
无瓣海桑人工混交林结构及幼苗天然更新能力
2019-04-10 03:15:36黄晓敏卢昌义
福建林业科技 2019年1期
黄晓敏,卢昌义
(1.厦门大学环境与生态学院,福建 厦门 361102;2.河口生态安全与环境健康福建省高校重点实验室(厦门大学嘉庚学院),福建 漳州363105)
无瓣海桑(Sonneratiaapetala)是隶属于海桑科(Sonneratiaceae)海桑属(Sonneratia)的红树树种,由于其能在恶劣的滩涂上存活,并能改善滩涂的立地条件,因此1985年从孟加拉国引种至海南省东寨港。目前已在海南、广东、广西和福建等地大面积种植,是华南沿海生态恢复应用中最重要的红树树种[1-3]。为丰富下层植物种类,提高林分稳定性,人工红树林常被进行多样性改造。无瓣海桑人工混交林是否比红树纯林具有更好的生态效益,能否使外来种无瓣海桑演替成上层植物,乡土种乔木演替成中层植物,乡土种灌木演替成下层植物的复合健康林分体系是恢复生态学者关注的重要问题[3]。陈玉军等[4]在海南省三江河两岸的调查表明无瓣海桑群落的物种组成和物种多样性指标稍高于秋茄(Kandeliaobovata)群落,并且在无瓣海桑群落中仅有高龄级个体存在,而秋茄和桐花树(Aegicerascorniculatum)为旺盛增长种群。王旭等[5]调查发现海南省东寨港无瓣海桑与乡土种海桑(Sonneratiacaseolaris)混交林出现了自然更新的秋茄和桐花树等乡土红树植物,认为“引进种+乡土种”的人工恢复模式可以促进乡土红树林的更新。唐以杰等[5]认为混交林和无瓣海桑纯林之间大型底栖动物群落的优势种存在差异,在无瓣海桑林下混交红海榄(Rhizophorastylosa)或木榄这2种乡土红树植物,可提高林地底栖动物的生物量和物种多样性。
福建省厦门市海沧湾无瓣海桑人工混交林于2002年种植,林内红树植物有无瓣海桑、秋茄、红海榄和白骨壤(Avicenniamarina)。目前,该林分已生长16 a,是厦门市海湾典型的无瓣海桑人工混交林。黄龙娇[7]研究了该地无瓣海桑树种的交配系统和基因流情况;文玉叶[8]对该地无瓣海桑的生长情况进行了调查,并着重研究了其繁殖体特性。对于该地无瓣海桑人工混交林的林分结构和幼苗自然更新扩散能力情况未见详细报道。为探究无瓣海桑人工混交林的生态效应问题,本文对厦门市海沧湾16年生无瓣海桑人工混交林结构和幼苗更新扩散能力进行实地调查,以期为今后红树林群落恢复和保护工作提供参考,也为引种外来红树植物的监控提供阶段性的资料。
1 研究区自然概况
研究地点位于厦门市海沧区海沧大道未来海岸东侧西海域中,与厦门岛一水之隔,北邻马銮湾,东邻厦门西港,南面是九龙江入海口。地理坐标位于24°29′02″N、118°02′32″ E。试验地红树林种植地土壤属于沙质土,脚踩泥深度约8 cm,光滩地土壤属于粘壤土,脚踩泥深度20 cm以上。潮汐属正规半日潮,平均潮差3.99 m。高潮时海水平均盐度为28.8‰。属于南亚热带季风性气候,年均气温21 ℃,年均日照时间达2233.5 h,年均降水量1036 mm,全年基本无霜。该地无瓣海桑人工混交林于2002年种植,秋茄的种植密度为50 cm×50 cm,无瓣海桑以2 m×2 m的种植密度间种于秋茄林中,红海榄以2 m×2 m的种植密度间种于靠岸一侧,4 m以内的秋茄林缘,少量白骨壤在秋茄林内混交。
2 调查方法
2.1 群落调查
参考方精云等[9]的经典样方调查方法,于2018年6月对该群落进行样方调查,根据样地实际情况,随机设置了3个5 m×10 m的样方,对样方内的乔木、灌丛、幼苗进行每木调查,分别记录种名、高度、胸径、地径和冠幅。
2.2 重要值计算
根据高度及种群组成情况,将群落划分为4层,高度≥7 m的为乔木一层,高度介于2.5~7 m的为乔木二层,高度介于1.3~2.5 m的为灌木-小树层,高度<1.3 m的为幼苗层[10]。
乔木层和灌木-小树层种群的重要值计算方法参照许会敏等[11]的计算方法。重要值=(相对密度+相对频度+相对显著度)/3。幼苗层种群的重要值,因没有相对显著度,则重要值=(相对密度+相对频度)/2。
2.3 多样性指数计算
参照伊贤贵等[12]计算多样性指数的方法,根据样方调查数据计算多样性指数,包括Shannon-Wiener物种多样性指数、Margalef丰富度指数、Simpson生态优势度指数和Pielou均匀度指数。
2.4 幼苗天然更新和扩散调查
参考廖宝文等[13]调查幼苗更新和扩散情况的方法,于2018年6月分别在人工混交林内、林缘、光滩和互花米草(Spartinaalterniflora)地4种类型样地中设置了52个2 m×2 m样方,调查样方内幼苗种类、数量、高度、地径和冠幅。根据样地实际情况,样方设置具体为:在群落调查设置的3个大样方内随机设置4个样方,共调查12个样方;在无瓣海桑人工复合林东西两边林缘各从南到北随机设置3个样方,在北边林缘按距林0~5、5~10、10~15、15~20 m 4个梯度,每个梯度设置2个样方,在南边林缘随机设置2个样方,共调查16个样方;在光滩按距林缘0~2、2~4、4~6、6~8、8~10 m 5个梯度,每个梯度设置4个样方,共调查20个样方;在互花米草地随机设置4个样方。
3 结果与分析
3.1 林分结构特征
调查结果(表1)表明,该研究地共有红树植物3科4属,包括海桑科的无瓣海桑,红树科的秋茄和红海榄以及马鞭草科的白骨壤。该群落外貌简单,包括乔木层、灌木-小树层和幼苗层。整个群落林相整齐,位于乔木层的无瓣海桑密度为0.05株·m-2,平均高度为7.13 m。其中位于乔木一层的是无瓣海桑,密度为0.03株·m-2,平均高度为8.5 m;位于乔木二层的有无瓣海桑、秋茄和红海榄,密度分别为0.03、0.95、0.03株·m-2,平均高度分别为5.75、4.08、3.88 m。位于灌木-小树层的有秋茄、红海榄、白骨壤,密度分别为0.04、0.02、0.09株·m-2,平均高度分别为2.25、2.03、1.48 m。幼苗层只有秋茄幼苗,密渡为0.14株·m-2,平均高度为17 cm。
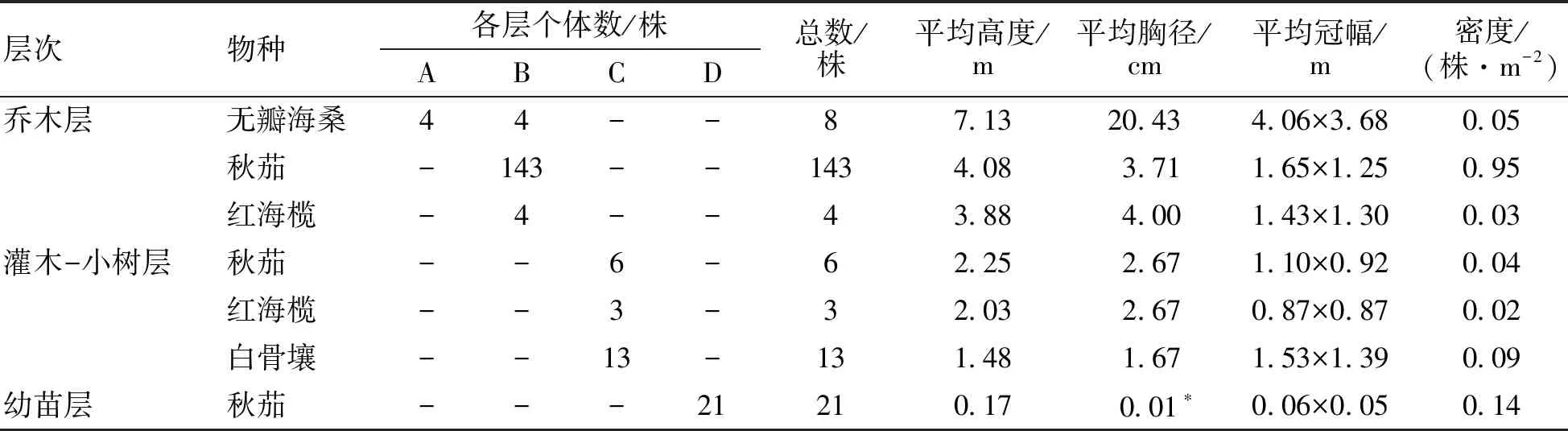
表1 无瓣海桑人工混交林林分结构特征
*:-为没有调查;*为平均地径(由于植株高度<1.3 m没有胸径);A为乔木一层;B为乔木二层;C为灌木-小树层;D为幼苗层;下同。
3.2 重要值
重要值是评价某一种植物在湿地群落中作用的综合性数量指标,是研究群落结构的重要指标之一,也是表征优势种的重要指标[14]。由表2可以看出,在乔木层中无瓣海桑和秋茄的相对频度都为40.0%,但由于秋茄的种群数量大于无瓣海桑的种群数量,所以秋茄的重要值最大,为57.1%,无瓣海桑的重要值为35.1%,而红海榄的重要值最小。在灌木-小树层,秋茄有小树生长,并且重要值最大;白骨壤相对频度虽较小,但其相对多度最大,所以其重要值也较大。红海榄在灌木-小树层的重要值比在乔木层的大。幼苗层只有秋茄生长。
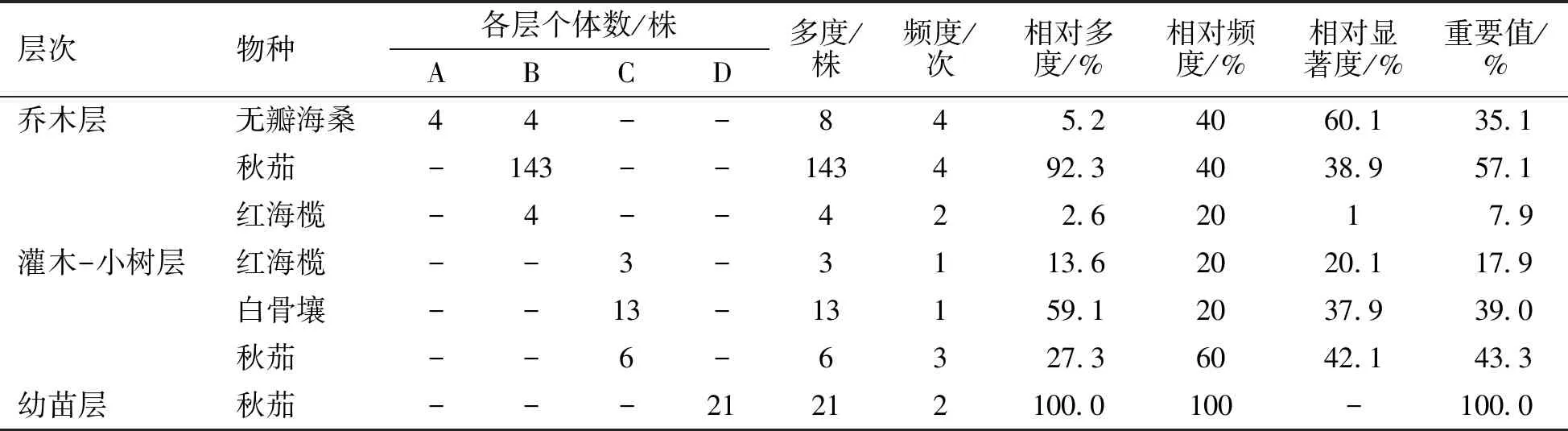
表2 无瓣海桑人工混交林不同层次物种重要值
3.3 物种多样性
物种多样性在某种程度上和生态系统之间的关联性能够反映生态系统的稳定[15]。由表3可以看出,该群落的多样性指数值分别为:Shannon-Wiener指数值为0.56,Margalef指数值为0.57,Simpson指数值为0.26,Pielou指数值为0.41,物种多样性较低。乔木层、灌木-小树层和幼苗层3个层次的物种多样性指数、丰富度指数、生态优势度指数和均匀度指数变化趋势较为一致,表现为灌木-小树层>乔木层>幼苗层。

表3 无瓣海桑人工混交林整个群落和不同层次的多样性指数
3.4 幼苗天然更新和扩散
在无瓣海桑人工混交林内、林缘、光滩和互花米草地4种类型样地设置的52个2 m×2 m的样方内,均未发现有无瓣海桑幼苗;而本地种秋茄和白骨壤都能进行天然更新和扩散。从表4可以看出秋茄在4种类型样地中都进行了天然更新和扩散,其中秋茄幼苗在光滩分布最多,达1.11株·m-2,其次是互花米草地,为0.44株·m-2,最少的是在林缘,仅为0.28株·m-2。白骨壤幼苗只有在互花米草地中生长,密度为0.06株·m-2。
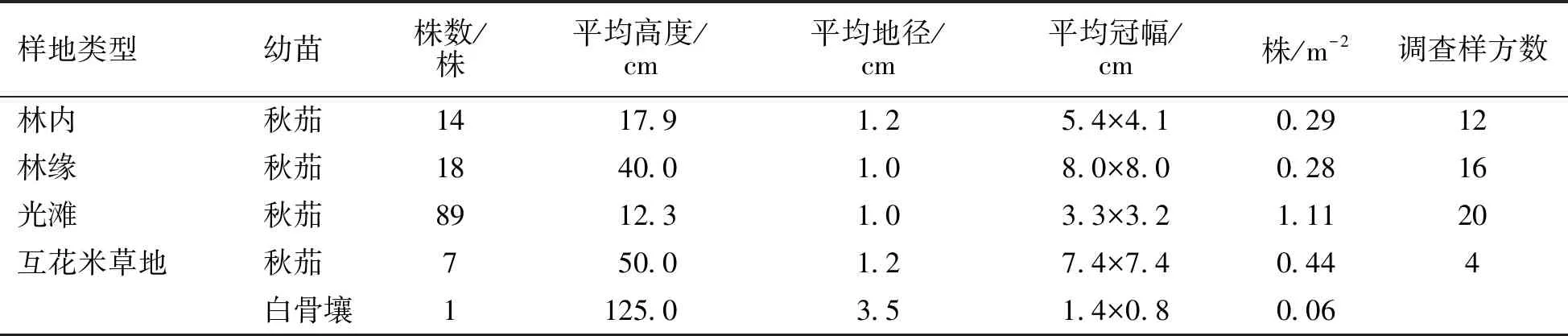
表4 调查样地内乡土红树植物幼苗情况
4 结论与讨论
研究结果表明,该研究地群落空间层次分层明显,各物种占据不同生态位,充分利用生境资源正常生长,在景观搭配和防护效用上起着不同的协调互补作用。乡土红树植物秋茄在该群落中占有绝对优势,对当地的环境适应能力比外来物种无瓣海桑更强。采用“引进种+乡土种”的人工种植恢复模式可增加群落的物种多样性,使群落层次更加丰富,群落结构更加稳定,促进乡土红树植物的天然更新和扩散。调查样地未发现无瓣海桑幼苗,说明其在该地天然更新和扩散极其困难。无瓣海桑对乡土红树植物生长和幼苗天然更新、扩散无抑制作用,并且能为乡土红树植物的天然定植和生长创造有利条件。在人工林恢复建设中可考虑以乡土树种为主,引进种与乡土种相结合的方式。但引进种植外来植物仍然有必要不断跟踪监控,以获取更多的现状和动态资料,全面掌握外来生物的引进和合理利用。
厦门市海沧湾无瓣海桑人工混交林于2002年人工种植,群落已有16 a。由于不同红树植物种类自身特性和对生境的要求不同[16],因此按照无瓣海桑、秋茄、红海榄和白骨壤各自的生长特性,将其按一定位置人工种植在海滩上排列成带。无瓣海桑和秋茄为这个群落的建群种,无瓣海桑相间种植在秋茄林中,白骨壤和红海榄数量较少,分散在群落的较高潮位。群落空间层次分层明显,位于乔木一层的只有无瓣海桑,密度为0.03株·m-2,平均高度为8.5 m;位于乔木二层的红树植物按平均高度从高到底排列依次是无瓣海桑、秋茄、红海榄;位于灌木-小树层的红树植物按平均高度从高到底排列依次是秋茄、红海榄、白骨壤。无瓣海桑虽然位于乔木一层,但现存密度小,对其下层红树植物不造成遮光影响。各红树树种在群落中占据不同生态位,充分利用生境资源正常生长,并且各自树形有着明显差异,在景观搭配和防护效用上也起着不同的协调互补作用。幼苗层有乡土红树植物秋茄幼苗的更新,而并未发现无瓣海桑幼苗和历年各级小树,可见无瓣海桑在林内自然更新成林困难,对乡土红树植物的更新也没有抑制作用,而且还为乡土红树植物幼苗定植提供了有利条件,促进整个群落的自然更新。乡土红树植物秋茄在乔木层、灌木-小树层和幼苗层中均有分布,并且在各层次秋茄的重要值都为最大,可见乡土红树植物秋茄在群落中占有绝对优势,对当地的环境适应性较强。
该研究地各多样性指数与珠海无瓣海桑人工林多样性指数[17]比较可以发现,其数值略小于珠海的无瓣海桑人工林多样性指数值,并且两地物种多样性指数均比陆地植被群落的物种多样性低,主要因为海岸潮间带环境恶劣,能够生长的植物不多,并且也与滩涂生境、人工种植方式、种植年限以及演替程度有关。乔木层多样性指数值比灌木-小树层要低,由于乔木层各树种数量分布不均匀,而灌木-小树层的物种数量分布均匀,所以灌木-小树层多样性指数值大,群落结构较为稳定,优势度大,是生长旺盛的层次。
在4种类型样地内进行幼苗天然更新和扩散情况的调查发现,4种类型样地中均未发现无瓣海桑幼苗。彭友贵等[1]研究发现盐度高于5‰时,无瓣海桑种子发芽开始受到抑制;高于8‰时,幼苗生长开始受到抑制;盐度高于15‰以上时,种子不能出土萌发。该研究地高潮时海水平均盐度为28.8‰,厦门其他海域海水的盐度更高,因此盐度很可能是通过抑制无瓣海桑种子萌发、幼苗生长而限制其自然更新和扩散的主要原因。该研究样地中乡土红树植物秋茄和白骨壤都能进行天然更新和扩散,无瓣海桑对乡土红树植物的天然更新成林和扩散没有产生影响。林内和林缘秋茄幼苗密度较小,在光滩和互花米草地秋茄幼苗密度较大,并且在互花米草地出现了白骨壤幼苗,张乔民等[18]研究发现红树林可以生长在各种底质的海岸上,以淤泥质潮滩最普遍和生长最好。何琴飞等[19]研究也发现红树林生长较好的立地类型为土壤养分较高的内、中滩泥质或沙泥质土。而该研究地林内和林缘都为沙质土,光滩和互花米草地为淤泥质土,因此土壤质地很可能是影响乡土红树植物幼苗扩散分布的原因。
*:感谢厦门大学环境与生态学院诸姮老师协助数据分析,感谢杨志伟老师提供研究区红树林的种植资料,感谢参与本次调查工作的厦门大学环境与生态学院王秀丽、宋瑞瑞、彭建同学。
猜你喜欢
热带林业(2023年3期)2023-10-30 10:38:34
小猕猴学习画刊(2022年4期)2022-05-05 01:52:59
广东第二课堂·小学(2021年10期)2021-12-03 04:35:50
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:32:10
海洋通报(2020年5期)2021-01-14 09:27:04
天然产物研究与开发(2016年6期)2016-06-05 10:29:31
艺术评鉴(2016年13期)2016-05-30 16:19:05
发明与创新(2015年6期)2015-12-26 11:17:18
发明与创新·中学生(2015年6期)2015-06-01 00:40:24
发明与创新(2015年22期)2015-04-17 23:22:59
