
微生物改造有机质不同演化阶段油气的生成
2019-04-19 09:44丁安娜
沉积学报 2019年2期
丁安娜
甘肃省油气资源研究重点实验室/中国科学院油气资源研究重点实验室,兰州 730000
石油是生物地质作用的产物,无论是海相沉积还是陆相沉积,任何时代生成的石油,都遵循这一规律。石油、天然气的生成过程就是在地壳内一定条件下进行的生物地球化学过程,各种有机质在厌氧条件下被微生物逐级降解,由大分子化合物降解为中等化合物及简单化合物并形成烃类。
1 有机质的厌氧降解
1.1 厌氧降解特征
有机质在有氧降解中已经得到初步改造,沉积物中厌氧降解则是有氧降解的继续,它同有氧降解有明显不同:
(1) 有氧降解的环境必须含有一定氧气,而厌氧分解是在无分子氧的状态下进行。
(2) 有氧分解一般速度快,时间短,而厌氧降解速度慢,时间长。
(3) 在有氧分解中参与作用的为好氧微生物和兼性微生物,而厌氧降解中均为厌氧微生物。
(4) 好氧分解有机质最终产物为CO2,厌氧降解往往形成许多还原的中间产物,其最终产物是低分子烃类和天然气。
1.2 厌氧降解速度
在不同的埋藏深度,降解速度有一定区别,主要取决于以下几个重要因素:
(1) 有机质组成分子量的大小
微生物对营养的摄取是以化合物通过细胞膜的方式实现的,分子量小、成分简单的化合物容易被厌氧微生物降解和摄取,如单糖、二糖等。分子量大的化合物需要多次降解成简单化合物才能被利用。
(2) 有机质在水中的溶解性
微生物的生长必须有适量的水,水除了作为细胞物质组成外,还是细胞吸收营养物质的介质。一般情况下微生物细胞中水占总重量的80%以上,细胞质中发生的生物化学反应都是在水环境中进行的,形成石油的有机质也是通过水来输送的,易溶于水的有机质将较快地被分解利用[1-3]。沉积物中有机质的水溶性是成烃母质的重要性能。
线性高分子能生成氢键,使水分子很快进入高分子结构中,非线性高分子仅有部分区域生成氢键,水分子只能渗入部分高分子结构中,使得线型高分子比同类支键高分子水溶性要好。
有机质分子量越大越难溶,分子量大在水中的黏度大,增加了分子运动的阻力,分子的扩散速度减慢。
结晶程度也有影响,结晶性好的物质由于其分子间的结合力强而难于溶解。淀粉和纤维素都是由葡萄糖环链节单元所组成,纤维素的分子构型是β-1.4型,没有歧化型,分子结构非常对称,通过氢键作用形成坚固的结晶而难以溶解。淀粉含有较多歧化点,不易结晶,易溶于水。
(3) 微生物生长的需要
微生物生长所需的营养主要是一些有机化合物,如脂肪、蛋白质、淀粉、糖类、有用元素等。在三大类生物大分子中,脂类的分子数最小,骨架碳数通常小于100,是多种生物膜的主要组成物质。糖是三大分子中最大的一类,是微生物主要的能量来源。蛋白质的生物功能多,形成各种酶,地质作用极为重要。其中短链、分子量小、极性小的部份优先被降解吸收。
每一种复杂的有机质都能被一种或多种微生物完全或部分地降解成更小的化合物,但降解速度不同。根据上述原则,易降解的有机质有叶绿素、单糖、二糖、淀粉、半纤维素等。较易降解的有脂肪酸、氨基酸、低聚糖、树脂等,难以降解的物质有纤维素,木质素等[4],需要到成熟阶段中后期至高成熟阶段才能被降解(图1)。
2 未熟—低熟油的生成
我国低成熟油气藏的产出层位为上二叠统、侏罗系至新近系。具有高丰度的烃源岩,源岩中富含藻类和水生生物,为腐泥型和偏腐泥型。沉积单元持续沉降,能保持良好的还原环境。但是不同盆地低熟油藏成藏的埋深相差较大,松辽盆地北部为800~1 300 m,江汉盆地1 135~2 812 m,辽河2 300~2 900 m,济阳凹陷1 800~3 500 m,可以看出,低熟油藏需要埋藏至一定深度才能形成[5]。
低熟油生成于地质演化时间较短的低成熟阶段。其成烃母质主要为分子量小、易于水解的物质,如藻类、水生生物中脂肪、蛋白质的短链易溶部分,高等植物的枝、叶、果实,低等植物、草本植物的碳水化合物、淀粉、半纤维素、叶绿素、维生素等,由于从复杂的有机质降解为中等和短链简单化合物、去含氧基团[6-8]、产氢耗氢有一个过程[9-10],低熟油只能在到达一定深度后才能够大量生成。
单糖,如葡萄糖、果糖、甘露糖等六碳糖和木糖、阿拉伯糖等五碳糖易溶于水,是厌氧微生物的良好碳源和能源,微生物可较早地将它们逐步分解并利用。
淀粉是一种多糖(分子量大约为1万至百万),是葡萄糖通过糖苷链连接而成的一种大分子物质,易溶于水。分解淀粉的微生物普遍存在,淀粉水解即成为葡萄糖。在以淀粉作为生长碳源和能源的微生物中,可以利用淀粉酶,将淀粉水解成双糖及单糖后,再被微生物利用。
半纤维素是植物细胞壁的主要成分,易于降解。草本植物中,半纤维素含量很高,占干重的10%~20%,在根、茎、穗中可高达30%。半纤维素的构成多为缩戊糖(木糖和阿拉伯糖)、多缩己糖(半乳糖、甘露糖)及多缩糖醛酸。厌氧菌利用半纤维素酶将半纤维素水解为单糖和糖醛酸,再发酵成乙酸、丁酸、甲酸、乙醇等,除去含氧基团成为烃类。
脂肪酸是生物体中重要的有机质组成,是脂类的重要组成部分,通过脱羧而形成相应碳数的正构烷烃,低碳数脂肪酸对低熟油的生成具有明显的贡献。在未低熟阶段,微生物利用转化有机质有一定局限,使得低熟油的资源量整体上低于成熟油。

图1 厌氧微生物降解有机质随深度变化示意图Fig.1 Changes in degraded organic matter through anaerobia with depth
3 成熟油的生成
成熟油存在生油门限,即主力生油母质开始大量生油的深度。不同盆地不同沉积相带大量生油的深度不同,一般为1 000~2 700 m,门限温度60 ℃~100 ℃。松辽盆地青山口组一段,生油层为深水—半深水沉积,I型有机质为主,成油门限深度约1 100 m,门限温度56 ℃。东营凹陷第三系生油母质属I~II1型,成油门限深度为2 200 m,温度为93 ℃。冀中坳陷下第三系生油层有机质为I~II2型,生油门限深度2 700 m,相应地层温度为95 ℃[6,11]。
成熟油生烃的主力有机质为来自低等水生生物、藻类的脂肪和蛋白质,其次为来自低等植物、高等植物的碳水化合物,它们经过微生物降解[12-13],到达约1 000 m埋深,才能大量生油,形成工业聚集。
脂肪酸初始阶段不溶于水。脂肪酸分子由非极性的碳氢链和极性的羧基基团组成,因此一个分子有疏水和亲水两部分组成,且长链脂肪酸的碳氢链占有分子体积的极大部分,因而总体来说是疏水的,但分子中存在极性基团仍可为水所浸润。经过长期浸泡和微生物作用,至一定深度才能被降解,大量参与成烃。另一个原因是微生物群体存在竞争,总是生长最快的微生物占优势。脂肪分解菌竞争不过淀粉分解菌和糖类分解菌等,当环境中有其他容易利用的碳源与能源物质时,油脂类物质一般不被微生物利用,只有当环境中不存在占优势的其他碳源物质时,微生物才能分解和利用脂类进行生长,微生物学称这种现象为分解代谢抑制。所有脂肪酸都有一个长的碳氢链,以线性碳氢链为主,分支或环状的为数很少,其一端有一个羧基(图2),碳氢链有的是饱和的,如软脂酸、硬脂酸等。有的碳氢链有一个或几个双链,为不饱和脂肪酸,如油酸、亚油酸、亚麻酸等[14]。具有直线型的脂肪酸极易被微生物降解,一个羧基容易被除去,所以到一定深度后脂肪酸可以快速、集中生油,是生成石油的主要组分。
蛋白质是由许多氨基酸通过肽键连接起来的大分子化合物,分子量变化范围很大,从大约6 000到1 000 000。可分为纤维状蛋白质和球状蛋白质。纤维状蛋白质一般不溶于水,影响了其在低熟阶段参与大量成油。球状蛋白质部分溶于水,通过酶催化,蛋白质水解成短肽,在肽酶作用下,进一步分解成氨基酸。
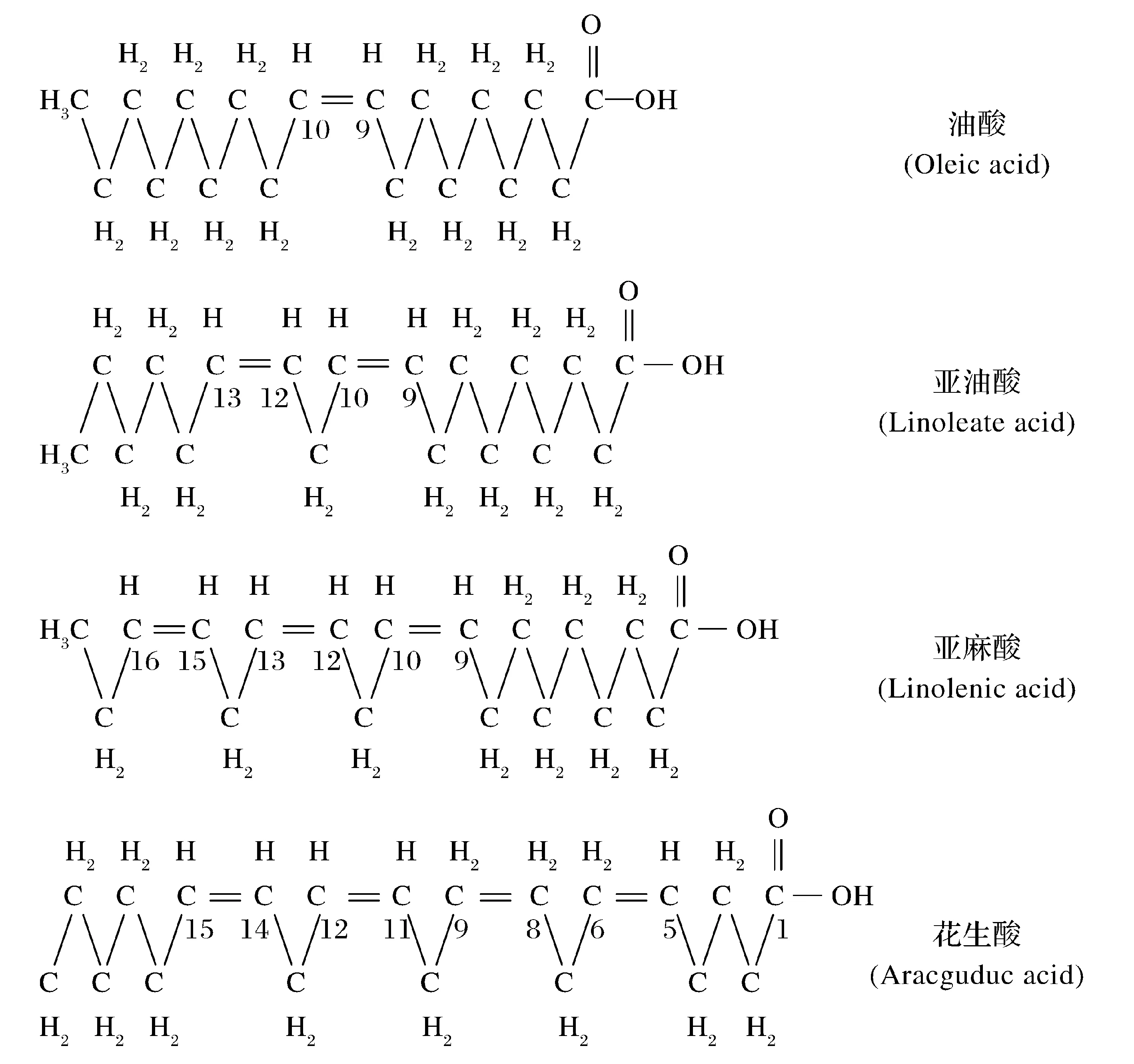
图2 各种不饱和脂肪酸的结构[14]Fig.2 Structure of several unsaturated fatty acids[14]
当蛋白质水解成各种氨基酸时,如甘氨酸、丙氨酸、亮氨酸、天门冬氨酸等,均主要呈直链结构,末端含一个羧基(表1)[15-16]。易除去含氧官能团而成为烃类,为成熟阶段另一个重要的生油母质。
富含脂肪、蛋白质的沉积层多为深湖—半深湖相沉积,低等水生生物、藻类输入为主,为I~IIA型有机质,这种生油岩生油效率高的另一原因是形成有机黏土复合体,深湖相沉积黏土矿物发育,黏土具有较大的比表面积和荷电性,可以通过氢链、离子偶极力、阳离子等链合方式相互结合与吸附,有机质进入黏土矿物层间可被吸附于黏土矿物表层,有机质多带负电荷,易吸附带阳离子的元素形成酶。有机黏土复合体内部发育着各种厌氧微生物(如脂肪酸分解菌、氨基酸分解菌、各种产氢菌、产氢产乙酸菌、脱硫菌、产甲烷菌……),它们互营联合,组成“微生物联合伙伴”,使生油过程快速高效。
陆生植物生长时会尽量伸展枝叶,以更多地获取阳光和空气,吸收CO2,因而富含含氧基团,在空气中生长的时间越长,含量越高。其枝杆以芳香结构为主,以支撑植物的躯体。富含陆生植物的III型有机质其结构以环状芳香结构为主(图3)[16-17]。纤维素是由葡萄糖通过糖苷链连接而成的大分子化合物。高等植物形成的纤维素分子,其葡萄糖数量为14 000个,纤维素分子中若葡萄糖数量大于6个时已不溶于水,能分解利用纤维素生长的微生物均具有纤维素酶。根据柴达木盆地涩北气田气源岩的研究成果,在深度到达1 450 m时才开始出现来自纤维素的成分,因此微生物对III型有机质降解速度到达主生油带深度更大,温度更高。
4 高演化生气阶段
产甲烷细菌是一类形态多样并且具有特殊细胞成分的严格厌氧古细菌,依其生存温度而言,有低温、中温、嗜热、超嗜热产甲烷菌,高演化阶段甲烷气由嗜热产甲烷菌和超嗜热产甲烷菌形成,其产甲烷模式为H2+CO2,位于自然界碳循环食物链的末端。
表1从蛋白质水解液中分离出来的L型氨基酸[15-16]
Table1SeparatedL-aminoacidsfromproteinhydrolysate[15-16]
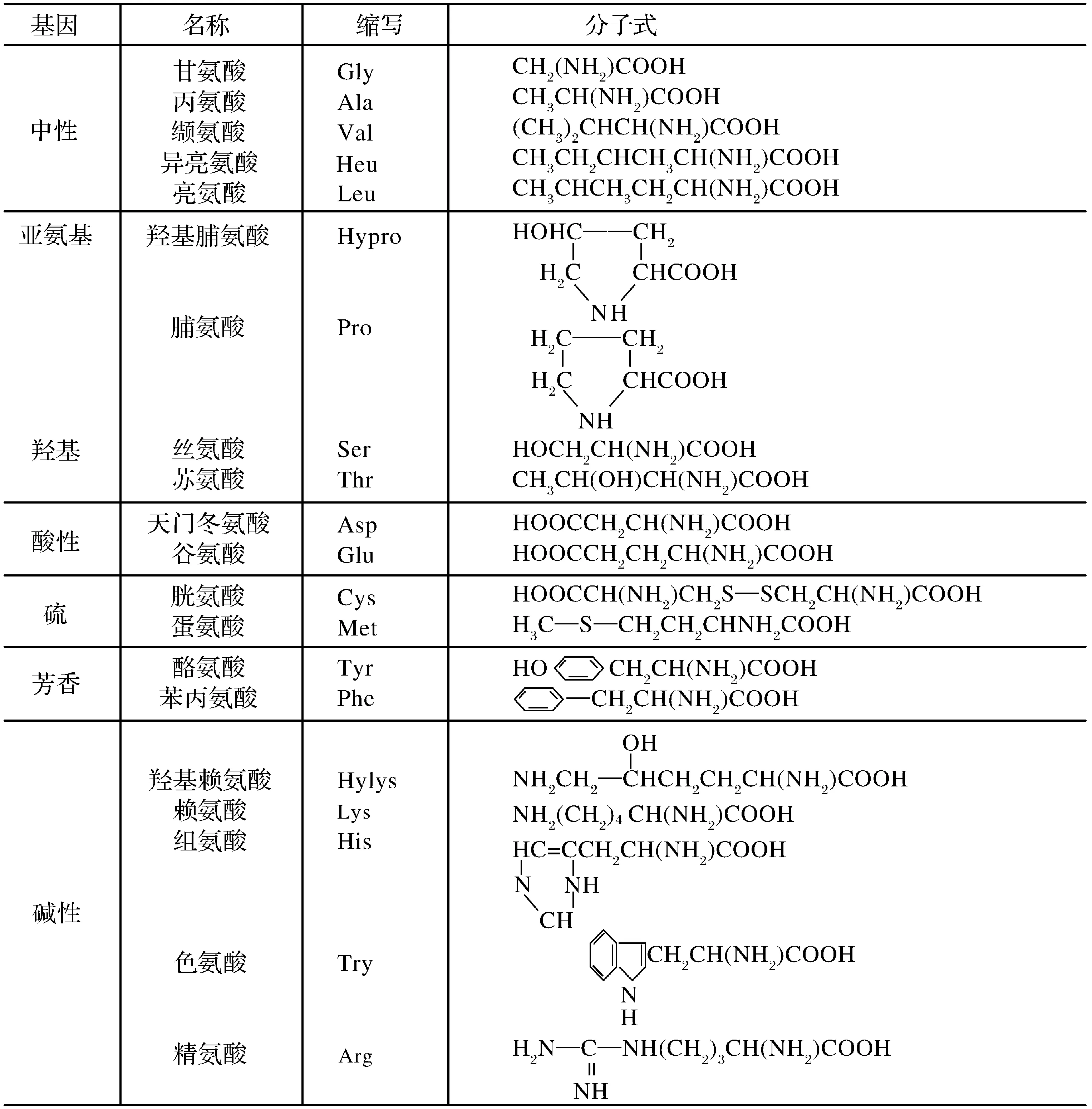
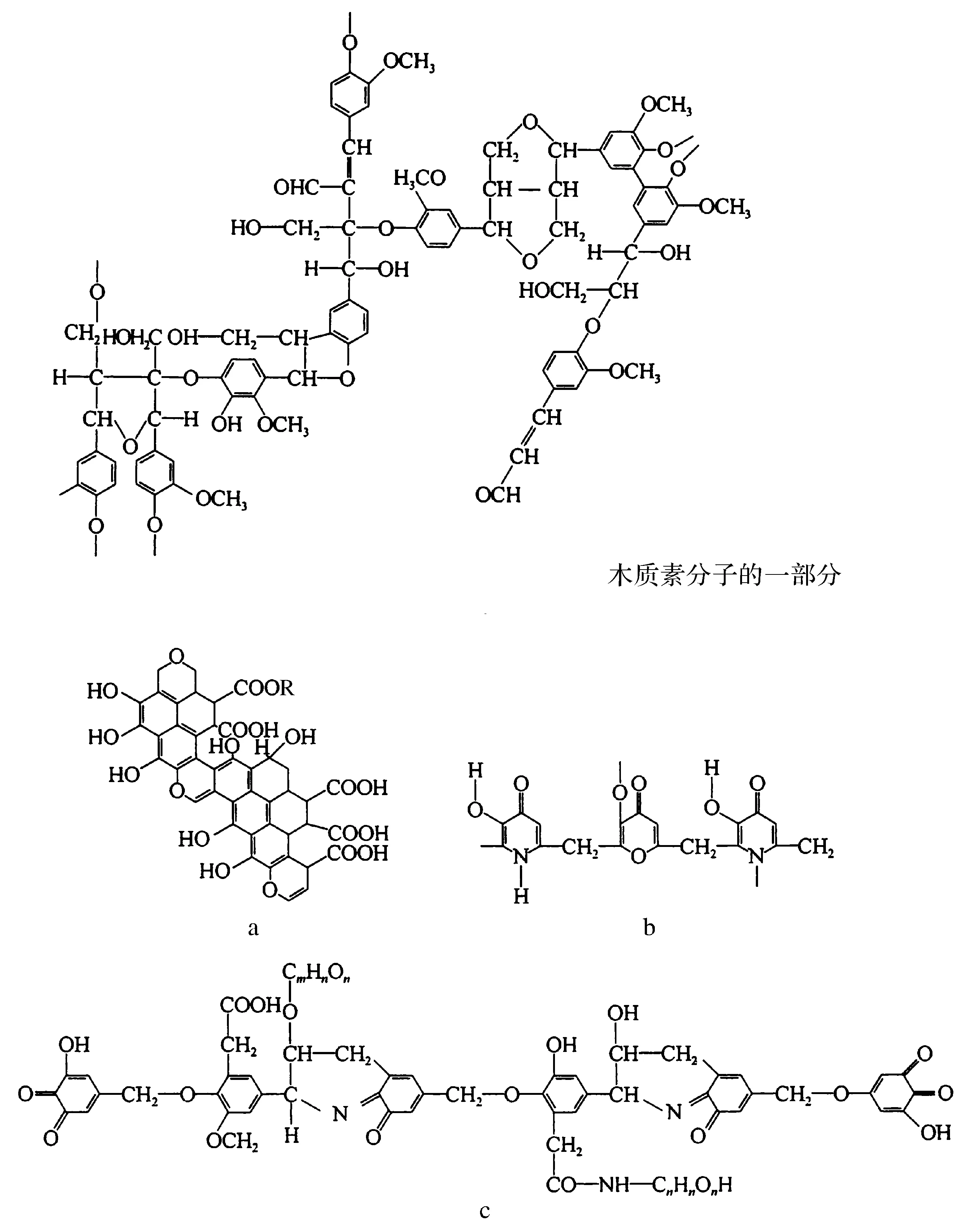
图3 腐殖酸的基本结构[16-17]a.富克斯结构模型;b.费尔伯克结构模型;c.特拉古洛夫结构模型Fig.3 Basic structure of humic acids[16-17]
目前,已成功分离鉴定出许多嗜热产甲烷菌,其最高温度为90 ℃~110 ℃(表2)[18-20]。H2和CO2是沉积物中产甲烷菌能够大量利用的底物,纤维素、木质素、高分子聚合物等许多在成熟阶段难以降解的有机质至高演化阶段可以提供碳源,形成CO2。
纤维素表面覆盖一层含有蜡和果胶的薄膜,不易溶于水。在高演化阶段,纤维素可被嗜热梭状芽孢杆菌等降解,其降解产物为乙醇、乳酸、甲酸等生成甲烷的前身物,通过H2和CO2途径被进一步代谢形成甲烷。
木质素是植物体的重要组成部分,含量仅次于纤维素和半纤维素。木质素中苯丙烷结构单元通过醚键和碳—碳键相联,一般情况下,这些键对微生物分解有极强的抵抗作用,分解速度慢。在高演化阶段,具有胞外酶的微生物可分解木质素,能裂解芳香类化合物,为天然气的形成提供碳源。
微生物学研究表明,在某些环境中产甲烷细菌可以产生少量的乙烷(C2)气体。当培养环境中存有卤化的碳氢化合物,如溴乙烷、二溴乙烷或二氯乙烷等时,在产生甲烷的同时能产生少量的乙烷、乙烯和乙炔。生态学观察表明巴氏甲烷八叠球菌在有乙醇和H2/CO2存在时产生乙烷,嗜热自养甲烷杆菌能将二溴乙烯转化为乙烷,将一溴乙烷转换为乙烷和甲烷[20],高演化阶段少量乙烷及重烃气的形成机理尚待进一步研究。
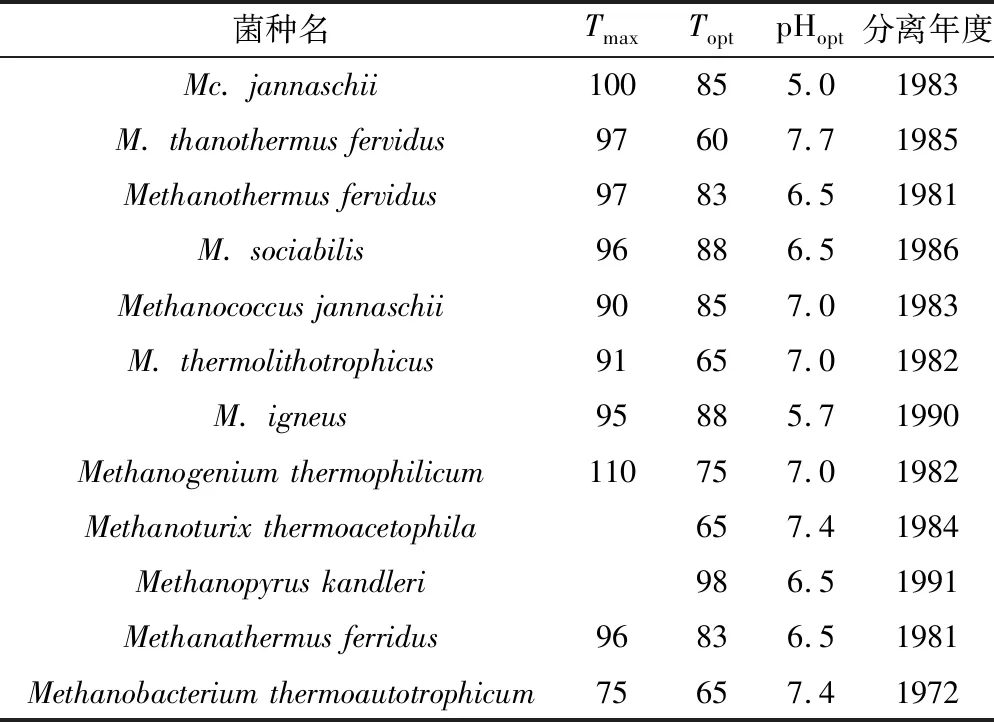
表2 嗜热产甲烷菌特性[18-20]Table 2 Separated out L-amino acids from protein[18-20]
注:Tmax:最大温度(℃),Topt:最适温度(℃),pHopt:最适pH值。
5 微生物形成油气的模式
微生物改造有机质形成石油天然气具有明显的阶段性(图4),不同阶段具有不同的生烃有机质和不同的微生物群体,并形成不同的油气资源,这对于判断油气的成因是十分重要的。
5.1 未低熟阶段——形成生物气、低熟油[21-23]
我国东南沿海全新世为海陆相过渡带,沉积层为河漫滩相、浅海相、河口湾相、湖沼相,厚15~60 m,源岩以II2~III型为主,形成浅层生物气藏。在柴达木盆地三湖拗陷,第四系厚达3 200 m,有巨厚的腐殖—腐泥型或草本腐殖型气源岩,形成了大型生物气藏。形成低熟油藏的微生物均为低温—中温型厌氧微生物,生油母质为分子量小,简单的有机质或经过微生物降解后易溶于水的有机质。
5.2 成熟阶段——形成成熟油藏
形成成熟油的主要组分是脂肪、蛋白质、糖类、淀粉等,这些物质形成了石油中的饱合烃,芳烃馏分主要来自植物。
石油演化程度越低,非烃组分越丰富,重质烃含量高,原油比重大。非烃组成主要是含氧、氮、硫等元素的有机化合物,含氧化合物为酸、醇、酮和醚等。酸类有饱和酸、环烷酸、芳香酸、异戊间二烯酸,还有具甾族结构和霍烷结构的酸类。由于厌氧微生物对非烃的降解,除去含氧、含硫基团,形成更多的轻质烃,随着埋藏深度的增加,非烃组分大大减少,石油比重变轻。
5.3 高演化阶段——形成天然气藏和凝析油藏
在高演化阶段,嗜热厌氧微生物对已经形成的石油和已被改造的有机质继续进行降解形成低分子烃类。C1~C4轻烃比例迅速增加,在地层温度和压力超过轻烃相态转变的临界值时,这些轻质烃就会发生逆蒸发,反溶解于气态烃之中,形成凝析气和富含气态烃的湿气。
干酪根是经过实验室处理的有机集合体,在热压模拟实验中它将受到嗜热菌、超嗜热菌的继续降解而成油成气。温度的增加对有机质的分解起到一定作用,但主要作用是厌氧微生物的参与,没有微生物作用,沉积物中的有机质降解便不可能那么快速,含氧基团无法去除,没有微生物的产氢和用氢,便不可能形成大量的饱和烃类。
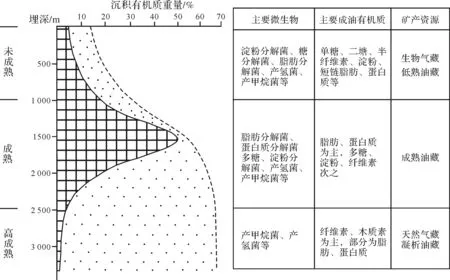
图4 微生物形成油气的模式图Fig.4 Formation of petroleum and natural gas from microbial activity
在许多全岩光片中,可发现生油岩中微生物改造有机质的作用是极其强烈的。较多藻类体已失去藻类的细胞结构,群体球藻中各单体的藻壁被降解而腐化成一体,形成大小不等的絮状、云雾状无定形体,荧光下呈金黄色。有的经微生物强烈改造成无规则外形,有的从一个侧面开始生物降解,另一侧面则保留了藻类体的外形。一部分已形成石油,一部分正在被改造,这就是石油形成过程中的真实面貌。
综上,微生物改造有机质形成石油、天然气是一个自然的过程,只要具有连续沉降的盆地,沉积岩中有机质丰富,生油过程便可发生。成油过程比成气复杂,天然气在埋深数十米,第四纪形成天然气藏。油藏需源岩埋藏至1 000 m以下,第三纪成藏。
微生物具有生理学的多能性和灵活性,很高的生长速率。嗜热菌代表着生命对环境的极限适应能力,也是生物遗传最为丰富的宝藏。细菌是地球上最先出现的生命有机体,它们生存于生命存在的任何角落。微生物无时无刻不与人类相伴,带来危害,也带来福音,形成石油、天然气对人类便是一个最大的恩惠。
参考文献(Rdeferences)
[1] Whiticar M J, Faber E, Schoell M. Biogenic methane formation in marine and freshwater environments: CO2reductionvs. acetate fermentation—isotope evidence[J]. Geochimica et Cosmochimica Acta, 1986, 50(5): 693-709.
[2] 惠荣耀,李剑,张英,等. 生物气源岩评价方法[J]. 天然气工业,2009,29(2):18-22. [Hui Rongyao, Li Jian, Zhang Ying, et al. A study on evaluation procedures of biogas source rocks[J]. Natural Gas Industry, 2009, 29(2): 18-22.]
[3] 阎葆瑞,张锡根. 微生物成矿学[M]. 北京:科学出版社,2000:109-120. [Yan Baorui, Zhang Xigen. Microbial minerogenetology[M]. Beijing: Science Press, 2000: 109-120.]
[4] Zeikus J G. Chemical and fuel production by anaerobic bacteria[J]. Annual Review of Microbiology, 1980, 34: 423-464.
[5] 侯读杰,冯子辉,王铁冠,等. 松辽盆地石油地球化学研究新进展[M]. 北京:石油工业出版社,1999. [Hou Dujie, Feng Zihui, Wang Tieguan, et al. New development on petroleum geochemistry of Songliao Basin[M]. Beijing: Petroleum Industry Press, 1999.]
[6] 王铁冠,钟宁宁,侯读杰,等. 低熟油气形成机理与分布[M]. 北京:石油工业出版社,1995. [Wang Tieguan, Zhong Ningning, Hou Dujie, et al. Formative machine and occurrence of lower-mature oil and gas[M]. Beijing: Petroleum Industry Press, 1995.]
[7] Zobell C E. Geochemical aspects of the microbial modification of carbon compounds[M]//Advances organic geochemistry. New York: Macmilan, 1964: 339-356.
[8] 惠荣耀,丁安娜. 微生物在石油生成中的作用(一):降解和去含氧基团[J]. 沉积学报,2017,35(6):1274-1283. [Hui Rongyao, Ding Anna. The role of microorganisms in the oil generation (I): The degradation of macromolecular organic matter and the elimination of oxygen-containing groups[J]. Acta Sedimentologica Sinica, 2017, 35(6): 1274-1283.]
[9] Zobell C E. Microbial transformation of molecular hydrogen in marine sediments, with particular reference to petroleum[J]. AAPG Bulletin, 1947, 31(10): 1709-1751.
[10] 惠荣耀,丁安娜. 微生物在石油生成中的作用(二)——氢代谢及多源输入[J]. 沉积学报,2018,36(5):1023-1031. [Hui Rongyao, Ding Anna. Role of microorganisms in oil generation (Ⅱ): Hydrogen metabolism and organic matter input from many origins[J]. Acta Sedimentologica Sinica, 2018, 36(5): 1023-1031.]
[11] 黄第藩,张大江,王培荣,等. 中国未成熟石油成因机制和成藏条件[M]. 北京:石油工业出版社,2003:355-657. [Huang Difan, Zhang Dajiang, Wang Peirong, et al. Genetic mechanism and accumulation condition of immature oil in China[M]. Beijing: Petroleum Industry Press, 2003: 355-657.]
[12] Killops S D, Howell V J, Massond M S, et al. Role of bacteria in oil generation from higher plant material: Biomarker evidence from oils from Korea Bay Basin[M]//Manning D C. Organic geochemistry, advances and applications in energy and the natural environment. Manchester: Manchester University Press, 1991: 53-55.
[13] 惠荣耀,丁安娜,连莉文. 甲烷气在低演化沉积物中的形成[J]. 石油勘探与开发,1994,21(4):103-104. [Hui Rongyao, Ding Anna, Lian Liwen. Formaton of methane gas in lower-evolutional sediments[J]. Petroleum Exploration and Development, 1994, 21(4): 103-104.]
[14] 蒂索 B P,威尔特 D H. 石油形成和分布[M]. 徐永元,徐濂,郝石生,等译. 2版. 北京:石油工业出版社,1989:20-21. [Tissot B P, Welte D H. Petroleum formation and occurrence[M]. Xu Yongyuan, Xu Lian, Hao Shisheng, et al, trans. 2nd ed. Beijing: Petroleum Industry Press, 1989: 20-21.]
[15] 汪小兰. 有机化学[M]. 2版. 北京:高等教育出版社,1987:229. [Wang Xiaolan. Organic chemistry[M]. 2nd ed. Beijing: High Education Press, 1987: 299.]
[16] 王启军,陈建瑜. 油气地球化学[M]. 武汉:中国地质大学出版社,1988:73-91. [Wang Qijun, Chen Jianyu. Geochemistry of petroleum and natural gas[M]. Wuhan: China University of Geosciences Press, 1988: 73-91.]
[17] Stevenson F J, Butler J H
a. Chemistry of humic acids and related pigments[M]//Eglinton G, Murphy M T J. Organic geochemistry. Berlin, Heidelberg: Springer, 1969: 534-556.
[18] Stetter K O. Thermophiles: General, molecular, and applied microbiology[M]. New York: John Wiley and Sons, 1986: 39-45.
[19] 和致中,彭谦,陈俊英. 高温微生物学[M]. 北京:科学出版社,2001. [He Zhizhong, Peng Qian, Chen Junying. High temperature microbiology[M]. Beijing: Science Press, 2001.]
[20] 赵一章,连莉文. 产甲烷细菌及其研究方法[M]. 成都:成都科技大学出版社,1999. [Zhao Yizhang, Lian Liwen. Methanogenic bacteria and methodology[M]. Chengdu: Chengdu Science and Technology University Press, 1999.]
[21] Bazhenova O K, Arefiev O
a. Immature oils as the products of early catagenetic transformation of bacterial-algal organic matter[J]. Organic Geochemistry, 1990, 16(1/2/3): 307-311.
[22] 惠荣耀,连莉文. 产甲烷菌等微生物群体在中浅层天然气藏形成中的作用[J]. 天然气地球科学,1994,5(2):38-39. [Hui Rongyao, Lian Liwen. Effects of microbial colony methanogens and others in formation of middle-shallow gas reservoirs[J]. Natural Gas Geoscience, 1994, 5(2): 38-39.]
[23] 丁安娜,孟仟祥,崔明中,等. 微生物改造有机质与未—低熟石油的形成[J]. 天然气地球科学,1999,10(1/2):23-29. [Ding Anna, Meng Qianxiang, Cui Mingzhong, et al. Bacterial reworking of organic matter and the formation of immature-low mature fossil oil[J]. Natural Gas Geoscience, 1999, 10(1/2): 23-29.]
猜你喜欢
中国设备工程(2023年2期)2023-02-13
中国设备工程(2022年12期)2022-07-11
纺织科技进展(2021年3期)2021-06-09
纺织科技进展(2021年3期)2021-06-09
陶瓷学报(2021年1期)2021-04-13
小学科学(学生版)(2020年10期)2020-10-28
中国麻业科学(2019年2期)2019-06-18
西部大开发(2017年7期)2017-06-26
少儿科学周刊·儿童版(2015年7期)2015-11-24
浙江理工大学学报(自然科学版)(2015年7期)2015-03-01
