
垂直方向磷素竞争对杉木根系生长及生物量分配的影响
2019-05-13 01:50许静静吴文景RashidMuhammadHaroon马祥庆吴鹏飞
生态学报 2019年6期
许静静,李 琦,吴文景,Rashid Muhammad Haroon U,马祥庆,吴鹏飞,*
1 福建农林大学林学院,福州 3500022 国家林业局杉木工程技术研究中心,福州 3500023 驻马店水土保持监测中心,驻马店 463000
磷作为植物生长发育过程中所必须的矿质元素,常以多种方式参与植物代谢过程[1]。但我国大部分农林用地严重缺磷,这极易限制植物正常生长[2]。加之,磷在土壤中移动速率慢,多集中于土壤表层[3],导致大部分植物根系分布于土壤表层,从而加剧了邻株根系之间对有限磷素资源的竞争,这种竞争甚至比地上部分对光的竞争更为激烈[4-7]。然而,植物根系可通过改变其空间分布,迅速占领养分竞争区域,或退出竞争区域,以维持自身正常生长[8]。罗维成等[9]研究确证,当有竞争植物存在时,疏叶骆驼刺(Alhagisparsifolia)与竞争植物的根系会优先占有靠近它们“共有”空间这一侧,即便是自己另一侧与竞争植物拥有同样大小的空间;但苦豆子(SophoraAlopecuroides)会诱导根系向具有丰富养分资源的区域生长,以避免邻株根系对有限资源的激烈竞争[10]。还有植物可通过根系错位来避免同种基因型植物的竞争[11]。邻株竞争引起的这种根系空间重叠或错位分布的应对策略对植物生长具有重要意义[12-14]。因此,有关邻株植物根系对地下共有资源的竞争已成为当前研究热点。
杉木(Cunninghamialanceolata(Lamb.) Hook)作为我国南方重要的速生造林树种,长期以来多代连栽人工林栽培制度及南方红壤有效磷匮乏现象已导致杉木人工林产量下降日趋明显[15]。马祥庆等[16-20]突破传统上利用施肥提高杉木林生产力的营林措施,从杉木高效率利用磷素的角度出发,筛选出了一系列高效利用土壤磷的杉木家系和无性系,同时,初步揭示出在低磷胁迫环境中,磷素高效利用杉木基因型的形态生理学适应机制。特别地,有些杉木基因型可通过根系的大量增生以寻觅更多有效性的磷源来满足植株生长需求[21]。邹显花等[21]在研究杉木根系对不同磷斑块浓度觅磷效率中发现,在表层低磷情况下通过根系的增生提高觅磷效率。韦如萍等[22]也发现缺磷胁迫下杉木根系表面积和根尖数量明显增多。然而,这些研究结果多以杉木单株为研究对象,未能考虑实际林分中杉木邻株之间对共有资源的竞争情况,特别是在杉木人工林首次抚育间伐(9—10年生)之前,邻株杉木根系之间难免发生竞争现象。李琦等[23]研究报道:杉木地径和根长等根系形态指标在不同竞争强度和供磷水平中均明显增加。当邻株杉木根系在水平方向生长空间充裕,但垂直方向空间受限的情况下,低磷处理促进了根长度、体积、表面积、平均直径的增生,以有利于根系在水平生长空间内的觅磷效率[20]。那么,在土壤表层非常拥挤的环境下,根系是否会改变其生长趋向性,向下觅磷生长而在垂直方向引起竞争呢?
鉴于此,本文拟通过设置在一个在水平方向上极度拥挤的空间中,分析杉木在垂直方向磷素竞争对邻株杉木幼苗根系生长的影响。试验以同一杉木无性系幼苗为研究对象,设计竞争处理和3个供磷水平的室内沙培模拟试验,采用破坏性的方式收获幼苗,分别在苗木种植的前期(50 d)、中期(100 d)和后期(150 d)测定不同竞争与不同供磷水平条件下杉木根系的生物量、根冠比、比根长、根表面积、根直径、根体积等指标的变化规律,旨在揭示出低磷环境中杉木根系对土壤表层生长空间极度拥挤的响应策略,为提高南方红壤林区杉木林生产力提供科学依据。
1 材料和方法
1.1 试验材料
研究对象选择福建省洋口国有林场培育的杉木无性系“洋020”,该无性系具有侧枝短、冠幅窄、树冠稀松、生长快、材质好等特点,适合密植的造林方式,目前造林初始密度3600—3900株/hm2[24]。参与试验材料为18个月大、高度(19.8±0.4)cm、地径(3.5±0.1)mm,且生长健壮、长势均一的杉木幼苗。
1.2 试验设计

图1 邻株竞争处理盆栽示意图Fig.1 Experimental design to study adjacent plants competition
于2013年7月—12月在福建农林大学植物园温室大棚,采用长10 cm、宽10 cm、高40 cm的自制玻璃装置进行沙培盆栽试验(图1)。为构建邻株竞争环境试验,以每盆栽植2株根基距离2.5 cm(平均1/2冠幅的宽度)的杉木幼苗,并以单株栽植的幼苗为对照组,即无竞争处理。选用洗净河沙(总磷含量(0.11±0.004)mg/kg,有效磷为痕量)为培养基质,每盆沙子的重量为:4.6 kg,并设计3个不同的供磷水平:不供磷处理(P0,0 mg/kg KH2PO4)、低磷处理(P6,6 mg/kg KH2PO4)和正常供磷处理(P12,12 mg/kg KH2PO4),每个处理组12盆。选用特定的吸水树脂(water-absorbent resin,该树脂具有不含磷素、吸附性强、缓慢释放养分、植物根系可穿透等特点[25]),在吸附KH2PO4之后与洗净的河沙混合均匀后用于盆栽试验,以尽量模拟自然土壤环境中磷素的分布状况,从而突破传统试验中不断从培养基质表层供磷的施磷方法,有效避免了外施磷素在杉木幼苗生长基质表层不断累积的现象。为保证试验期间杉木幼苗的正常生长,每隔3 d浇一次不含磷素的Hoagland营养液[26],每盆每次浇50 mL。为平衡不同供磷处理间K+含量的差异,选择用KCl来替代。每天傍晚浇纯水50 mL。
采取破坏性试验方式进行参试苗木的收获,即分别在试验盆栽50 d、100 d、150 d后,每次收获4盆。苗木进行收获时,用水不断冲洗杉木根系沙土,加速其从盆栽容器底部圆孔流走,以保持幼苗根系的完整性。待根系和培养基质完全分离后,进行杉木幼苗根系取样及形态生长指标的测定。
1.3 测定方法
1.3.1根系形态参数测定
分别在试验盆栽前、盆栽后第50 d、100 d和150 d,利用根系图像分析系统软件WinRHIZOTM(version4.0b,Regent Instruments Inc.,Canada)进行杉木幼苗根系总长度、总表面积、平均直径和总体积等形态指标的测定。
1.3.2生物量测定
将收获后的杉木幼苗分为根系和地上部分,经108℃杀青30 min,再由80℃烘干至恒重,分别称量测定每株各部分干物质质量[27]。分别计算根冠比及比根长(specific root length),其中比根长作为单位质量根系的长度,可以表征根系生理与形态功能的一个重要指标[28]。
1.4 数据统计
苗木根系生长指标采用其增量,即收获时与盆栽前根系总长度、总表面积、平均直径、总体积的差值进行分析,邻株栽植数据表示同一测定指标的平均值。利用 SPSS(19.0)进行竞争处理和供磷水平双因素方差分析(Two-way ANOVA)。若两个因素无显著交互作用(P>0.05),则进行单个因素的方差分析,利用 LSD 多重比较方法进行检验(P=0.05)。采用Origin Pro 8.5软件将所有数据结果以平均值±标准误差表示,* 和不同字母分别代表不同处理间有显著差异(P<0.05)。
2 结果与分析
2.1 垂直方向磷素竞争对邻株杉木根系生物量的影响
3个不同试验阶段中,竞争处理和供磷水平对杉木幼苗地上部生物量、根系生物量和根冠比的影响均不存在显著的交互作用(P>0.05,表1)。从单个因素上来看,试验初期(50 d)时,竞争处理对3个生物量指标的影响均未达显著水平,表现为杉木地上部生物量与非竞争处理的单株幼苗地上部生物量基本持平,而根系生物量、根冠比均低于非竞争处理的单株幼苗(图2中50 d所示);至试验末期(150 d)时,竞争处理对根系生物量和根冠比的影响达显著水平(P<0.05),竞争与非竞争条件下生物量指标均增加,且呈上升趋势(图2),地上部生物量随胁迫时间的延长未达显著水平,但呈上升趋势(图2)。供磷水平对杉木幼苗地上部生物量、根系生物量、根冠比的影响均未达显著水平(表1)。在胁迫时间的影响下,地上部生物量、根系生物量和根冠比仍呈增加趋势(图3),其中地上部生物量随着供磷水平的增加呈下降的趋势(图3)。根系生物量和根冠比则在前期和中期随着供磷量增加而下降,而在后期上升,且增量远多于前期和中期(图3)。
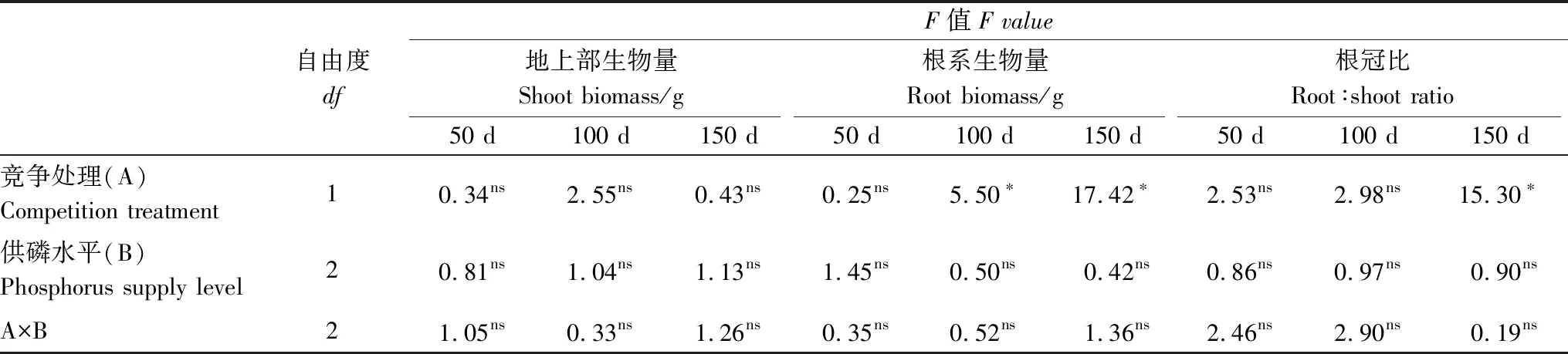
表1 不同试验时期竞争处理和供磷水平对杉木幼苗生物量的影响情况分析
*,P< 0.05; ns,P≥ 0.05; 50 d、100 d和150 d表示试验过程中分别在3个收获阶段所测定的生物量指标

图2 竞争处理对杉木幼苗生物量的分配Fig.2 Biomass distribution pattern of Chinese fir seedlings during competitive treatments* 表示竞争处理与对照(非竞争处理)之间的差异达显著水平(P<0.05)

图3 不同供磷水平对杉木幼苗生物量的分配Fig.3 Biomass distribution pattern of Chinese fir seedlings under different phosphorous supply levelsP0:不供磷处理,no phosphorus treatment;P6:低磷处理,low phosphorus treatment;P12:正常供磷处理,normal phosphorus treatment
2.2 垂直方向磷素竞争对杉木幼苗根系形态生长的影响
双因素方差分析结果(表2)表明,竞争处理和供磷水平对杉木幼苗根长增量、根平均直径增量指标的影响存在明显的交互作用(P>0.05),而对根表面积增量、根体积增量的影响存在不明显的交互作用(P>0.05),竞争处理对根系形态指标的影响均达显著水平,但供磷水平对根系形态指标的影响部分达显著水平(表2)。竞争处理中杉木根系形态增量均明显高于非竞争处理的单株幼苗,且随着胁迫时间的增加,根系形态增量均表现为显著上升趋势,其中在胁迫中期和后期的增量明显高于前期(图4,图5)。随着供磷水平的提高,根表面积和根体积增量大体上呈先上升后下降的趋势(图5),而根长和根平均直径增量分别在中期和后期也才呈现此趋势(图4)。
总的来说,竞争处理下的杉木幼苗根长、根表面积、根平均直径、根体积增量远大于对照处理下幼苗的增量,随着胁迫时间的延长表现为显著上升趋势。4个形态指标中均呈现出随着供磷水平的增加,大体呈现先上升后下降的趋势;随着胁迫时间的延长表现为显著的上升趋势。

表2 竞争处理和供磷水平对杉木幼苗根系形态生长的影响
*,P<0.05; ns,P≥0.05。50 d、100 d和150 d表示试验过程中在3个收获阶段所测定的根系形态生长指标
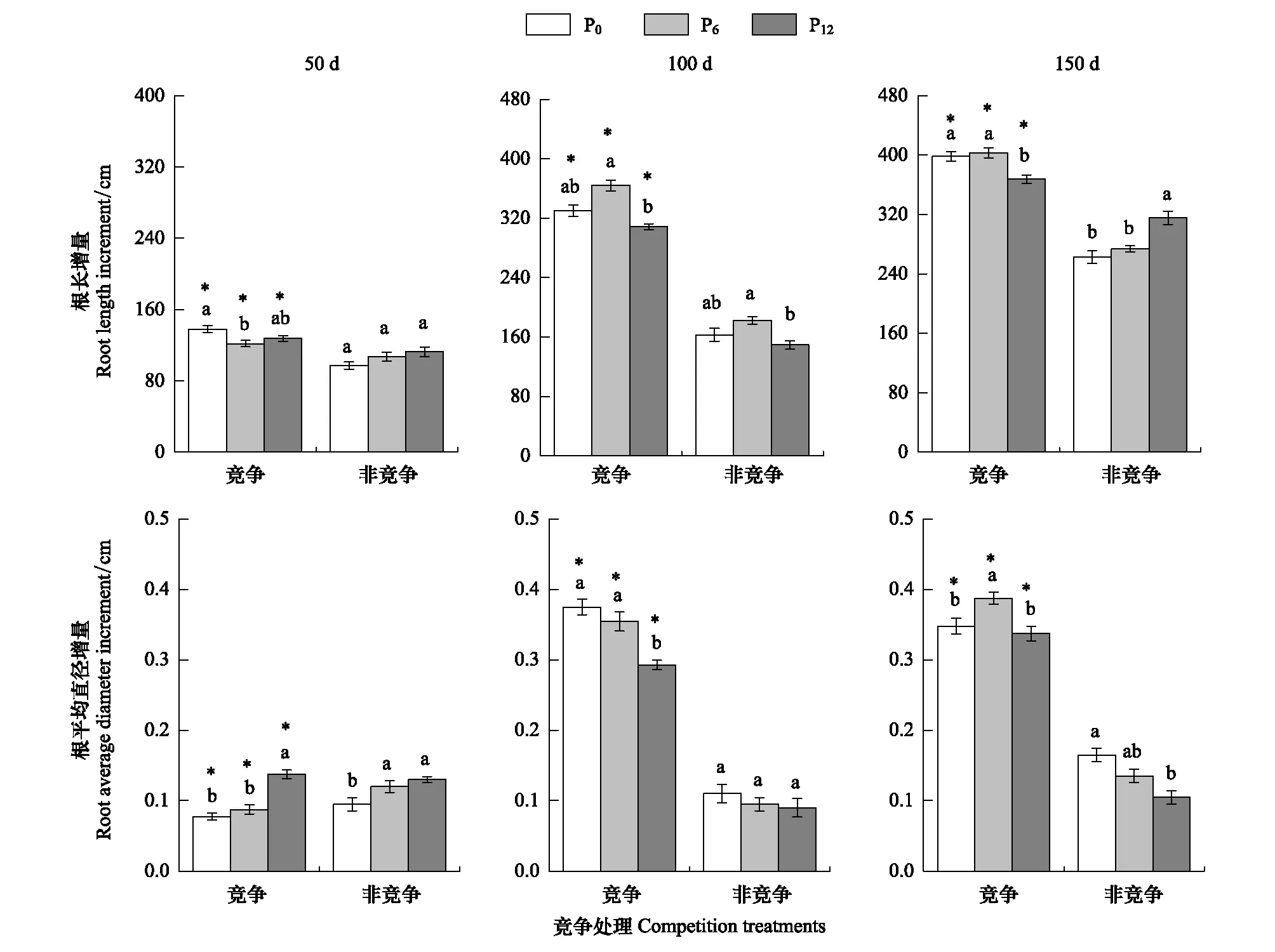
图4 竞争处理和供磷水平下杉木幼苗根系长度与平均直径的增量Fig.4 Increments of root length and average diameter respectively of Chinese fir seedlings under competition treatments and phosphorous supply levels*表示竞争处理与对照(非竞争处理)之间的差异达显著水平(P<0.05);不同小写字母表示不同供磷水平差异显著(P<0.05)
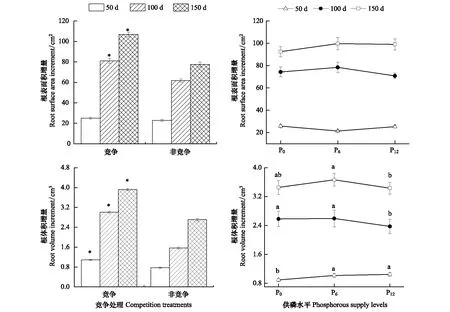
图5 竞争处理和供磷水平下杉木幼苗根系形态的增量Fig.5 Increments of root surface area and volume respectively of Chinese fir seedlings under competition treatments and phosphorous supply levels* 表示竞争处理与对照(非竞争处理)之间的差异达显著水平(P<0.05);不同小写字母表示不同供磷水平差异显著(P<0.05)
2.3 垂直方向磷素竞争对杉木幼苗比根长的影响
竞争处理和供磷水平对杉木幼苗比根长的影响不存在明显的交互作用(P>0.05),从单因素方差分析可以看出,竞争处理对比根长的影响均达到差异显著(P<0.05,表3)。在竞争处理中,比根长明显高于非竞争处理单株幼苗,且随着胁迫时间的延长,竞争处理和对照组的幼苗出现先升后降的趋势(图6)。而供磷水平对杉木幼苗比根长的影响不显著(P>0.05),整体随着供磷水平的增加呈现先升后降的趋势,随着胁迫时间的延长,比根长中期和后期明显高于前期增量(图6)。
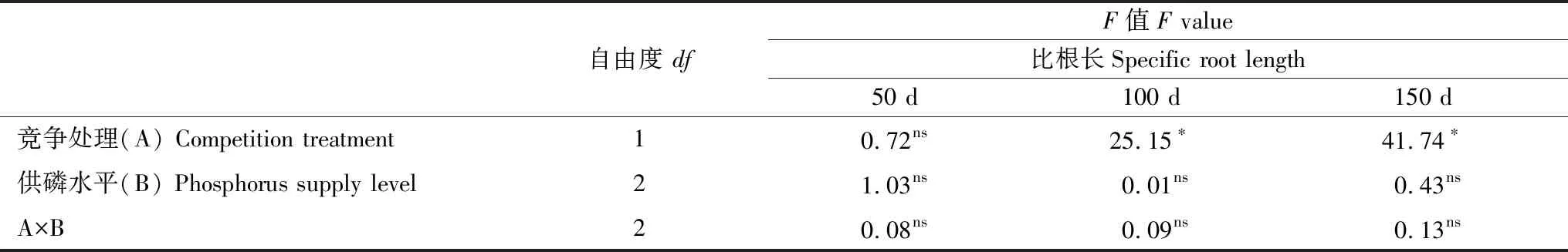
表3 不同试验时期竞争处理与供磷水平对杉木幼苗比根长的影响情况分析
*,P< 0.05; ns,P≥ 0.05。50 d、100 d和150 d表示试验过程中在3个收获阶段所测定的生物量指标
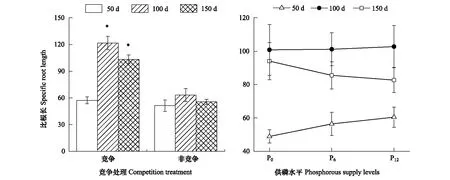
图6 竞争处理和供磷水平下杉木幼苗比根长的增量Fig.6 The increment of specific root length of Chinese fir seedlings under the competition treatments and phosphorous supply levels* 表示竞争处理与对照(非竞争处理)之间的差异达显著水平(P < 0.05);不同小写字母表示不同供磷水平差异显著(P < 0.05)
3 讨论
竞争是植物群落中普遍存在的现象,在植物生长发育过程中,与相邻个体会不断竞争水分、光照和养分[29]。尤其当植物根系遭受邻株竞争时,会通过调整根系生长趋向及空间分布格局,包括增加或减少根系长度[30-31],以主动寻找非竞争区域的资源[8],或有意避开竞争区域。许华森等[14]在对大豆(Glycinemax)和核桃(JuglansRegia)的研究中发现,其根系在垂直和水平方向上的竞争强度有所差别。Novoplansky等[32]通过建立竞争模型认为根系空间结构的差异性使植物吸收不同深度的养分。本文通过模拟根系在水平生长空间受限的条件下,研究发现:邻株竞争处理和供磷水平对杉木幼苗地上部生物量、根系生物量和根冠比的影响均不存在显著的交互作用。但从单个因素上来看,竞争处理对杉木地上部生物量的影响未达显著水平(图2),但根系生物量与根冠比均明显低于对照非竞争处理(图2),这与马雪红等[33]在马尾松(Pinusmassoniana)邻株竞争的研究结论一致,即植株干物质显著地低于单株种植处理。
然而,随着竞争胁迫时间的增加,至胁迫后期(150 d),根系生物量与根冠比明显增大,说明较长时间竞争处理可促使杉木通过调整各器官的生物量分配格局,以维持自身将受限的资源最大化地吸收[3],激发植物竞争能力[35],尤其地下部分对资源的竞争表现主导地位[36]。范高华等[35]在研究密度对尖头叶藜(Chenopodiumacuminatum)生物量分配格局的影响时发现,尖头叶藜的生存空间因密度的加剧而遭受限制时,这种拥挤的竞争环境使其生物量的分配发生了改变,即减少地上生物量的分配量,而增加根系生物量,导致根冠比增大,这与本研究有相似之处。但与Watt等[37]对辐射松(Pinusradiata)和汪金松等[38]对臭冷杉(Abiesnephrolepis)在竞争条件下生物量分配的研究结果不同,即存在林木竞争时,杉木根冠比并未发生变化,根冠比与竞争指数之间存在相对较低的相关性。本研究中杉木根冠比受邻株竞争的影响显著,杉木将更多的生物量分配到根系的激烈竞争中,这可能与本研究垂直方向竞争的生长环境有关,这是杉木应对狭小空间竞争的一种适应策略。另有研究报道:植物通常会根据养分胁迫程度对根系分配的生物量进行调整,以保证植物体内部养分平衡和磷素吸收能力[39]。廉满红等[40]研究发现,不论低磷胁迫时期多长,熊猫豆(Phaseoluscoccineus)的根系生物量和根冠比明显增加,这与本文研究结果相一致(图3)。
杉木属浅根系树种,无明显主根,根系分布较浅[41]。但在土壤养分缺乏时,根系会及时启动觅磷策略[21],特别是由于磷素在土壤中移动性差,植物根系对磷的获取主要依赖于根系接触到土壤体积的大小及根形态可塑性变化的强度[42-43]。已有研究表明,在低磷胁迫以及邻株竞争下植物根系会启动一些形态方面的适应性机制[44],如通过根系的长度、表面积、直径和体积等形态学指标的增加或减少,以适应低磷胁迫对自身生长的影响,从而有助于植物更高效吸收和利用土壤磷素[22,45]。本研究表明,竞争处理和供磷水平对杉木根系长度和平均直径增量的影响均存在明显交互作用,其中竞争处理明显促进了根长、表面积、直径和体积等指标的生长(表2),且竞争条件下根长增量明显高于非竞争处理。根系长度的显著增加,有利于根系占领较多的土壤空间和公共资源,扩大其觅养的空间范围,特别是当根系向深层拓殖时,对移动性较差的矿质离子(如磷素)的吸收表现尤其重要。而且,竞争处理明显促进比根长的增大,有利于增强根系活力,这说明邻株竞争条件下,杉木根系可能采取了主动觅养策略[20]。
然而,竞争处理和供磷水平对杉木根系表面积和体积增量指标的影响不存在明显交互作用(表2),这可能是因为竞争处理和供磷水平的影响效应相互叠加的结果[20]。随着供磷水平的增加,根表面积和根体积增量等形态指标呈现显著的先上升后下降趋势,即在胁迫的中期和末期,在低磷胁迫下,杉木幼苗吸收磷素能力最强。这是由于植物为最大限度的利用土壤中有限的磷素,会主动调整根系构型的变化,从而调节根系对养分的吸收[46-47],这种适应性表现为植物大部分根系向介质表层集中,根系分枝角度变大,根向地性减弱,根系变浅[48-49]。在本文设计的竞争条件下通过测定根系参数旨在揭示根系的生长趋向性,基于本文没有测量根系夹角、三维构型等用来反映根系构型的指标、方法,故无法准确定量的分析竞争和非竞争条件下的根系构型发生的改变,而且植物对逆境环境的表型可塑性响应是在体内生理代谢等活动的调控下完成,因此在今后的研究中应进一步加强这方面的研究,以更全面阐明杉木对低磷和竞争胁迫的响应机制,为维持杉木人工林长期生产力提供科学依据。
4 结论
竞争处理中杉木地上部生物量与非竞争对照处理基本持平,根系生物量、根冠比均低于对照处理的单株幼苗。随着胁迫时间的延长,竞争与对照条件下生物量指标均增加。与胁迫前期相比,胁迫后期杉木幼苗地上部生物量、根系生物量、根冠比均明显增大。地上部生物量和根系生物量随着供磷量的增加呈下降趋势。竞争处理下的杉木幼苗根系长度、表面积、平均直径和体积的增量均明显大于非竞争处理;而且这4个根系形态指标均随着供磷水平的增加,呈现出先上升后下降的趋势,而随着胁迫时间的延长呈明显上升趋势。竞争处理明显提高了杉木的比根长,增强根系觅磷能力。
猜你喜欢
农业工程学报(2022年5期)2022-06-22
农民致富之友(2020年8期)2020-05-11
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
现代农业科技(2017年16期)2017-09-22
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
江苏农业科学(2014年1期)2014-07-18
土壤与作物(2013年4期)2013-03-11
