤有机质分解速率下降。该地区属于热带山区受高温高湿的气候影响,导致有机质降解速率快,这与碳氮比值较低相吻合。森林和竹林碳氮比值相对较高,这与其地表物质的输入水平有关,由于大量的植物凋落物的输入,森林和灌丛植被会增加土壤有机碳含量,同时减少土壤总氮含量,进而导致碳氮比值升高[17]。耕地碳氮比值高与农户对土地施用的肥料和管理水平普遍相关,并且该地区大量种植水稻,耕地长期处于低温淹水状态,有机质分解缓慢故而积累,从而使碳氮比处于较高水平。橡胶林和休耕地由于缺乏地表植被,有机碳分解较快,导致碳氮比较低。在各用地类型中,土壤碳氮比值均随着土层加深而减少,并且除竹林和橡胶林外,0—20 cm土壤的碳氮比均显著高于其他土层(p<0.05),竹林中各土层碳氮比无显著差异(p<0.05),橡胶林中,0—20 cm和20—50 cm的碳氮比显著高于其他两个土层(p<0.05)。
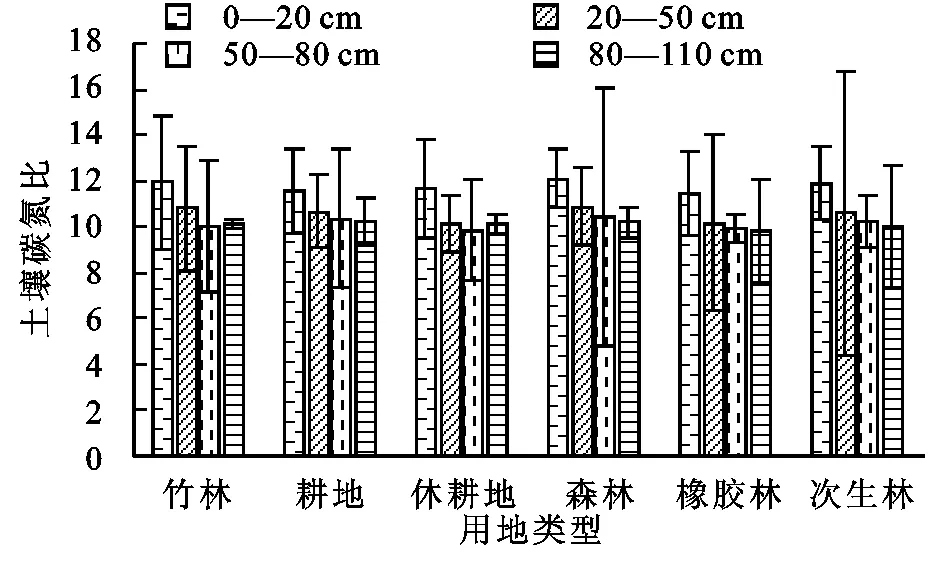
图4 不同土地利用方式下不同土层土壤碳氮比值
3.5 不同土地利用方式土壤磷钾含量剖面分异
由图5可知,0—20 cm土层中,土壤磷含量的排序为:竹林(0.47 g/kg)>森林(0.35 g/kg)>耕地(0.34 g/kg)=次生林(0.34 g/kg)>橡胶林(0.29 g/kg)>休耕地(0.28 g/kg),竹林的磷含量显著高于休耕地;20—50 cm土层中,土壤磷含量的排序为:竹林(0.38 g/kg)>耕地(0.31 g/kg)>次生林(0.30 g/kg)>森林(0.29 g/kg)>橡胶林(0.28 g/kg)>休耕地(0.27 g/kg);50—80 cm土层中,土壤磷含量的排序为:竹林(0.37 g/kg)>耕地(0.30 g/kg)>次生林(0.29 g/kg)>橡胶林(0.28 g/kg)=休耕地(0.28 g/kg)=森林(0.28 g/kg);80—110 cm土层中,土壤磷含量的排序为:竹林(0.35 g/kg)>次生林(0.30 g/kg)=耕地(0.30 g/kg)>休耕地(0.29 g/kg)>橡胶林(0.28 g/kg)>森林(0.27 g/kg)。土壤磷主要来源于土壤矿物,枯枝落叶及人为施肥。竹林是磷含量最高的土地利用方式,这与有机碳总氮的情况相似。除休耕地和次生林外,其余4种土地利用方式下的土壤磷含量基本上都随土层加深而减少,且森林中0—20 cm土层的磷含量显著高于其他土层(p<0.05),人类活动强度大,作物产量高,土壤表层枯枝落叶多是表层土壤磷含量高的主要因素,受雨水淋溶影响表层磷元素发生迁移转化,流失或沉积于土层下部,耕地与橡胶林磷含量垂直差异不明显,可能与地下水位高有关。休耕地和次生林中随土层加深磷含量先降低后升高,呈现出“U”形分布,表明磷含量与母岩密切相关[18]。磷含量在各土地利用方式中的组内标准差都较大,这可能与土壤磷含量受成土母质和地形影响极为显著有关。
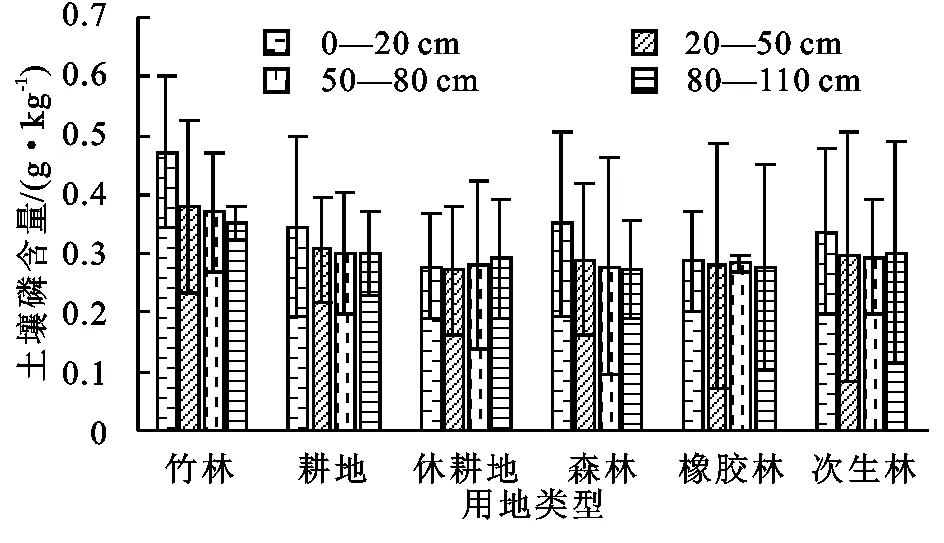
图5 不同土地利用方式下不同土层土壤磷含量
3.6 不同土地利用方式土壤钾含量剖面分异
由图6可知,0—20 cm土层中,土壤钾含量的排序为:竹林(15.68 g/kg)>次生林(11.65 g/kg)>耕地(11.06 g/kg)>橡胶林(10.54 g/kg)>休耕地(9.83 g/kg)>森林(9.76 g/kg),竹林的钾含量显著高于休耕地和森林;20—50 cm土层中,土壤钾含量的排序为:竹林(17.51 g/kg)>次生林(13.93 g/kg)>橡胶林(12.94 g/kg)>耕地(12.28 g/kg)>森林(12.23 g/kg)>休耕地(11.91 g/kg);50—80 cm土层中,土壤钾含量的排序为:竹林(19.04 g/kg)>次生林(15.11 g/kg)>休耕地(14.42 g/kg)>橡胶林(13.99 g/kg)>森林(13.74 g/kg)>耕地(13.49 g/kg);80—110 cm土层中,土壤钾含量的排序为:竹林(19.12 g/kg)>次生林(15.83 g/kg)>休耕地(15.41 g/kg)>森林(15.22 g/kg)>耕地(14.15 g/kg)>橡胶林(13.41 g/kg)。各土层中,次生林的钾含量仅次于竹林,这是因为土壤钾素的释放被灌木和杂草的根系固定在土壤中,且林下植被快速的养分归还能迅速补充土壤钾素。所有用地类型中的钾含量都随土层的加深而增加,且0—20 cm土层的钾含量都显著低于其他土层(p<0.05)。有研究证明,各土层中,土壤钾元素具有较强的淋溶性,钾含量与土壤黏粒含量呈显著正相关,本研究中,随土层加深,黏粒所占百分比在增大,土壤黏粒比表面大,含量高则吸附和维持的土壤钾元素就越高[19],所以钾含量随土层加深而增加。
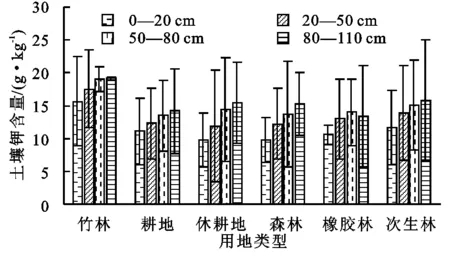
图6 不同土地利用方式下不同土层土壤钾含量
3.7 不同土地利用方式碳氮稳定同位素含量剖面分异
土壤的碳稳定同位素反映了当地的C3与C4植被的相对丰度[20],表征植被演替过程中土壤剖面有机碳的δ13C值的分布与植被类型的关系,氮同位素则反映了不同氮源的贡献,体现了氮源信息和地球化学过程的综合[21]。由表1可知,0—20 cm的土层中,森林的δ15N自然丰度显著低于竹林和耕地;20—50 cm的土层中,森林的δ15N自然丰度显著低于竹林和休耕地;其他土层中的碳氮同位素在不同土地利用类型下无显著差异。竹林中,碳氮同位素在各土层中无显著差异;在耕地和休耕地中,0—20 cm土层的碳氮同位素都显著低于其他3个土层深度的;森林和次生林的碳氮同位素都随着土层加深而增加,并且每层之间都具有显著性差异;橡胶林中,0—20 cm土层的δ15N自然丰度显著低于50—80 cm和80—110 cm,δ13C自然丰度在各土层中无显著差异。所有的δ13C自然丰度值都接近-27‰,已有研究表明:自然界C3植物碳同位素组成在-22‰~-34‰之间,C4植物的碳同位素组成在-9‰~-19‰之间[22]。此结果与该研究区主要为C3植物的植被类型吻合。枯枝落叶、化肥及有机肥的使用可能导致δ15N的丰富度明显改变[23]。大多数情况下,δ15N值不高,然而有时由于植被类型的改变和微生物活动会影响有机质转变,导致氮同位素丰度相应升高。0—20 cm土层中,竹林和耕地的δ15N显著高于森林的;20—50 cm土层中,竹林和休耕地的δ15N显著高于森林的。土壤有机质主要来源于地表植被的枯枝落叶,在排水良好的土壤中,随土层加深,土壤的碳氮同位素丰度会逐渐升高。但输入土壤中的有机物的植物类型是影响土壤碳氮同位素的主要因素,土壤内固有的碳氮分馏过程只会导致土壤剖面的δ13C和δ15N的变幅在1‰~3‰之间[24]。本研究中碳氮同位素变幅较小,与随土层加深黏粒含量增多有关系,因为随着土壤组成粒径的减小,与之结合的有机质腐殖化程度逐渐升高,因此土壤黏粒中有机质分解较为充分,从而产生较小的同位素分馏[25]。
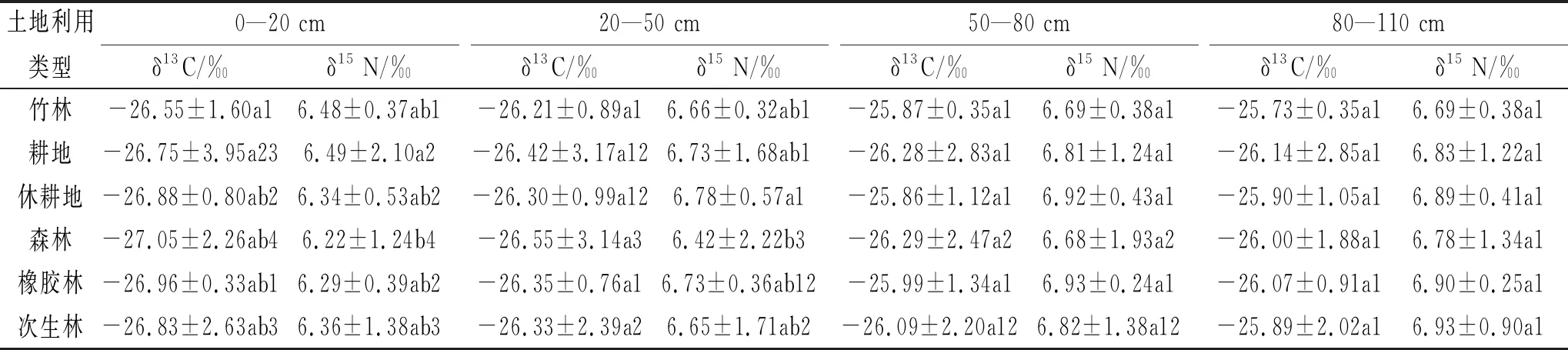
表1 不同土地利用类型和土层深度的碳氮稳定同位素丰度
注:表中数据为平均值±标准误,不同小写字母表示同一土层深度的同一土壤养分在不同土地利用类型之间差异在p<0.05水平显著,不同数字表示同一土地利用类型下的同一土壤养分在不同土层深度之间的差异在p<0.05水平显著。
3.8 不同土地利用方式土壤颗粒组成含量剖面分异
由表2可知,在0—20 cm的土层中,休耕地的砂粒所占百分比显著高于竹林的,休耕地和森林的粉粒和黏粒的颗粒所占百分比显著低于竹林的;20—50 cm的土层中,竹林的粉粒所占百分比显著高于休耕地、森林和橡胶林;在其他土层中,各用地类型下的土壤不同粒径颗粒所占百分比无显著差异。在竹林和橡胶林中,不同土层深度下的3种粒径的颗粒所占百分比无显著差异;在耕地和休耕地中,随着土层加深砂粒所占百分比在逐渐降低,黏粒所占百分比在逐渐升高;在森林和次生林中,砂粒所占百分比随土层深度加深而降低,黏粒所占百分比在逐渐升高,并且0—20 cm土层的黏粒所占百分比显著低于50—80 cm和80—110 cm土层。由此可见,竹林的土壤颗粒组成明显小于其他用地类型,这与研究所得竹林是各养分含量最高的用地类型相吻合,由于不同粒径的土壤颗粒对养分的吸附力和粘着性等存在明显差异,小粒径土壤颗粒中的养分相较于大粒径土壤颗粒的养分往往更难以矿化,不易被微生物所分解释放[26]。根据国际制土壤质地分级标准,耕地、休耕地、森林和橡胶林4种土地利用类型中除0—20 cm土层外土壤质地都属于壤质黏土,0—20 cm土层的土壤质地属于黏壤土,竹林和次生林的所有土层的土壤质地均属于壤质黏土。
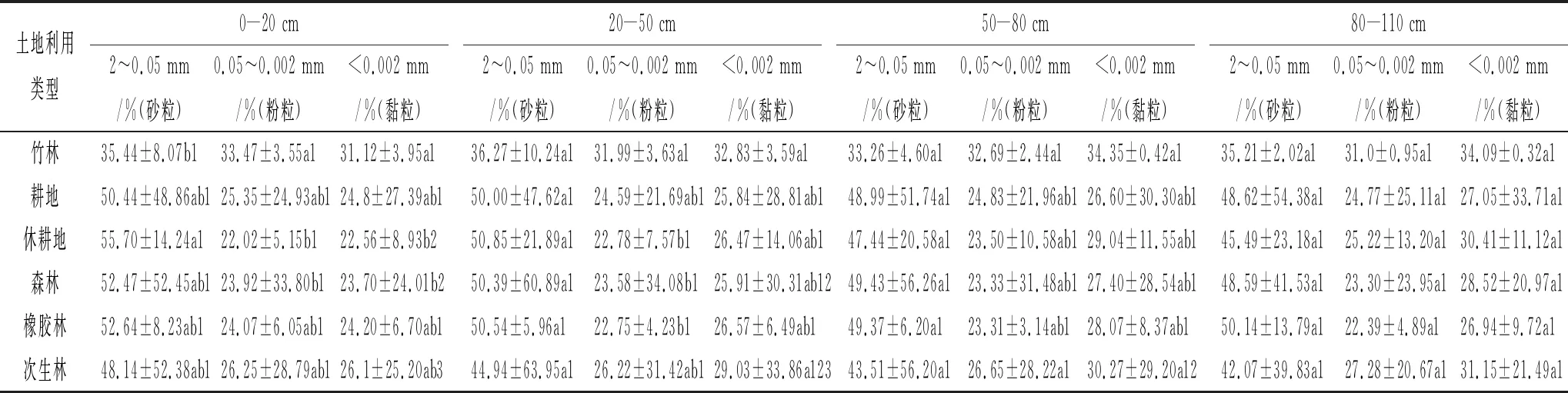
表2 不同土地利用类型和土层深度的原状积累土样的不同粒径土壤颗粒所占百分比
注:表中数据为平均值±标准误,不同小写字母表示同一土层深度的同一土壤养分在不同土地利用类型之间差异在p<0.05水平显著,不同数字表示同一土地利用类型下的同一土壤养分在不同土层深度之间的差异在p<0.05水平显著。
4 结 论
(1) 受热带地区高温高湿和土壤表层大量枯枝落叶的影响,不同土地利用方式下土壤pH均呈酸性。由于施肥的原因,导致耕地的pH高于其他用地类型。pH随土层变化不明显,因为随着土层深度的增加,植被类型或施肥对土壤酸度的影响越来越小。
(2) 在各土层中,土壤有机碳,总氮含量的大小顺序以及在不同土地利用方式之间的显著性差异较为一致。受植被保护和施肥耕作等人为因素影响,在各土层中,均是休耕地与橡胶林的有机碳、总氮含量较低,竹林是有机碳总氮含量最高的土地利用方式,耕地的有机碳和总氮含量在50—110 cm土层中较高。土壤有机碳总氮含量均随着土层自上而下逐层降低,表层土壤与底层土壤的有机碳总氮含量都达到显著性差异。
(3) 该地区属于热带山区受高温高湿的气候影响,导致有机质降解速率快,碳氮比值较低。耕地长期处于低温淹水状态,有机质分解缓慢故而积累,从而使碳氮比处于较高水平。竹林是磷钾含量最高的土地利用方式,这与有机碳总氮的情况相似。除休耕地和次生林外,其余4种土地利用方式下的磷含量基本上都随土层加深而减少,休耕地和次生林磷含量呈现出“U”形分布,各用地类型中钾含量均随土层加深而增加。
(4) 各土地利用方式下,δ13C的自然丰度在各土层中均无明显差异,δ13C的自然丰度都接近-27‰,与该研究区主要为C3植物的植被类型吻合。后续将对植物根系和凋落物的碳氮同位素进行分析,进而与土壤碳氮同位素进行比较,从而更好地反映土壤碳氮同位素的来源与转化过程。本研究中碳氮同位素变幅较小,与随土层加深黏粒含量增多有关系。竹林的土壤颗粒组成明显小于其他用地类型,这与研究所得竹林是各养分含量最高的用地类型相吻合。本研究针对不同粒径土壤的理化性质在不同土地利用方式下的剖面分布进行了探讨,之后的研究将致力于分别探究不同粒径大小的土壤颗粒中的理化指标,更加深刻的揭示土壤颗粒大小与土壤理化性质间的相关关系。
(5) 有研究证明,土地利用变化对土壤性质的影响在北纬20°~25°影响较大[27],可能的主要原因是本研究区山地多,坡度陡,并且降雨量大,雨热同季,加剧土壤有机质分解的同时,水土流失情况也比较严重,大大降低了土壤各养分含量,故该地区土壤较为贫瘠,与刘忠等人对老挝中部的土壤养分研究一致。研究证明,植被恢复有一定的土壤培肥作用,有必要进一步增强对土地的合理人为干扰,如耕作中加强水土保持措施和加大有机肥的施入,避免化肥的使用[28]。总之,要扩大森林面积,尤其是天然林面积,控制水土流失,恢复退化的休耕地,对单一种植模式的橡胶林进行合适物种的混种,在耕地中种植长久作物,实行轮作种植,把低产农田变成草地或森林,扩大造林,集约管理农田,实行农林复合、林草复合经营模式[29],才能实现老挝北部山区的可持续农业发展。
猜你喜欢
热带作物学报(2022年6期)2022-07-08 06:45:10
现代青年·精英版(2021年11期)2021-12-16 04:20:17
露天采矿技术(2020年6期)2021-01-05 00:37:54
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
浙江农业学报(2016年7期)2016-06-15 20:30:08
西南农业学报(2016年5期)2016-05-17 05:42:39
植物营养与肥料学报(2011年2期)2011-10-26 03:51:50
