
碱蓬SgP5CS基因过表达提高拟南芥耐旱性
2019-04-18 11:34郭丹丹杨清华朱丹华金杭霞
浙江农业学报 2019年4期
郭丹丹,杨清华,朱丹华,金杭霞,*
(1.浙江师范大学 化学与生命科学学院,浙江 金华321004; 2.浙江省农业科学院 作物与核技术利用研究所,浙江 杭州310021)
干旱胁迫是影响植物生长的主要非生物胁迫之一,具有重要的农艺学和经济学意义。气候变化预测表明,未来干旱事件频率将会上升[1-2],因此,增强植物耐旱性使植物适应干旱环境是提高作物产量行之有效的方法之一。植物适应环境胁迫重要的途径之一是渗透调节,脯氨酸(proline)是重要的渗透调节物质[3-4],它在稳定膜和细胞结构[5]、保护细胞免受氧化损伤[5-6]和激活植物体内多种反应信号中有着重要作用[7]。
在高等植物中,脯氨酸主要是由谷氨酸通过2种酶作用合成:Δ1-吡咯啉-5-羧酸合成酶(P5CS)和Δ1-吡咯啉-5-羧酸还原酶(P5CR)[8]。合成过程中,P5CS是催化谷氨酸转化为δ1-吡咯啉-5-羧酸盐(P5c)并还原成脯氨酸的关键酶。脯氨酸的重要功能在植物应激防御中得到了广泛的体现,目前已在木薯、蒙古冰草、苜蓿、烟草等植物中发现干旱胁迫下,P5CS基因过表达使植物体内脯氨酸含量上升,增强植物的应急防御能力,提升了植物耐受性[9-12]。
碱蓬(Suaedasalsa)是一种盐生植物,耐盐碱、贫瘠,生长于沿海地区。碱蓬对盐胁迫和干旱胁迫表现出较强的抗逆性,为研究盐胁迫和抗旱相关基因提供了有价值的材料来源[13]。本研究从碱蓬中克隆出P5CS基因,命名为SgP5CS,将其转入到拟南芥中,进行表达模式分析和耐旱性鉴定。
1 材料与方法
1.1 材料
碱蓬种植于浙江省农业科学院温室,光周期16 h/8 h(L/D),温度25 °C,生长1个月。用200 mmol·L-1甘露醇处理植株3 d,采集处理后的碱蓬和未经处理的碱蓬(对照)叶片,立即冷冻并保存在液氮中以备提取RNA;将生长7 d的转基因拟南芥幼苗分别用0 mmol·L-1(对照)和200 mmol·L-1甘露醇进行干旱胁迫2周。
1.2 方法
1.2.1 总RNA提取、cDNA合成和qPCR分析
以经过干旱处理3 d的碱蓬叶片为材料,采用Trizol法[14]提取总RNA,-70 ℃保存。GenBank数据库查找已有的大豆、水稻等植物的Δ1-吡咯啉-5-羧酸合成酶序列,用Clustalw2多序列比对后,设计包含完整开放阅读框(ORF)的简并引物SgP5CS-F(5’-ATGGACGCKWCTAGAGYYTTCGT-3’)和SgP5CS-R(5’-AGGCAGCGGGAGAGAGGACAAGAA-3’)。通过逆转录试剂盒(TOYOBO公司)合成cDNA第1条链。以cDNA为模板使用Pfu酶(TRANSGEN BIOTECH公司)进行PCR扩增,通过1%琼脂糖凝胶电泳检测产物并回收目的条带。使用Roche LightCycler® 480 II定量PCR仪对SgP5CS基因的表达进行检测。
1.2.2SgP5CS基因表达载体构建和拟南芥转化
参照金杭霞等[15]的方法进行表达载体的构建。将含SgP5CS的pEASY-Blunt质粒和pCAMBIA1301-gfp空载体分别双酶切(SacⅠ内切酶+BamH Ⅰ内切酶)。酶切产物用1%琼脂糖凝胶电泳进行分离,用DNA凝胶回收试剂盒回收大小约2 100 bp的目的条带和大小约14 kb左右的pCAMBIA1301-gfp载体片段。T4-DNA连接酶将2个DNA回收片段在室温条件下连接过夜。利用冻融法将重组质粒转入大肠埃希菌DH5α中,涂布LB平板(含100 mg·L-1卡那霉素),37 ℃培养12~16 h,挑单菌落摇菌、抽提质粒,经SacⅠ和BamH Ⅰ双酶切验证后送测序公司测序。测序验证后,命名为SgP5CS-pCAMBIA1301-gfp。将SgP5CS-pCAMBIA1301-gfp重组质粒利用冻融法转化农杆菌EHA105,涂布LB平板(含50 mg·L-1利福平和100 mg·L-1卡那霉素),28 ℃条件下培养形成菌落,挑单菌落进行PCR验证、抽提质粒、双酶切验证。
利用花序浸染法分别将含有重组质粒SgP5CS-pCAMBIA1301-gfp和pCAMBIA1301-gfp空载体的农杆菌转化拟南芥。T0代转基因拟南芥种子在含有50 mg·L-1潮霉素的MS培养基上筛选,选取抗性苗移植,PCR检测为阳性的转基因苗温室培养后收获T1代种子。
1.2.3 转基因拟南芥鉴定
在含50 mg·L-1潮霉素的MS培养基上筛选出SgP5CS过表达转基因株系,在16 h/8h(L/D)光暗周期,25 °C室内培养。提取筛选到的拟南芥抗性苗总RNA,以此为qRT-PCR模板,利用SgP5CS150-F(5’-TGGACCTGACCTCTGC-3’)和SgP5CS150-R(5’-CTTTCCTGGTACTTACTGC-3’)特异引物进行扩增,鉴定转基因拟南芥植株。
1.2.4 转基因拟南芥耐旱性和生理指标的测定
将T2代转基因拟南芥株系和对照组(转pCAMBIA1301-gfp空质粒拟南芥植株)种子表面灭菌,播种于含50 mg·L-1潮霉素的固体MS培养基上。7 d后,选择具有同等生长活力和长势一致的幼苗,分别移植到含0和200 mmol·L-1甘露醇的固体MS培养基上,2周后每个株系分别测定30株拟南芥的根长,并观察植株整体生长状态。脯氨酸含量测定采用茚三酮显色法[16],丙二醛(malondialdehyde,MDA)含量测定使用硫代巴比妥酸法[17-18]。
1.2.5 数据分析
实验数据采用Origin Pro 6进行统计分析和作图。
2 结果与分析
2.1 干旱胁迫下SgP5CS基因在碱蓬中的表达分析
为了探究SgP5CS是否受到非生物胁迫的诱导,对碱蓬幼苗进行了3 d干旱胁迫(200 mmol·L-1甘露醇)。通过qRT-PCR方法测定基因表达量,结果如图1所示。SgP5CS基因在干旱胁迫条件下表达水平是对照的43.7倍,具有极显著差异(P<0.01),说明干旱胁迫可显著诱导SgP5CS的表达。
2.2 SgP5CS基因植物表达载体的构建与验证
将含SgP5CS的pEASY-Blunt质粒和pCAMBIA1301-gfp空载体分别双酶切,回收片段再连接,最终使SgP5CS的ORF插入到pCAMBIA1301-gfp载体的组成型表达启动子35S之后。构建的重组表达载体SgP5CS-pCAMBIA1301-gfp经限制性内切酶SacⅠ、BamH Ⅰ双酶切后,得到2个不同大小的DNA片段,其中SgP5CS基因片段约2 100 bp(图2)。对酶切的载体再次测序验证,显示插入的片段序列与第1次测序结果相同,表明SgP5CS的ORF已经插入到质粒pCAMBIA1301-gfp中,说明植物双元表达载体SgP5CS-pCAMBIA1301-gfp构建成功。

**表示与对照差异极显著(P<0.01)。下同。** indicated significant difference at 0.01 level compared with control group. The same as bellow.图1 干旱胁迫诱导SgP5CS基因表达分析Fig.1 Analysis of SgP5CS gene expression induced by drought stress
2.3 SgP5CS基因转拟南芥植株的获得和筛选
利用花序侵染法将载体SgP5CS-pCAMBIA1301-gfp和空载体转入拟南芥,获得了转SgP5CS基因和转空载体(CK)T0代拟南芥种子,在含有50 mg·L-1潮霉素的MS培养基上筛选10 d后,选取抗性苗移植。对4株已开花的T1代拟南芥植株进行PCR验证,以转空载体的拟南芥植株为阴性对照,在4株抗性苗中都扩增出了约2 100 bp的DNA条带,而阴性对照未扩增出相应条带(图3)。
2.4 SgP5CS在转基因拟南芥中的表达
利用qRT-PCR方法对T2代转基因拟南芥幼苗进行检测,结果显示,相同浓度干旱胁迫下,转基因拟南芥P1、P2和P3株系中均检测到SgP5CS基因的表达且表达量显著上调,尤其P3株系表达量极显著高于对照(P<0.01),而在转空载体对照组中未检测到SgP5CS基因(图4)。
2.5 SgP5CS的过表达对转基因拟南芥耐旱性的影响
分别对SgP5CS过表达转基因拟南芥株系P1、P2、P3和转空载体的对照植株进行干旱(MS+200 mmol·L-1甘露醇)和正常条件(MS)处理。2周后,干旱胁迫下转SgP5CS基因拟南芥株系

1,DNA marker;2,SgP5CS-pCAMBIA1301-gfp酶切结果。1, DNA marker;2, Digestion results of SgP5CS-pCAMBIA1301-gfp. 图2 SgP5CS-pCAMBIA1301-gfp重组质粒双酶切鉴定Fig.2 Double digestion of recombinant plasmid SgP5CS-pCAMBIA1301-gfp

1,DNA marker;2~5,转SgP5CS基因拟南芥植株;6,阴性对照。1, DNA marker; 2-5,Transgenic Arabidopsis plants with SgP5CS gene; 6, Negative control.图3 SgP5CS转基因拟南芥PCR检测电泳图Fig.3 Electrophoresis of PCR amplification products of SgP5CS transgenic Arabidopsis

CK,转空载体拟南芥植株;P1~P3,转SgP5CS基因拟南芥植株。下同。CK, Transgenic Arabidopsis line with empty plasmid; P1-P3, Transgenic Arabidopsis plants with SgP5CS gene. The same as bellow.图4 干旱胁迫诱导拟南芥中SgP5CS基因表达分析Fig.4 Analysis of SgP5CS gene expression in Arabidopsis induced by drought stress
P1、P2和P3的根系长度与对照相比均极显著增加(图5),分别是对照的3.75、3.35和4.63倍。干旱胁迫下植株的根系长度均低于正常条件下,其中,对照植株在干旱处理下根系长度仅为正常条件下生长的17.12%,P1、P2和P3植株分别为65.8%、59.52%和77.27%。同时,在未添加甘露醇的MS培养基上,由根系长度可以看出,P1、P2、P3生长状况与对照相似(图5、图6-a);而干旱条件下生长状况差异明显,SgP5CS过表达转基因拟南芥株系的生长状况明显优于对照,对照植株受胁迫影响明显,表现出严重受损症状(图6-b)。结果说明SgP5CS过表达转基因拟南芥对干旱胁迫的耐受性更强。

图5 干旱胁迫对转SgP5CS基因拟南芥根长的影响Fig.5 Effects of drought stress on root length of transgenic Arabidopsis with SgP5CS Gene

图6 转SgP5CS基因拟南芥和CK干旱胁迫下的生长状况Fig.6 Effects of drought stress on the growth of transgenic Arabidopsis with SgP5CS gene and empty plasmid
2.6 丙二醛和游离脯氨酸含量变化
干旱胁迫(200 mmol·L-1甘露醇)2周后,SgP5CS基因过表达拟南芥株系P1、P2和P3中游离脯氨酸含量升高,与对照具有极显著差异(P<0.01),分别是对照植株的1.61、1.45和1.73倍;在正常条件下SgP5CS基因过表达拟南芥植株的脯氨酸含量与对照植株无明显差异,所有植株干旱胁迫后脯氨酸含量均高于正常条件下生长的植株(图7)。丙二醛含量是衡量植物对极端环境的耐受性指标[19],结果表明,干旱胁迫下丙二醛含量与对照相比极显著下降(图8),分别下降了50.02%、54.35%和74.51%,尤其P3植株中含量仅为对照的25.49%。结果显示:生长状况明显优于对照的转基因拟南芥体内的脯氨酸水平较高,丙二醛含量较低,而表现出严重受损症状的对照植株相反,表明干旱胁迫下转SgP5CS基因拟南芥与对照相比表现出较强的耐旱能力。
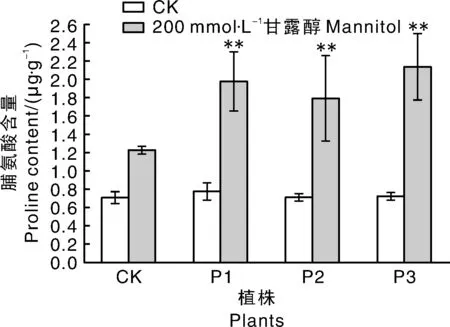
图7 干旱胁迫对转基因拟南芥脯氨酸含量影响Fig.7 Effects of drought stress on proline content in transgenic Arabidopsis
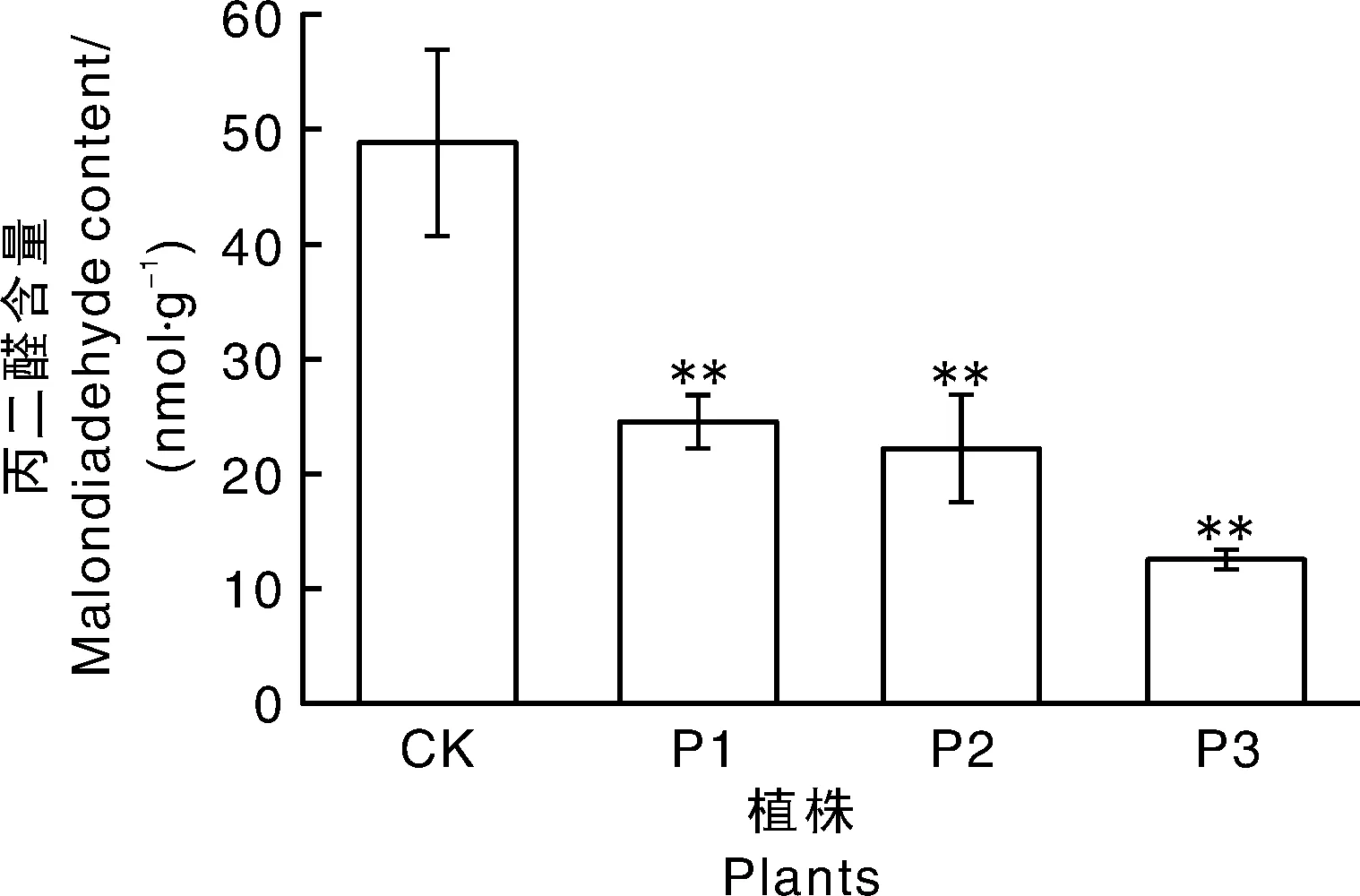
图8 干旱胁迫对转基因拟南芥丙二醛含量影响Fig.8 Effects of drought stress on malondialdehyde content in transgenic Arabidopsis
3 讨论
在高等植物中,脯氨酸在各种胁迫反应中起着关键作用,如应对高盐[20]、缺水[12]、极端温度[21]和重金属毒性[22]等。脯氨酸积累、P5CS基因过表达和对非生物胁迫的耐受性之间的关系已在许多实验中得到充分证明。P5CS基因受特异性调控,可被干旱、盐度、ABA诱导[23],盐胁迫下编码P5CS的基因表达上调,合成更多的脯氨酸帮助植物抵抗渗透失衡的变化[24]。过表达P5CS基因的植物,包括小麦、甘蔗、水稻、马铃薯、柑橘和鹰嘴豆等[25-30],体内脯氨酸积累可以降低脂质过氧化程度,维持细胞结构的完整性来增强植物的应急防御能力,使植物在不同的胁迫环境下维持基本的生理过程。本研究发现,转基因拟南芥SgP5CS基因的表达明显受到干旱胁迫的诱导,这与黄文华[31]对蒙古冰草干旱胁迫下P5CS基因定量表达分析结果一致。
植物主要通过根系来维持自身的生长需求,植物根系脯氨酸的运输和积累可以在极端的环境下通过不同调控途径来促进植株的根系生长[32]。在盐胁迫和干旱胁迫条件下,豇豆P5CS基因在鹰嘴豆和高粱中过表达,脯氨酸含量增加进而促进植物根系伸长和根系生物量增加[30,33]。干旱胁迫会引起植物体内代谢和氧化反应等方面的变化,适量的脯氨酸积累能够使植物对有限的环境条件具备一定的适应能力,这对植物生长具有重要的意义[34]。
本研究克隆碱蓬SgP5CS基因并导入到拟南芥中,使SgP5CS基因在拟南芥过表达后进行相关的指标测定和耐旱性鉴定,结果显示,经干旱胁迫诱导2周后,SgP5CS过表达的拟南芥植株的表现优于对照,具有较长的根系,同时游离脯氨酸的含量显著增加而丙二醛含量显著降低。总之,SgP5CS基因在拟南芥中过表达能够增强拟南芥耐旱性,但SgP5CS基因在干旱胁迫下的调控机制有待进一步研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
江西农业学报(2021年8期)2021-09-08
安徽化工(2021年3期)2021-05-29
园艺与种苗(2020年12期)2021-01-08
上海师范大学学报·自然科学版(2020年5期)2020-12-18
棉花学报(2020年3期)2020-08-08
植物研究(2020年6期)2020-03-05
科学与财富(2019年15期)2019-10-21
上海农业科技(2019年3期)2019-06-25
飞碟探索(2015年9期)2015-11-05
