
千岛湖片段化景观中蜘蛛群落物种多样性和功能群结构及其影响因素
2019-05-13 01:50周文婕黄杰灵谢央央罗媛媛
生态学报 2019年6期
周文婕,仲 磊,黄杰灵,谢央央,罗媛媛,*
1 中国计量大学生命科学学院,杭州 3100182 浙江大学生命科学学院,杭州 310058
生境片段化(或生境破碎化)是指大面积连续分布的生境变成空间上相对隔离的多个片段化小生境[1]。近几十年来,因人类活动的影响,生境片段化现象日益严重[2]。人为活动所造成的生境片段化是生物多样性丧失的主要原因之一[3]。生境片段化形成的片段的面积大小和隔离度等因素对生物多样性的影响成为当前保护生物学研究的一个热点[4-5]。
蜘蛛隶属于节肢动物门(Arthropoda)蛛形纲(Arachnida)蜘蛛目(Araneae),是一类种类繁多、数量巨大、分布广泛的捕食性生物,对抑制农林害虫发生、维持生态系统稳定等方面具有重要作用[6- 8]。蜘蛛和其他无脊椎动物类群相比,能被有效捕捉和鉴定[9- 11]。且由于蜘蛛对环境因子变化高度敏感,可以作为监测环境变化的指示类群,能够很好地反映环境变化过程及其对生物多样性的影响[12-13]。因此,蜘蛛被广泛地用作环境对生物多样性影响的指示生物[14-15]。
千岛湖位于浙江省淳安县境内,是1959年新安江水电站建设蓄水形成的人工湖泊,水面面积573 km2,岛屿面积409 km2,水位最高(108 m高程)时面积大于2500 m2的岛屿有1078个[16]。千岛湖库区内的岛屿和周边湖水形成的景观就是全球生境片段化的缩影,原有连续山脉的一座座山峰因水库大坝建设形成千余个岛屿(片段),组成了独一无二的片段化景观,故从生物遗传、个体、群落、生态系统和景观等各个层次,千岛湖库区是开展科学研究的理想场所,已成为全球生境片段化研究八个重要平台之一[17]。目前,已在千岛湖地区进行了不少研究,但主要是关于脊椎动物和植物的研究[18- 20],对节肢动物也进行了部分研究,多集中于昆虫,对蜘蛛的研究仅可见于Wu 等[21],该文研究的是蜘蛛的beta多样性,并且与本研究采取的取样方法不同,在功能群的划分上也有所不同。
本文选取千岛湖生境片段化景观中16个陆桥岛屿的蜘蛛群落作为研究对象,对不同岛屿上的蜘蛛多度以及种类分布做了调查研究,分析其物种多度、物种多样性、季节动态和功能群结构,并对不同大小岛屿上蜘蛛功能群组成的差异进行分析,从而探讨千岛湖片段化对蜘蛛群落物种多样性、功能群结构和季节变化等的影响,旨在丰富千岛湖库区景观生态学和生物地理学研究,为片段化景观中生物多样性保护对策的制定提供科学依据。
1 研究区域与研究方法
1.1 样岛选取
根据最新高清晰度卫星照片(或者航片资料)和淳安县开发总公司提供的千岛湖景观参数地形图,并结合千岛湖库区开展的相关工作[22-23]进行实地勘查,选取了16个不同面积和隔离度(距最近大陆的距离)的陆桥岛屿作为研究样岛,样岛编号沿用千岛湖植物群落调查使用的统一编号(图1)。
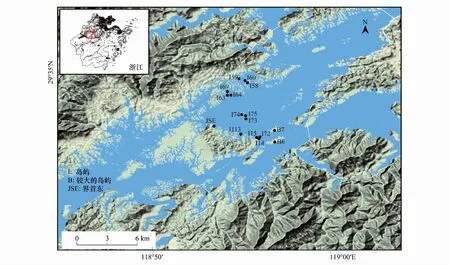
图1 千岛湖16个研究岛屿分布图Fig.1 Distribution of study islands in the Thousand Island Lake region
考察千岛湖库区的研究岛屿的实地状况,并参考Terborgh等[24]研究方法,按面积大小将这16个岛屿划分为2种类型:小岛(面积<1 hm2)11个,大岛(面积>1 hm2)5个。按照岛屿面积的大小确定采样样方个数,分别设立10个、15个、20个和25个样方,其中由于JSD岛屿面积大,故设立了30个样方(表1)。
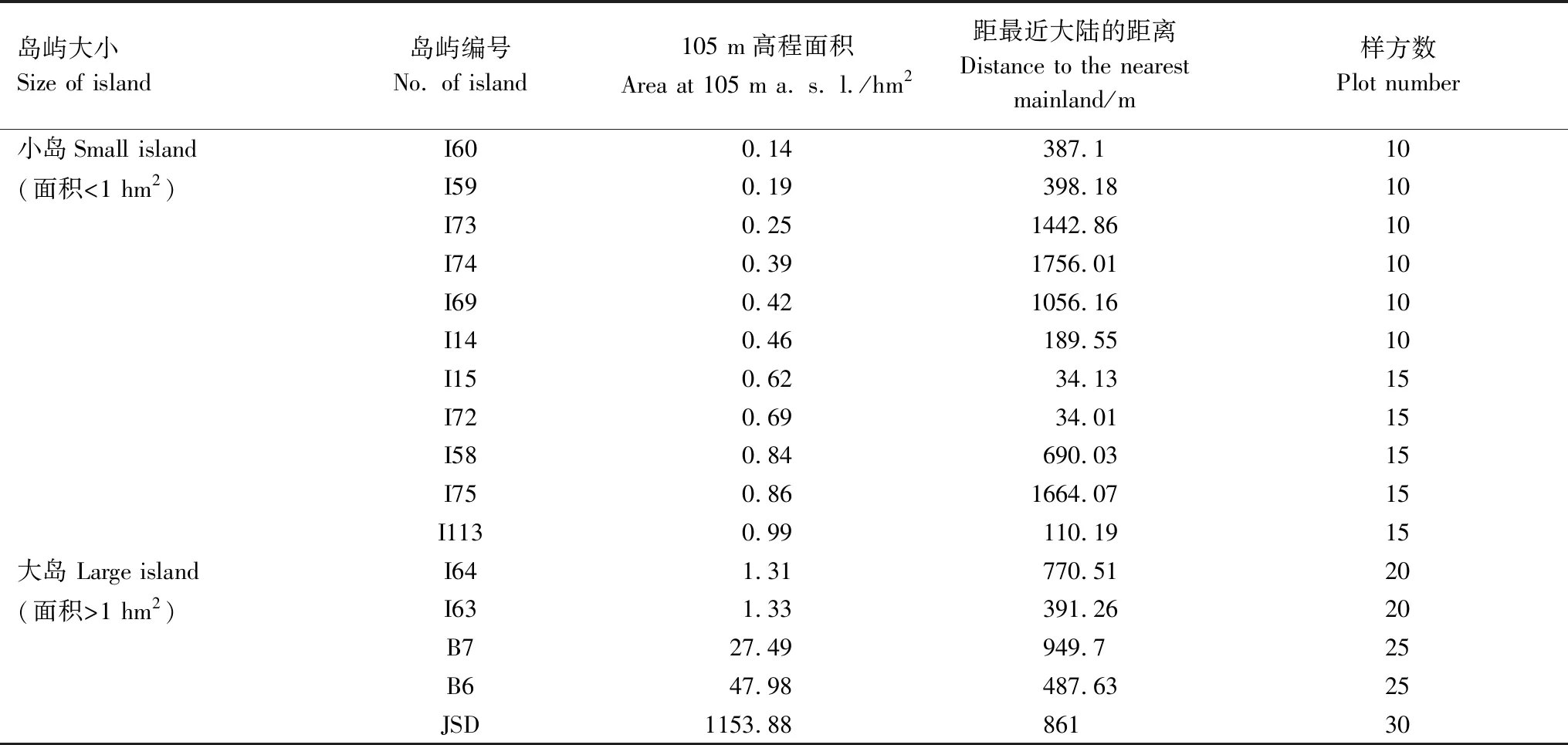
表1 千岛湖样岛的地理参数
表中的岛点符号请参看图1
1.2 取样和标本鉴定

图2 每个样方内的Winkler法剖面图Fig.2 The profile Winkler method in every plot
2010年春季至2011年秋季,春、夏、秋共六个季度均采用Winkler法[25]采集样岛上的蜘蛛,这是一种相对简便高效的方法,对生境的破坏不大,具有保持土壤完整,方便长期在野外进行调查土壤动物等优势[26]。相对于巴氏陷阱法[27],Winkler法取样时既可以取土表土样(可包括枯枝落叶),也可挖取深层土样,故Winkler法对生活在地表和土壤深层的土壤节肢动物均有较好的分离效率[28-29]。使用高为20 cm,面积为0.5 m×0.5 m的采集框,沿着土壤纵剖面挖取土样,厚度为2 cm左右,将挖取得到的土样和其表面的枯枝落叶一起全部放到密封袋里,用孔径为2 cm的筛网去掉密封袋中的土样内较大的石块,再把去掉石块剩下的土样等分成三袋,装入网筛孔径为2 mm的Winkler Sacks中,经过72 h的悬挂后,将布袋底部装有甘油和75%酒精混匀物的塑料杯取下,收集其中的动物样本,最后再把袋子里的土样重新倒回原来的地方(图2)。采集到的标本使用浓度为75%的酒精来保存。
分类鉴定时,将蜘蛛样本取出放到培养皿中,参考中国土壤动物检索图鉴[30]以及《中国蜘蛛原色图鉴》[31],分类鉴定到形态种。分类鉴定后统计每个样岛蜘蛛的数目与种类,比较分析物种多样性。
根据蜘蛛结网和捕食行为的特点,参考Höfer与Brescovit[32]和Sørensen[33]等的功能群分类方法,将从千岛湖16个样岛中所采集的蜘蛛分为五种类型:穴居型、结皿网型、结圆网型、伏击型和游猎型。

图3 16个样岛蜘蛛物种累积曲线Fig.3 Rarefaction curves of spider species on 16 study islands
1.3 数据处理
分别对各样岛6次收集的蜘蛛进行物种数及多度(个体数)的记录,并对2年所获得的蜘蛛进行优势种、常见种以及稀有种的分析,蜘蛛的优势等级以某一种多度占个体总数的百分比来划分[34]。
分析蜘蛛的α多样性,选用的α多样性指数为Shannon多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(J)[35]。采用单因素方差分析、线性回归等方法对不同样岛的功能群差异以及不同大小岛屿上蜘蛛的功能群的多度差异进行分析。
运用R语言对采样数据进行分析,计算物种累积曲线,验证采样充分性,利用物种累积曲线判断抽样量是否充分是根据曲线的特征来判断:如果曲线一直急剧上升,几为直线,表明抽样量不足,需要增加抽样量;如果曲线在急剧上升后变为渐近线,上升舒缓,则表明取样充分[36]。对不同的样岛的蜘蛛群落进行PCA排序,分析各岛屿群落相似性。
在以上分析中,均选用R 3.0.2的vegan 2.0-10 软件完成[37]并作图。
2 结果与分析
2.1 采样充分性分析
由图3可知,16个研究样岛中绝大部分的物种累积曲线在初期上升剧烈,后期相对趋于平缓,表明对相应样岛的蜘蛛群落采集较充分。另外,该曲线逐渐接近渐近线,说明如果继续增加取样,蜘蛛的种类数将达到最大值。
2.2 蜘蛛群落物种组成
2010年与2011年均分为春、夏、秋三季取样6次共获得蜘蛛30科82种1438头(附表),其中螲蟷科(Ctenizidaesp.2)在16个岛屿上均有分布。优势种为六眼幽灵蛛属(Spermophorasp.1)、管巢蛛科(Clubionidae sp.2)、螲蟷科 (Ctenizidae sp.2)、蟹蛛科(Thomisidae sp.3)分别占总数的7.37%、6.61%、5.84%、5.29%;常见种为节板蛛科(Heptathela sp.2)、螲蟷科(Ctenizidae sp.1、C.sp.3、C.sp.4)、跳蛛科(Salticidae sp.1、S.sp.4)、管巢蛛科(Clubionidae sp.1)、平腹蛛科(Gnaphosidae sp.1、G.sp.2、G.sp.3)、佐蛛科(Zoridae sp.1)、近管蛛科(Anyphaonidae sp.1)、六眼幽灵蛛属(Spermophorasp.3、S.sp.5)、卵形蛛科(Oonopidae sp.5)、栉足蛛科(Ctenidae sp.1)、逍遥蛛科(Philodromidae sp.1、P.sp.4、P.sp.6)、巨蟹蛛科(Sparassidae sp.3、S.sp.6)、蟹蛛科(Thomisidae sp.1)、栅蛛科(Hahniidae sp.1)分别占总数的2.02%、3.96%、2.36%、2.16%、3.48%、2.5%、3.48%、1.46%、1.81%、2.43%、1.04%、2.85%、4.45%、1.25%、1.32%、2.57%、2.57%、2.29%、3.62%、2.02%、2.09%、2.09%、3.69%,其他种的多度占总数的百分比均小于1.0%,为本地区的稀有类群。
以蜘蛛的物种及每个物种所对应的多度为属性对16个研究样岛进行PCA分析,结果(图4)表明:样岛I63、I58、I74、I75、B6、B7在图中分布较散,说明它们与其他样岛的相似性相对较低;其余岛屿分布较为密集,但并未重合,说明相似性较高但也有区别。
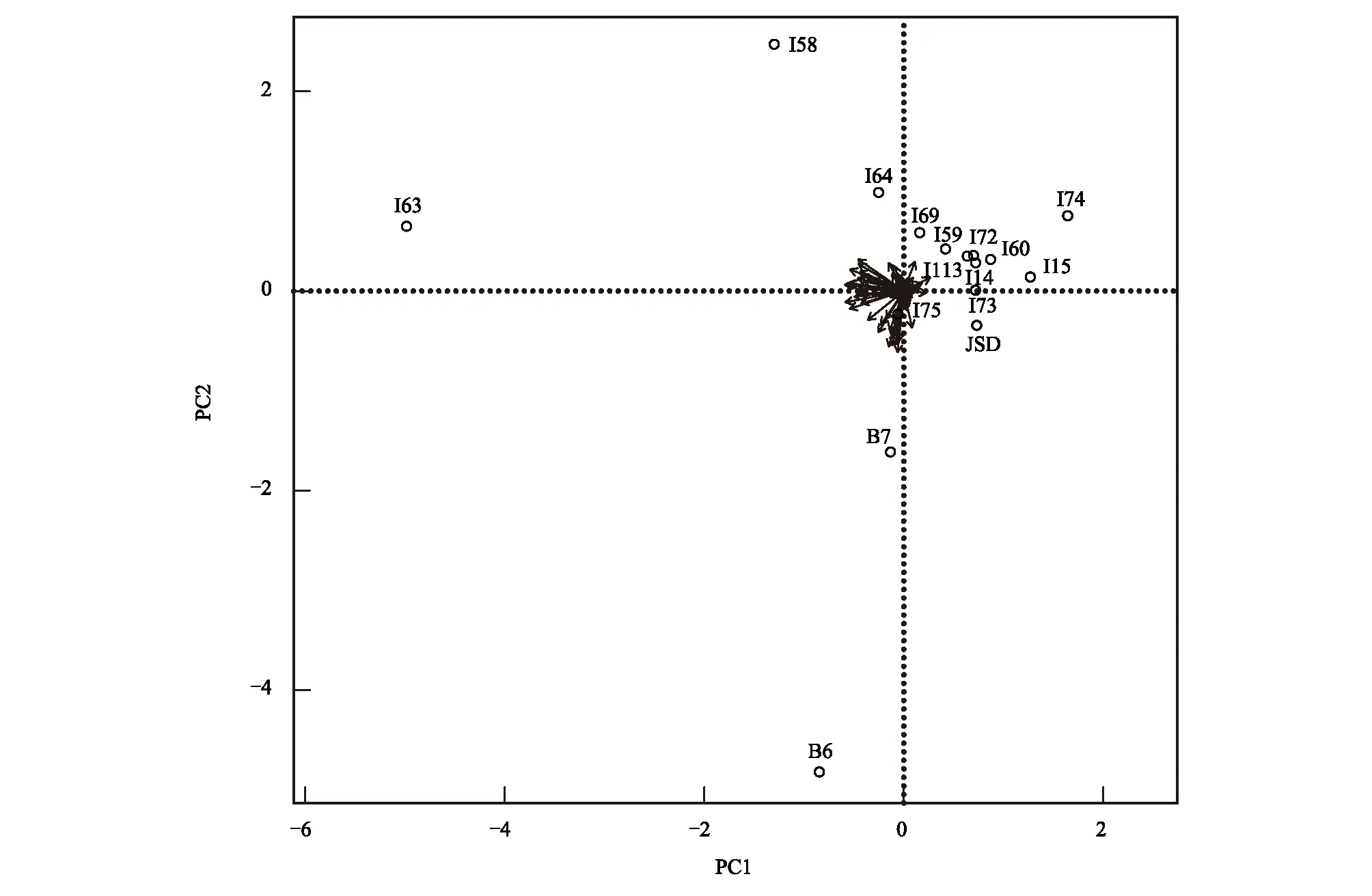
图4 82种蜘蛛与16个岛屿关系的PCA排序图Fig.4 Ordination diagram of the first two axes of principal component analysis for 82 spider species and 16 islands in the Thousand-Island Lake
2.3 蜘蛛的α多样性
本次研究中大岛与小岛间的α多样性无显著差异(P>0.05)。计算α多样性指数发现(表2),样岛I58与I75的Shannon多样性指数相对其他岛屿较高,同时这两个样岛的Simpson指数最高,样岛I113的Pielou均匀度指数最高;样岛I14的Shannon多样性指数、Simpson指数最低,样岛B6的Pielou均匀度指数最低。

表2 千岛湖各样岛蜘蛛的α多样性
*代表最小值;**代表最大值
2.4 蜘蛛物种多样性格局的季节动态
在6次取样中(表3),其中2010年夏季所获得的蜘蛛多度最少,2010年秋季所获得的蜘蛛的多度最多,不同季度采样所获得的蜘蛛在岛屿间不存在显著差异。

表3 千岛湖各样岛蜘蛛物种丰富度及多度分布
千岛湖16个岛屿样岛的蜘蛛物种丰富度秋季最高(图5),Shannon指数在春季与秋季间差异显著(P<0.05),夏季与秋季间差异显著(P<0.05);Simpson指数与Pielou指数在不同季度间差异不显著
2.5 功能群分析
蜘蛛功能群的组成上根据蜘蛛结网和捕食行为的特点,将从千岛湖16个样岛中所采集的蜘蛛分为五种类型:穴居型:节板蛛科、螲蟷科、弱蛛科、地蛛科、大疣蛛属;游猎型:拟平腹蛛科、跳蛛科、管巢蛛科、平腹蛛科、狼蛛科、猫蛛科、盗蛛科、光盔蛛科、佐蛛科、逸蛛科、近管蛛科;结皿网型:皿蛛科、幽灵蛛科、六眼幽灵蛛属、漏斗蛛科;伏击型:卵形蛛科、栉足蛛科、逍遥蛛科、巨蟹蛛科、蟹蛛科;结圆网型:园蛛科、球体蛛科、类石蛛科、栅蛛科、卷叶蛛科。
其中伏击型蜘蛛的多度最高,其次为游猎型蜘蛛,结圆网型蜘蛛多度最低,穴居型蜘蛛与其他各功能群呈显著差异(P<0.05);游猎型蜘蛛与伏击型蜘蛛的多度均显著高于结皿网型蜘蛛与结圆网型蜘蛛(P<0.05);其余功能群间不存在显著差异(图6)。

图5 千岛湖样岛蜘蛛物种丰富度的季节动态 Fig.5 Species abundances of spiders in different seasons in the Thousand-Island Lake region
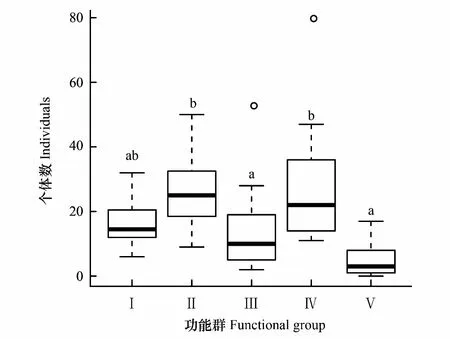
图6 不同蜘蛛功能群的多度差异 Fig.6 Individuals of various spider guilds across sampling islands

图7 千岛湖不同类型区域蜘蛛功能群比例 Fig.7 Proportion of various spider guilds across different type of sampling islands in Thousand-Island Lake region
蜘蛛的功能群组成和数量在小岛与大岛这两种不同的岛屿类型上表现出显著差异。小岛上伏击型蜘蛛的物种数显著高于大岛上的物种数(P<0.05);其余功能群在不同类型的岛屿间不存在显著差异。比例上,结皿网型蜘蛛和伏击型蜘蛛在小岛上占功能群总体数目的比例显著低于大岛上的比例(P<0.05),小岛上的游猎型蜘蛛占功能群总体数目的比例显著高于大岛上的比例(P<0.05),结圆网型蜘蛛与穴居型蜘蛛在不同类型的岛屿间不存在显著差异(图7)。
3 讨论
本次调查共设样岛16个,历时两年6次取样,即除了冬天,其余每个季节均进行了两次采样。共获得蜘蛛标本3503头,用于统计分析的成蛛1438头,归属于30科82种。
由物种累积曲线可知,本次调查中蜘蛛采样基本充分。蜘蛛的物种数随采样次数的增加而呈递增趋势,但具体到各个样岛又有所区别,大部分岛屿蜘蛛的物种数均呈现较为平缓的上升趋势。
目前蜘蛛的相关研究表明群落的结构组成、种群的数量变动与环境因子的关系密切[38- 40]。2002年于晓东等[41]对东灵山地区地表甲虫的研究,指出植被类型和人为干扰程度是决定该地区物种分布和数量组成的关键因素。本研究所选取的样岛形成时间短,面积尺度小,岛屿集中,植被类型较相近,几乎均为马尾松,生境较一致[42-43],而Tsai等[44]2006年曾报道在亚洲热带岛屿,对天然林、耕作林地、次生林和草地中蜘蛛多样性比较的研究,结果表明严重受干扰的耕作林地中蜘蛛多样性比其他生境高。因而,人为干扰少时,蜘蛛群落的个体数量很容易受到环境变化的影响,更利于各类群蜘蛛栖息[45]。
在物种的组成上,螲蟷科(Ctenizidae sp.2)在16个岛屿上均有分布,16个样岛的植被类型均为马尾松林。优势种为六眼幽灵蛛属(Spermophorasp.1)、管巢蛛科(Clubionidae sp.2)、螲蟷科(Ctenizidae sp.2)、蟹蛛科(Thomisidae sp.3),说明这四种蜘蛛更适应人为干扰少的环境,这与郑国等研究指出的幽灵珠科更喜欢生境复杂,人为干扰较少的环境[46]相符合。而采用巴氏陷阱法采集的千岛湖31个样岛所获得的蜘蛛的优势种分别为狼蛛科、平腹蛛科、光盔蛛科、蟹蛛科和异纺蛛科[47]。差异的产生除受气候和环境等因素影响外,采集方法的不同可能也是一个重要原因[46]。PCA分析表明,部分岛屿(I63、I58、I74、I75、B7)在图中分布较散,物种组成相似性较低,其余岛屿分布较为密集,但并未重合,说明相似性较高但已有区别。
本次调查中中大岛与小岛间的α多样性无显著差异。蜘蛛的功能群结构分析显示,穴居型的螲蟷科、游猎型的管巢蛛科、结皿网型的六眼幽灵蛛属、伏击型的蟹蛛科为优势种。其中伏击型蜘蛛的多度最高,其次为游猎型蜘蛛,结圆网型蜘蛛多度最低,穴居型蜘蛛与其他各功能群呈显著差异;游猎型蜘蛛与伏击型蜘蛛的多度均显著高于结皿网型蜘蛛与结圆网型蜘蛛;结皿网型蜘蛛和伏击型蜘蛛在小岛上占功能群总体数目的比例显著低于大岛上的比例,小岛上的游猎型蜘蛛占功能群总体数目的比例显著高于大岛上的比例;伏击型蜘蛛的物种数在小岛与大岛间存在显著差异(P<0.05)。伏击型蜘蛛物种丰富度存在显著差异的可能原因是天然林落叶层较厚,适合于伏击性蜘蛛躲避和捕食[48]。在2007年喻国辉等[49]对2种不同采样方法对麦田蜘蛛群落结构的比较研究,结果显示相同年份不同采样方法获得的蜘蛛群落组成不同,如陷阱法主要采集地表游猎型蜘蛛、地面穴居型蜘蛛和少数伏击性蜘蛛,单位面积法主要采集落叶层结微网型蜘蛛和少数地表游猎型蜘蛛、地面穴居型蜘蛛,扫网法主要采集冠层蜘蛛和灌木层结网型蜘蛛等[48]。
蜘蛛群落对季节性降雨和气温变化较敏感[50-51]。这是由于不同的蜘蛛对温湿度喜好不同,而能改变小生境的气候因子对蜘蛛群落影响较大[52-53]。Wagner等[54]还认为蜘蛛的季节性变化可能与春秋季蜘蛛的行为和繁殖特性不同有关。本研究中,Shannon-Wiener指数在不同季节间差异显著,秋季物种丰富度最高,反映了季节性降雨和气温变化对它的深刻影响。因而,生境片段化对蜘蛛功能群的分布产生了一定影响。
综上所述,千岛湖库区的生境片段化对蜘蛛的物种及功能群组成都已经产生了一定的影响,蜘蛛的部分功能群在小岛与大岛间表现出显著差异,Shannon-Wiener指数在不同季节间差异显著,秋季物种丰富度最高。
致谢:浙江省淳安县新安江开发总公司和淳安县林业局为野外调查提供帮助,黄杰灵、谢央央同学野外采样,谢央央同学做了物种的分类与鉴定,浙江大学千岛湖科学研究基地为野外调查提供帮助,特此致谢。
附表:
附表 蜘蛛物种组成和多度
Attached table Species composition and abundance of spiders
种名Species name总多度Total abundance出现的岛屿数Number of islands presented相对多度Relative abundance/%漏斗蛛科 AgelenidaeAgelenidae sp. 1860.56近管蛛科AnyphaonidaeAnyphaonidae sp. 141122.85Anyphaonidae sp. 2960.63园蛛科AraneidaeAraneidae sp. 1210.14Araneidae sp. 2740.49Araneidae sp. 3110.07地蛛科AtypidaeAtypus sp. 1940.63Atypus sp. 2750.49管巢蛛科ClubionidaeClubionidae sp. 150153.48Clubionidae sp. 295146.61栉足蛛科 CtenidaeCtenidae sp. 137132.57螲蟷科CtenizidaeCtenizidae sp. 157143.96Ctenizidae sp. 284165.84Ctenizida sp. 334122.36Ctenizidae sp. 431102.16卷叶蛛科DictynidaeDictynidae sp. 1520.35Dictynidae sp. 2110.07Dictynidae sp. 3110.07平腹蛛科GnaphosidaeGnaphosidae sp. 12181.46Gnaphosidae sp. 226131.81Gnaphosidae sp. 335102.43Gnaphosidae sp. 41260.83栅蛛科HahniidaeHahniidae sp. 153123.69Hahniidae sp. 2540.35节板蛛科LiphistiidaeHeptathela sp. 1430.28Heptathela sp. 229112.02弱蛛科LeptonetidaeLeptoneta sp. 1110.07Leptoneta sp. 2110.07Leptoneta sp. 3110.07Leptoneta sp. 4210.14皿蛛科LinyphiidaeLinyphiidae sp. 1730.49Linyphiidae sp. 2320.21光盔蛛科LiocranidaeLiocranidae sp. 1970.63Liocranidae sp. 21040.7狼蛛科 LycosidaeLycosidae sp. 1750.49大疣蛛属MacrotheleMacrothele sp. 1110.07卵形蛛科OonopidaeOonopidae sp. 1440.28Oonopidae sp. 2330.21Oonopidae sp. 31260.83Oonopidae sp. 41050.7Oonopidae sp. 519111.32猫蛛科 OxyopidaeOxyopidae sp. 1110.07逍遥蛛科PhilodromidaePhilodromidae sp. 137122.57Philodromidae sp. 2540.35Philodromidae sp. 31260.83Philodromidae sp. 433132.29Philodromidae sp. 5850.56Philodromidae sp. 652153.62幽灵蛛科PholcidaePholcidae sp. 1110.07Pholcidae sp. 2330.21盗蛛科 PisauridaePisauridae sp. 1220.14跳蛛科SalticidaeSalticidae sp. 150153.48Salticidae sp. 2860.56Salticidae sp. 3540.35Salticidae sp. 436132.5
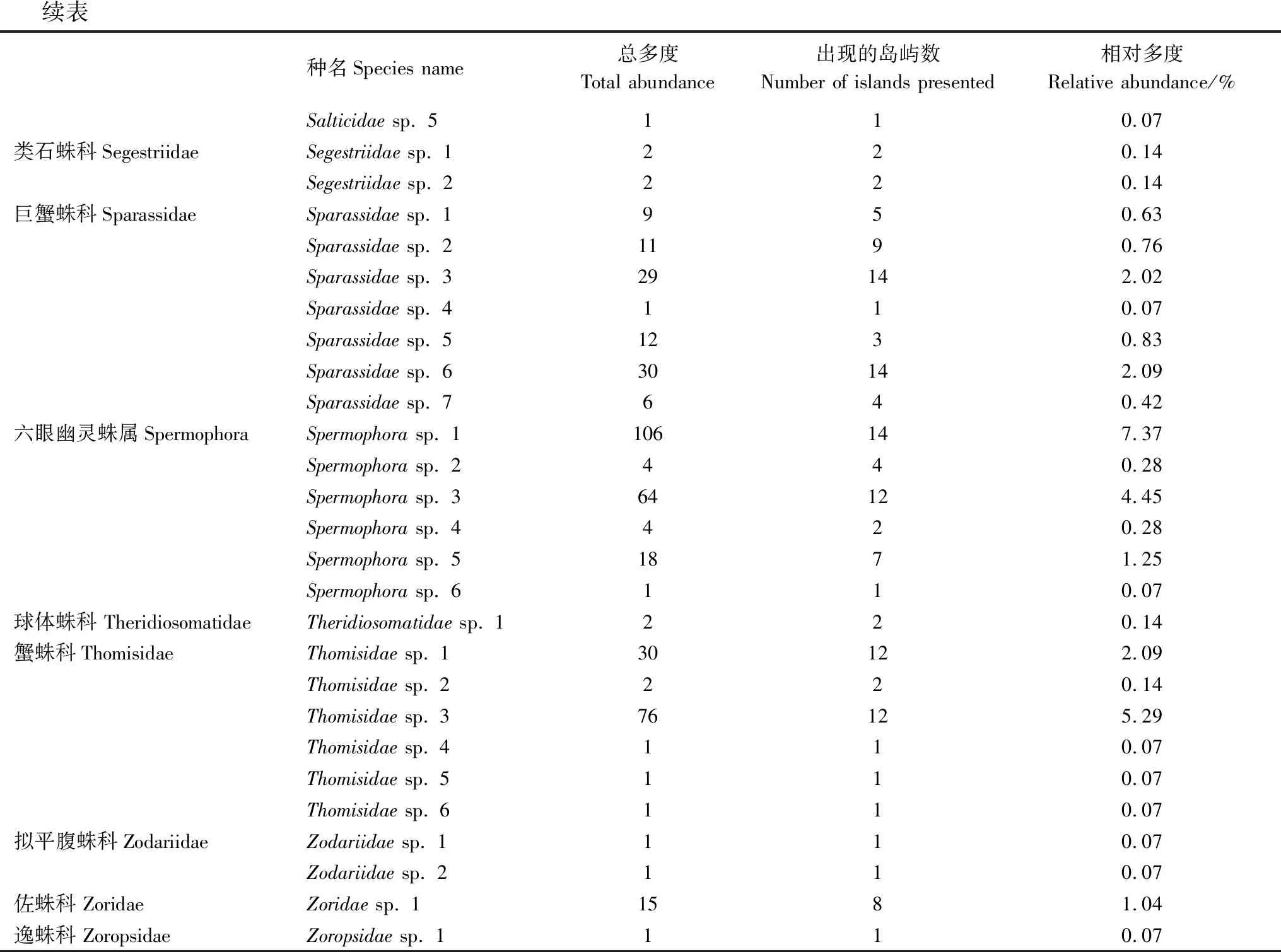
续表种名Species name总多度Total abundance出现的岛屿数Number of islands presented相对多度Relative abundance/%Salticidae sp. 5110.07类石蛛科SegestriidaeSegestriidae sp. 1220.14Segestriidae sp. 2220.14巨蟹蛛科SparassidaeSparassidae sp. 1950.63Sparassidae sp. 21190.76Sparassidae sp. 329142.02Sparassidae sp. 4110.07Sparassidae sp. 51230.83Sparassidae sp. 630142.09Sparassidae sp. 7640.42六眼幽灵蛛属SpermophoraSpermophora sp. 1106147.37Spermophora sp. 2440.28Spermophora sp. 364124.45Spermophora sp. 4420.28Spermophora sp. 51871.25Spermophora sp. 6110.07球体蛛科 TheridiosomatidaeTheridiosomatidae sp. 1220.14蟹蛛科ThomisidaeThomisidae sp. 130122.09Thomisidae sp. 2220.14Thomisidae sp. 376125.29Thomisidae sp. 4110.07Thomisidae sp. 5110.07Thomisidae sp. 6110.07拟平腹蛛科ZodariidaeZodariidae sp. 1110.07Zodariidae sp. 2110.07佐蛛科 ZoridaeZoridae sp. 11581.04逸蛛科 ZoropsidaeZoropsidae sp. 1110.07
猜你喜欢
人文天下(2022年7期)2022-09-15
思维与智慧·上半月(2022年4期)2022-04-08
落叶果树(2021年6期)2021-02-12
四川文学(2020年10期)2020-02-06
——千岛湖站
法人(2019年1期)2019-01-10
现代园艺(2018年1期)2018-03-15
中国自行车(2017年1期)2017-04-16
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
儿童故事画报(2015年5期)2015-07-22
