
基于非标记定量法的三疣梭子蟹精子塞蛋白质组学研究
2019-03-21 09:04宣富君赵雪冰嵇雅芹刘千姿鲍成满张春丽
浙江海洋大学学报(自然科学版) 2019年5期
宣富君 ,何 杰,赵雪冰,嵇雅芹,刘千姿,鲍成满,张春丽
(1.盐城师范学院海洋与生物工程学院,江苏盐城 224002;2.江苏省盐土生物资源研究重点实验室,盐城师范学院,江苏盐城 224002;3.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江舟山 316021;4.华南农业大学海洋学院,广东广州 510642)
精子塞是交配过程中雄性个体在雌性生殖道中形成的一种障碍物,形式多样,一般呈固态胶质状[1]。它广泛存在于动物界,包括线虫[2]、昆虫[3-4]、蜘蛛[5]、爬行类[6]和哺乳动物[7-8]等各大门类,通常被解释为雌性在多次交配过程中雄性个体间竞争的结果[9]。然而,尽管距文献第一次报道已有160 a[10],但我们现在对精子塞的认识还十分有限,对它的起源问题[11-12],大多数物种精子塞的成分、形成机制及功能问题均尚未明确[5,10]。阐明精子塞存在意义的最好途径就是充分揭示它的组成成分和形成机制[8]。目前,物种间有关精子塞的研究仍以解剖学组织学为主[5,10,13],而在分子生物学层面,仅少数物种涉及[14]。根据现有的报道,精子塞通常含有大量精液成分,形成则基本依赖于交配过程中所接受的精液蛋白[12,15-17]。根据同科锯缘青蟹Scylla serrata 人工模拟精子塞的工作揭示三疣梭子蟹Portunus trituberculatus 的这种精子塞至少部分可由雄性精液和雌性分泌物共同作用的产物组成[17],而目前国内外关于这类精子塞的组成成分、形成机制及相关功能,尤其是梭子蟹这种可能基于两性分子间的功能机制研究并不多见,基本还处于空白[14]。
生物大分子质谱技术和生物信息学分析的发展为蛋白质组学研究提供了有力的技术支持,质谱已成为蛋白质鉴定与定量的重要工具[18-19]。基于质谱的非标记定量检测方法已应用于临床样本比对、实验室生物信号通路研究等各个方面[20-21]。利用基于质谱的非标记定量技术进行蛋白质组定量的方法优势明显:①灵敏度高:可检测出低丰度蛋白;②分离能力强:可分离出酸/碱性蛋白,小于10 KD 或大于200 KD 的蛋白、难溶性蛋白等;③适用范围广:可以对任何类型的蛋白质进行鉴定,包括膜蛋白、核蛋白和胞外蛋白等;④减少了前处理过程中的样品损失,在肽段的检测和蛋白质覆盖率方面具有优势;⑤采用单个样品单独分析,不受样品来源和数目的限制;⑥方便与蛋白质绝对定量技术相结合,对生物体系中表达蛋白质绝对含量进行测定;⑦自动化程度高:液质连用,自动化操作,分析速度快,分离效果好。
三疣梭子蟹是我国重要的海洋经济蟹类,近98%的天然资源分布在我国的大陆架上,2015-2016年海捕量稳定在55 万t 左右[22]。本研究旨在运用基于质谱的非标记定量的方法,分析三疣梭子蟹交配活动所形成精子塞的蛋白质组成,鉴定精子塞形成可能的关键蛋白,为进一步探讨精子塞的形成与功能机制及后期人工授精等相关工作奠定基础。
1 材料与方法
1.1 实验材料
本实验于2015年9月10 日从浙江舟山养殖基地选取健康且未交配的成熟雄蟹25 只(头胸甲宽135~150 mm;硬壳,处于蜕壳间期C),待交配雌蟹25 只(头胸甲宽110~130 mm;硬壳,游泳足倒数第二关节边缘出现血丝,处于蜕壳前期D1-D2),分别暂养于实验室两个长宽高为3 m×3 m×1 m 的水泥池内需。选择标准参照宣富君等[11]三疣梭子蟹生殖蜕壳及交配行为的观察研究。
待熟悉室内养殖环境(至少1 周),选取其中12 对健康雌(游泳足倒数第二关节边缘出现明显红色月牙,处于蜕壳前期D3-D4)雄个体并移入实验室定制的12 只长宽高为0.5 m×0.5 m×0.5 m 的玻璃缸内进行配对实验。期间,利用红外线摄像对其交配行为进行实时监控,并适时解剖交配后的雌蟹获取完整精子塞;将所获完整精子塞分为上下两部分-80℃低温保存,其中4 个精子塞的上下部分送上海美吉逾华生物医药科技有限公司测序,结果用于本文精子塞蛋白成分的比较分析。实验所用试剂均为国产分析纯。
1.2 主要仪器
高校液相色谱仪(Waters,USA),色谱分离90 min (A:0.1% 甲酸,B:ACN,0.1%甲酸,流速:300 nL·min-1,柱温:40℃)。质谱仪(Thermo Fisher Scientific,USA),MS 扫描范围(m/z) 300~1 200,质谱分辨率70 K。数据采集软件:Thermo Scientific Proteome Discoverer softwareversion 1.4。反相柱信息:Thermo Scientific Acclaim PepMap C18 column (100 μm×2 cm,3 μm particle size)。
1.3 实验方法
1.3.1 样品蛋白提取
取样本液氮研磨。加入裂解液(7 M 尿素,2 M 硫脲,0.1% PMSF,65 m MDTT)裂解样品。冰浴超声,超声条件为70~75 W,超声5 s 停10 s,超声3~5 min,冰上放置40 min。离心30 min (14 000 r·min-1、4℃),取上清。按照体积比(样品:丙酮为1 :5)向样品中加入预冷的丙酮,离心20 min (14 000 r·min-1、4℃),弃上清。待残余的丙酮挥干后向样品管中加入1 mL 裂解液复溶沉淀,备用。
1.3.2 还原烷基化和酶解
取出需要的样本量加入终浓度10 mM DTT,在56℃下反应30 min 后加入终浓度20 mM IAA,室温下避光反应30 min。每管各加入预冷的丙酮(丙酮:样品体积比为5:1),-20℃沉淀2 h。离心20 min(14 000 r·min-1,4℃),取沉淀。用含1 M 尿素的TEAB 溶解液20 μL,混悬,充分溶解样品。取样品置于2 mL 超滤离心管,4℃下10 000 g 离心3 min,反复至样本完全超滤完成。按照质量比1:50 (酶:蛋白)加入Trypsin 在37℃酶解15 h。终止:酶解液加入终浓度0.5%的TFA 终止酶解,浓缩冻干。
1.3.3 Bradford 定量
准备BSA 标准品的标准曲线,8 个的标准品量(0.5 μg·μL-1BSA)依次为0,1,2,4,8,12,16,20 μL,向每管添加纯水,使最终体积为20 μL。将样品稀释5 倍,每个样品各取20 μL 至管中。向每管添加180 μL蛋白质分析反应剂,混合,室温培养10 min。测量595 nm 下的吸光度,依据标准曲线计算出样品浓度。
1.3.4 液相色谱-二级质谱联用
参考YIN Xuefei,et al[23]的分析方法,样品在高pH 反向色谱中分离,然后进入串联质谱分析。
1.3.5 搜库和数据分析
质谱原始数据采用Mascot 软件比对uniprot 数据库(https://www.uniprot.org/),然后对鉴定到的蛋白进行GO 功能注释和KEGG 通路分析。
2 结果与分析
2.1 数据定量分析
通过将得到的RNA-SEQ 数据库匹配的2 680 张谱图,85 个蛋白家族,Uniprot 数据库匹配1 074 张谱图,79 个蛋白家族使用PEAKS Label free quantitation 模块进行非标记定量。以P<0.05 且蛋白相对丰度>1.2 或<0.8 为质控标准,得到差异定量蛋白结果共22 个蛋白家族。
2.2 GO 分类结果分析
按照Uniprot 数据库中各蛋白质的GO 注释分别进行生物过程(Biological Process,BP)、细胞组分(Cellular Component,CC),分子功能(Molecular Function,MF)分类。由图1 可知,共计生物学过程56 条,细胞组成45 条,分子功能29 条。差异表达蛋白主要来自于细胞、细胞组分、细胞器以及大分子复合物,以结合和催化活性功能为主,70%的蛋白质主要参与代谢、细胞以及单生物等生物学过程。通过对差异蛋白进行功能更细致的GO 的三级、四级分析可知,差异蛋白主要是定位于细胞内细胞器,发挥核苷结合、核苷酸结合、核苷磷酸结合、核酸结合、负离子结合、转移酶活性、转移含磷基团等分子功能,参与大、小分子代谢过程以及磷代谢过程等生物学过程。
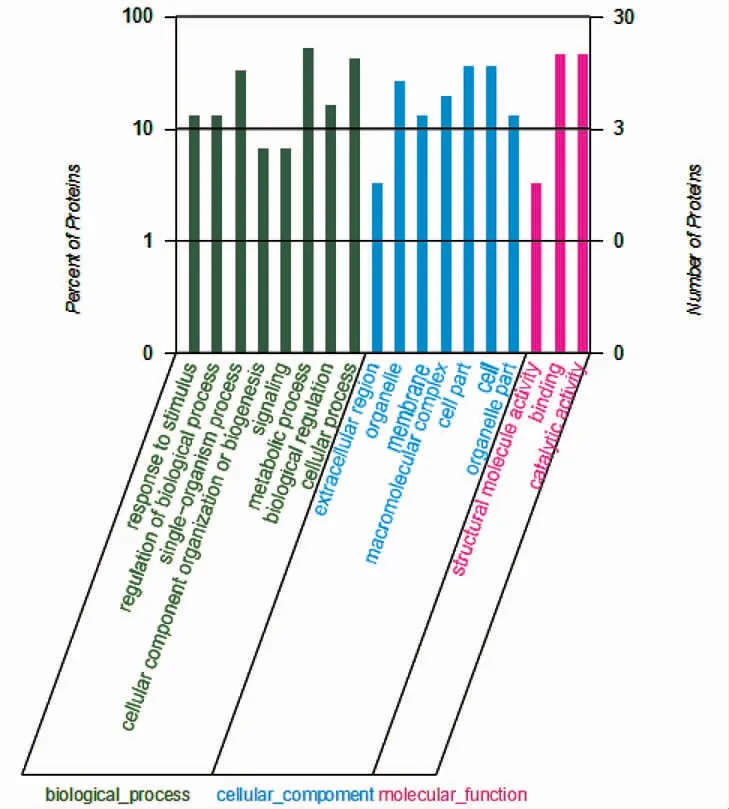
图1 差异蛋白的GO 二级分析结果图Fig.1 GO analysis of differently expressed proteins at the 2nd level
2.3 KEGG 分析
KEGG 分析显示的差异蛋白质可能参与的具体生物学通路前20 个条目(图2、图3)。三疣梭子蟹精子塞差异蛋白富集的主要信号通路有催产素信号通路,内质网蛋白加工,甲状腺激素合成,甲状腺激素信号通路等。对差异蛋白做KEGG 注释,根据差异蛋白参与的KEGG 代谢通路进行分类,与代谢功能相关的Global and overview maps 通路,与有机系统相关的Immune system、Endocrine system,与环境信息处理相关的Signal transduction 通路,与遗传信息处理相关的Folding,sorting and degradation 通路,与细胞过程相关的Transport and catabolism、Cell growth and death 通路对形成三疣梭子蟹精子塞关键蛋白具有重要意义。
2.4 表达差异分析
以P<0.05 且(蛋白相对丰度差异倍数<0.8 或>1.2)为标准筛选显著差异表达蛋白,针对两两分组的精子塞上下两部分的差异表达蛋白进行GO 注释的统计,结果显示未发现蛋白表达水平上调,发现下调共计27 个蛋白表达水平显著(图4)。其中生物学过程共10个蛋白表达水平下调(生物调节1 个,细胞过程3 个,代谢过程3 个,生物过程调节1 个,应激反应1 个,单生物过程1 个);细胞组分共9 个蛋白表达水平下调(细胞3 个,细胞区域3 个,大分子复合物1 个,细胞器2 个);分子功能共8 个蛋白表达水平下调(结合5 个,催化活性3 个)。
3 讨论
目前在研究啮齿动物时对精子塞中有4种假设:(1)精子释放假说:当精子塞解体时,允许精子在雌性生殖道内逐渐释放;(2) 精子运输假说:阻止精液在射精后流出阴道,或以其他方式促进精子通过子宫颈进入子宫;(3)交配刺激假说:促进交配刺激;(4)再次交配阻碍假说:阻止已经受精的雌性再被其他雄性受精[24]。每种功能假说均有相应证据支持,由于精子塞可能有助于交配刺激,这可能导致收缩,同时肌动蛋白对于细胞的收缩是必不可少。因此三疣梭子蟹蟹精子塞的关键蛋白可能包括肌动蛋白,这与我们的研究结果一致,所以我们的研究结果更支持交配刺激假说。
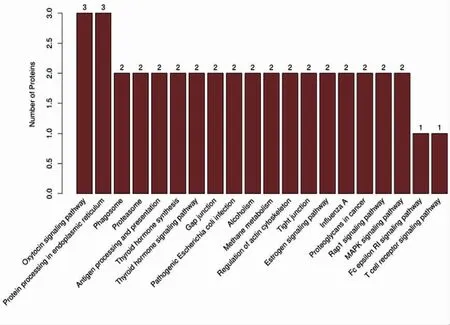
图2 KEGG 分析结果(前20 条信号通路)Fig.2 KEGG pathway analysis of differently expressed proteins (top 20)
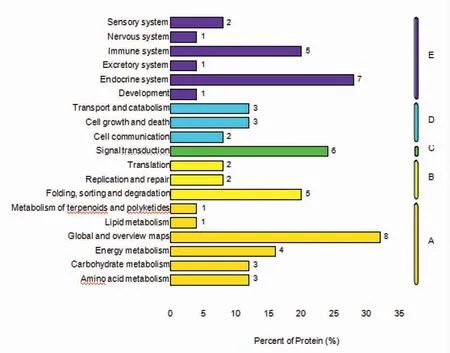
图3 KEGG 注释统计图Fig.3 KEGG annotated statistical chart
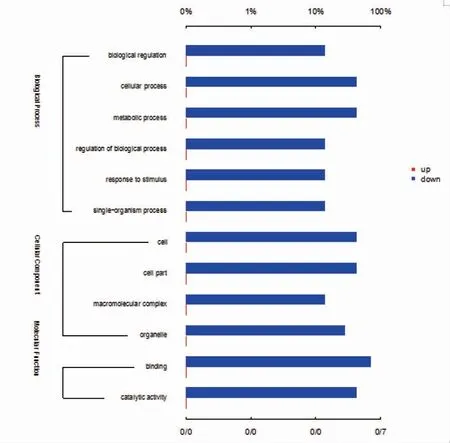
图4 上下调蛋白GO 注释柱形图Fig.4 GO annotated chart of up-down regulatory proteins
现有研究表明,精子塞通常含有大量精液成分,形成基本依赖于交配过程中所接受的精液蛋白[12,15-17]。例如,利用蛋白质组和RNA 干扰技术,ROGERS,et al[5]对疟疾传播者疟蚊Anopheles gambiae 精子塞的形成进行了研究,结果表明该精子塞主要通过一种名为转谷氨酰胺酶(雄性附腺前端分泌)的精液蛋白作用,使底物蛋白Plugin(另一种精液蛋白,富含谷氨酰胺残基,由雄性附腺后端分泌)发生交联反应凝聚而成。该机制与报道的有关哺乳动物精子塞的形成十分类似[25-26],但后者相对复杂,以小鼠Mus musculus 为例,其精子塞形成可分为2 个阶段:首先由其雄性生殖系统贮精囊分泌的具有自凝功能的底物蛋白SVSⅠ-Ⅲ发生自聚反应,形成以二硫键链接的高分子复合物HMWCs,随即雄性小鼠前列腺会分泌出作用蛋白TGM4(也是一种转谷氨酰胺酶,后者促使HMWCs 凝集,最终形成胶状固态精子塞[10,12]。此外,精子塞也可能是雄性生殖系统不同部位分泌的不同精液蛋白在雌性生殖道内形成的多个部分组成,如果蝇Drosophila melanogaster 的精子塞[4]:它可分为前后两个部分,前面主要由副性腺分泌的Acp36DE 蛋白组成,后面则是射精管球分泌的具自发荧光功能的PEB-me 蛋白,由于形成时间上的差异,精荚等遗传物质主要集中在前面部分。这种结构与三疣梭子蟹的精子塞类似[13-14],梭子蟹的精子塞也明显主要由前后两个部分组成,即顶部含大量精荚的雄性基质MM 和底部半透明部分TPSP[14]。与小鼠、疟蚊不一样,三疣梭子蟹精子塞的形成似乎与雌性分泌物有关,这与我们研究结果中三疣梭子蟹精子塞差异蛋白富集的主要信号通路是催产素信号通路相一致。因此,我们推测:梭子蟹科精子塞的形成很可能涉及一种基于两性分子间作用的全新机制,即以雄性精液蛋白为底物蛋白,而雌性纳精囊的分泌物为作用蛋白。
从非标记定量分析显示的结果来看,三疣梭子蟹的精子塞上下部分中共有22 种蛋白家族的表达存在显著差异,GO 功能分析结果显示:差异蛋白主要参与代谢、细胞以及单生物等生物学过程;发挥结合和催化活性的分子功能;主要定位于细胞、细胞组分、细胞器以及大分子复合物。且KEGG 通路富集分析显示:差异蛋白主要富集的信号通路是催产素信号通路和内质网蛋白加工。经过KEGG 注释通路分析显示精子塞中肌动蛋白,结合蛋白以及细胞内内质网腔的氧化还原酶(ERp57)等表达水平下调。肌动蛋白是真核生物中最丰富的蛋白质之一,分布于整个细胞中,能通过聚合形成微丝。肌动蛋白形成的微丝(F-肌动蛋白)是真核细胞骨架重要组成部分。在绝大多数真核细胞中,F-肌动蛋白形成大规模的网络结构,这种结构对于细胞器的运动、形状提供机械支持,并提供通过细胞质运输的转导信号等诸多功能[27],除此以外,对细胞的凋亡、迁移和增殖等生物学功能也有着重要的影响。结合蛋白(Bip)在未折叠蛋白反应信号传导通路中处于核心地位,被认为是未折叠蛋白反应的标志性分子,具有帮助错误折叠的蛋白恢复成正确折叠方式的作用。已有研究表明内质网中的蛋白质功能主要是监控蛋白质折叠是否正确,终末错折叠的蛋白质与结合蛋白结合[28],并通过蛋白酶体降解,结合蛋白表达水平的下调说明精子塞形成过程中,蛋白质正确折叠,蛋白质错误折叠率下降。已有报道证实ERp57 是一种存在于内质网中的分子伴侣,具有多种生物学功能,它能与不同的蛋白质折叠中间体相互作用,帮助多肽链的正确折叠和装配[29]。在小鼠精子中,已证实ERp57 定位于精子赤道部,在精卵融合过程中起着重要的作用。2006年,ELLERMAN,et al[30]报道ERp57 定位于小鼠顶体反应后精子的赤道区,与精卵融合有关。随后在人类精子的研究中也发现,ERp57 主要定位于精子的顶体和尾部,顶体反应后位于赤道部。猜测ERp57 有可能与三疣梭子蟹交配过程的精子顶体反应有关,ERp57 在精子塞形成过程中发挥重要作用。由此可知精子塞形成的关键蛋白为结合蛋白与肌动蛋白等,且与ERp57 存在密切联系。通过三疣梭子蟹精子塞表达差异分析,差异蛋白在生物学过程、细胞组成以及分子功能均出现表达水平下调,且没有表达水平上调,说明在精子塞形成过程中:生物调节,细胞过程,代谢过程,生物过程调节,应激反应,单生物过程等生物学过程会出现不同程度的增加;结合和催化活性同时也会提高;细胞、细胞区域、大分子复合物、细胞器等细胞组分增多。
猜你喜欢
四川蚕业(2021年1期)2021-02-12
中国生殖健康(2019年2期)2019-08-23
华夏地理(2019年2期)2019-07-24
中国生殖健康(2019年9期)2019-01-07
中国生殖健康(2019年7期)2019-01-06
中国生殖健康(2019年6期)2019-01-06
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
现代营销·经营版(2017年3期)2017-05-17
作文周刊·小学五年级版(2016年1期)2016-06-29
