
湿加松无性系表型遗传多样性研究
2019-03-19 08:54郭文丽李义良赵奋成廖仿炎钟岁英林昌明叶威方
植物研究 2019年2期
郭文丽 李义良 赵奋成 铁 军 廖仿炎钟岁英 林昌明 叶威方
(1.山西师范大学生命科学学院,临汾 041004; 2.广东省森林培育与保护利用重点实验室,广州 510520; 3.广东省林业科学研究院,广州 510520; 4.长治学院生物科学与技术系,长治 046011; 5.广东省台山红岭种子园,台山 529200)
湿加松(Pinuselliottii×P.caribaeaMorelet var.hondurensis)属松科(Pinaceae)松属(Pinus),为裸子植物。湿加松是经控制授粉后得到的杂交子代,其母本为湿地松(PinuselliottiiEngelmann),父本为洪都拉斯加勒比松变种(PinuscaribaeaMorelet var.hondurensis)。湿加松不仅具有母本干型通直圆满、分枝情况良好、抗风抗病虫害能力强、耐水渍、耐短期低温等优势,还具有父本早期生长迅速、生长量大等优势[1]。从上世纪末开始,我国木材市场就出现了紧缺产脂力高和出材量大的松树良种的现象,湿加松依靠独有的优势已成为我国华南地区兼具有建材产脂和大径材林培育潜力的重要树种[2]。
目前,我国湿加松无性系有两个来源:其一为引种自澳大利亚昆士兰州的湿加松,其二是上世纪九十年代以来广东省林业科学研究院与台山红岭种子园等单位联合培育的湿加松[3]。经过二十多年的大量研究,同等立地条件下我国广东省培育的部分湿加松优良家系在材积生长方面上优于澳大利亚F1代家系。
前人在研究不同植物表型性状普遍采用了主成分分析和聚类分析的方法,例如豌豆[4]、茶树[5~6]、谷子[7]、陆地棉[8]、甜高粱[9~10]、杜鹃红山茶[11]等作物已经进行了相关研究。本研究拟选取广东省台山市红岭种子园35个湿加松无性系作为试验材料,测量其8个表型性状,采用主成分分析和聚类分析相结合,综合分析湿加松无性系表型遗传多样性,为更高效的选择亲本提供理论依据,进一步为良种选育奠定基础。
1 试验材料及方法
1.1 试验材料、方法
湿加松无性系均来自广东省台山市红岭种子园,试验于2017年8月25日~2017年9月14日在红岭种子园进行。该种子园营建于2000~2003年,总面积18.5 hm2,分为3个大区39个小区,建园无性系共51个,每个小区16个无性系,无性系配置采用了三位顺序错位排列法,株行距为5 m×5 m。建园无性系均选自早期营建的湿加松遗传测定林,从测定林中选择生长量大、干形通直、分枝良好、结实正常、树体健康的优良单株,采集其穗条带回苗圃中嫁接、繁殖,由此组成建园材料。该地属南亚热带海洋性气候,年均温21.8℃,年降水量1 940 mm;平均海拔30 m,土壤为花岗岩发育的酸性砖红壤,pH值5.0~5.5,土层深厚。自然条件下适合湿加松生长发育、开花结实。本试验选定35个无性系,包括10个来自澳洲全同胞家系(EHA),2个来自澳洲半同胞家系F2(EHAF2),23个来自台山自配全同胞家系,每个无性系从种子园随机选取3~6分株不等,共160分株。试验材料详细信息见表1。
1.2 表型性状调查
设计对35个湿加松无性系从生长性状和形质性状两个方面调查,生长性状包括树高、胸径、材积;形质性状包括冠幅、分枝角、侧枝直径、轮枝距、皮厚。
树高:用测高仪(VertexⅣ超声波树木测高仪)测量选定湿加松的树高(单位:m)。
胸径:用胸径尺测量选定湿加松的胸径(1.3 m处)(单位:cm)。
材积:利用树高、胸径计算选定湿加松的材积(单位:m3,单株材积:V)的计算公式[12]如下:
V=(f×π×H×D2)/40 000
(1)
式中,f为形数,取f=0.5,π为圆周率,H为树高,D为胸径。
冠幅:先测量选定材料的东西、南北向冠幅,再计算平均值(单位:m)。
表135个湿加松无性系材料名称表
Table135clonesofP.elliottii×P.caribaeaMoreletvar.hondurensismaterialnametable
选3个有代表性的侧枝,距地面3~5 m,分别测量分枝角、侧枝直径、轮枝距。
分枝角:测量选定的3个侧枝的分枝角(单位:°)。
侧枝直径:用胸径尺测量选定的3个侧枝直径,再计算平均值(单位:cm)。
轮枝距:分别测量选定的3个侧枝之间距离,再计算平均值(单位:cm)。
皮厚:位于树高1.3 m处测量选定材料的树皮厚度(单位:cm)。
1.3 数据处理及统计分析
本研究采用R软件[13]对35个湿加松无性系的表型数据进行分析,使用summary()函数计算各表型性状的最小值、最大值、平均值,使用stat.desc()函数计算了标准误、方差、变异系数,以及进行了多样性指数、表型相关性分析、主成分和聚类分析。
遗传多样性指数采用Shannon-Weaver多样性指数(H),其计算公式为:
H=-∑PilnPi
(2)
式中:H为任一组中某一性状的多样性程度;P为某一性状第i个级别的种质份数占总种质材料的百分比;ln表示自然对数。
其中表型性状的相关性分析是间接选择的重要参数,记为rtt。计算公式如下:
(3)
式中:Covt1,t2类是两性状的协方差;Vt1、Vt2是两性状的遗传方差。
主成分分析采用降维把多维空间多个相关变量简化为少量几个相互独立的综合指标,可以更好地反映出各指标间的综合情况[14]。本研究使用psych程序包对35个湿加松无性系进行了主成分分析。
聚类分析采用了系统聚类法(层次聚类法),利用函数hclust()对35个湿加松无性系进行聚类分析。
2 结果与分析
2.1 35个湿加松无性系的表型性状差异
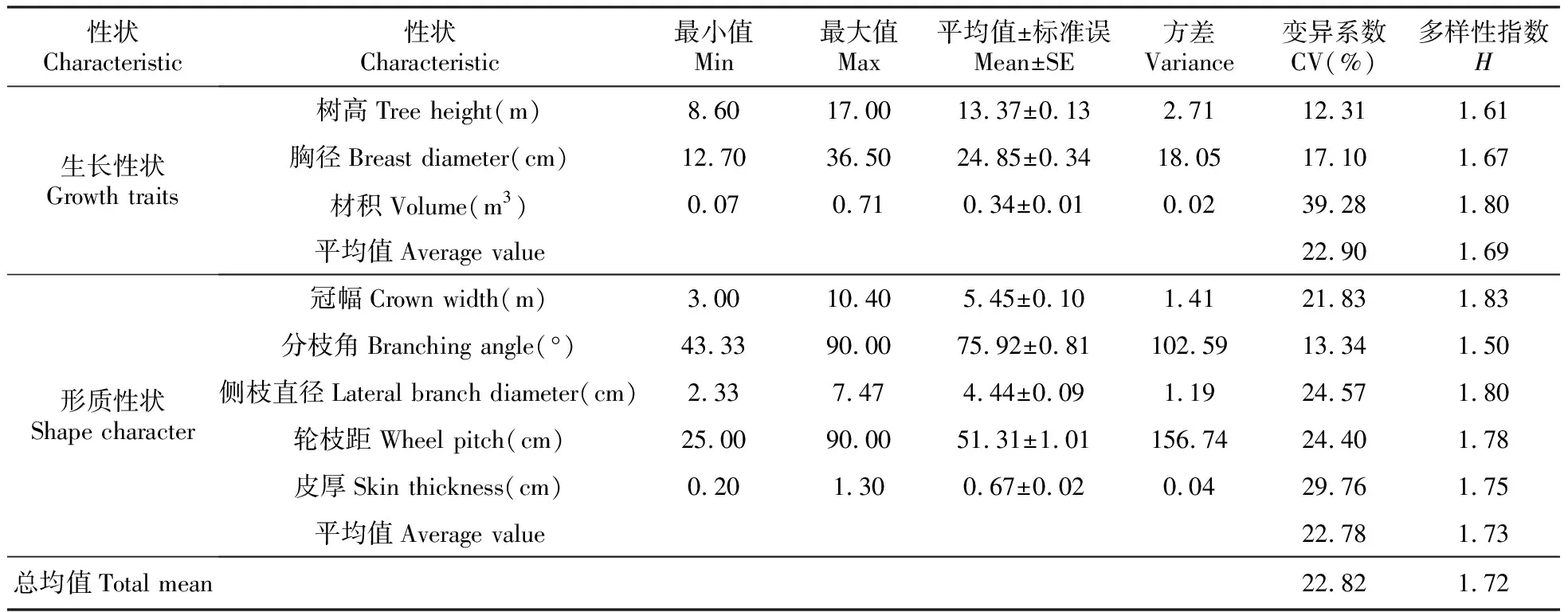
对35个湿加松无性系的8个表型性状进行了描述性统计分析,分析结果见表2。
表2可知,湿加松树高变幅8.60~17.00 m,平均值为13.37 m;胸径变幅12.70~36.50 cm,平均值为24.85 cm;材积变幅0.07~0.71 m3,平均值为0.34 m3;冠幅变幅3.00~10.40 m,平均值为5.45 m;分枝角变幅43.33~90.00°,平均值为75.92°;侧枝直径变幅2.33~7.74 cm,平均值为4.44 cm;轮枝距变幅25.00~90.00 cm,平均值为51.31 cm;皮厚变幅0.20~1.30 cm,平均值为0.67 cm。
8个表型性状变异范围为12.31%~39.28%,均值为22.82%,其中生长性状介于12.31~29.28%,均值为22.90%,形质性状介于13.34~47.25%,均值为26.86%。其变异系数由大到小依次为:材积>皮厚>侧枝直径>轮枝距>冠幅>胸径> 分枝角>树高。依据张岩等[15]可将变异系数分为三级,小(<25%)、中(25~50%)、大(>50%)。本研究只有小、中两级。其中,材积的变异系数最大,呈现了丰富的变异性;树高和分枝角的变异系数均小于15%,说明其遗传稳定性较好。
表235个湿加松无性系8个表型性状的描述性统计及遗传多样性
Table2Descriptivestatisticsandgeneticdiversityofeightphenotypictraitsin35clonesofP.elliottii×P.caribaeaMoreletvar.hondurensis
性状Characteristic性状Characteristic最小值Min最大值Max平均值±标准误Mean±SE方差Variance变异系数CV(%)多样性指数H生长性状Growth traits树高Tree height(m)8.6017.0013.37±0.132.7112.311.61胸径Breast diameter(cm)12.7036.5024.85±0.3418.0517.101.67材积Volume(m3)0.070.710.34±0.010.0239.281.80平均值Average value22.901.69形质性状Shape character冠幅Crown width(m)3.0010.405.45±0.101.4121.831.83分枝角Branching angle(°)43.3390.0075.92±0.81102.5913.341.50侧枝直径Lateral branch diameter(cm)2.337.474.44±0.091.1924.571.80轮枝距Wheel pitch(cm)25.0090.0051.31±1.01156.7424.401.78皮厚Skin thickness(cm)0.201.300.67±0.020.0429.761.75平均值Average value22.781.73总均值Total mean22.821.72
2.2 35个湿加松无性系表型性状的遗传多样性
对供试无性系的8个性状进行了遗传多样性分析(表2)。从表2可知,8个性状多样性指数变化范围在1.50~1.83,均值为1.72,生长性状介于1.61~1.80,均值为1.69,生长性状介于1.50~1.94,均值为1.77。其遗传多样性指数由高到低依次为:冠幅>材积>侧枝直径>轮枝距>皮厚>胸径>树高>分枝角,表明了表型性状的遗传多样性均较丰富。
2.3 35个湿加松无性系表型性状的相关性分析
对8对不同表型性状进行相关性分析(表3)。从表3可知,各性状之间存在着不同程度的相关性,且大部分呈显著或极显著相关。其中呈极显著正相关的有11对,依次为材积与皮厚(0.98)、冠幅与侧枝直径(0.94)、树高与皮厚(0.92)、树高与材积(0.89)、材积与侧枝直径(0.82)、树高与胸径(0.81)、材积与冠幅(0.72)、胸径与侧枝直径(0.71)、胸径与冠幅(0.69)、树高与轮枝距(0.62)、树高与侧枝直径(0.46);呈显著正相关的有5对,依次为轮枝距与皮厚(0.72)、胸径与轮枝距(0.63)、侧枝直径与皮厚(0.55)、材积与轮枝距(0.51)、树高与冠幅(0.36)。表明供试材料材积较高且皮较厚、冠幅与侧枝直径较大、树体高大以及轮枝距较大;皮厚与材积、树高呈极显著相关,与轮枝距、侧枝直径呈显著正相关,因此对供试材料的育种选择中应以材积为主要指标,其次为树高。
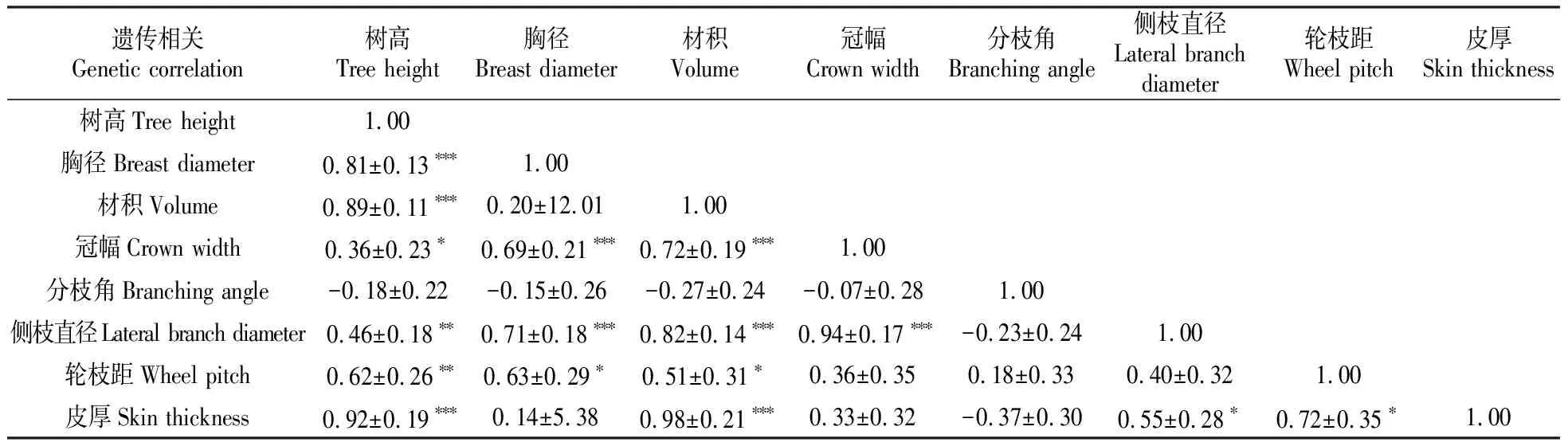
表3 8个表型性状之间的相关性分析
注:***,**和*为分别表示在0.001、0.01和0.05水平上存在显著相关。
Note:***,** and * indicate significant correlations at the levels of 0.001,0.01 and 0.05,respectively.
表4湿加松无性系各类群表型性状的特征
Table4TheaverageandcoefficientvariationamountofphenotypictraitsindifferentgermplasmgroupsofP.elliottii×P.caribaeaMoreletvar.hondurensis

性状Traits资源数Number树高Tree height(m)胸径Breast diameter(cm)材积Volume(m3)冠幅Crown width(m)分枝角Branching angle(°)侧枝直径Lateral branch diameter(cm)轮枝距Wheel pitch(cm)皮厚Skin thickness(cm)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)均值Mean变异系数CV(%)种质类群Germplasms groupsⅠ213.080.0424.090.050.320.114.730.0957.710.044.150.3346.690.050.690.41Ⅱ314.521.4526.811.280.423.115.820.6669.24138.894.540.0165.45500.790.005Ⅲ113.830.0226.471.280.380.0015.472.658034.723.510.0023412.50.670.005Ⅳ1313.1224.0610.580.320.0055.540.1573.512.54.424.9146.0912.50.640.02Ⅴ1213.720.7226.23320.380.025.660.00179.27868.064.852.4255.866.130.70.005Ⅵ412.811.2823.4280.650.270.054.851.8179.71312.53.80.6855.4100.620.005
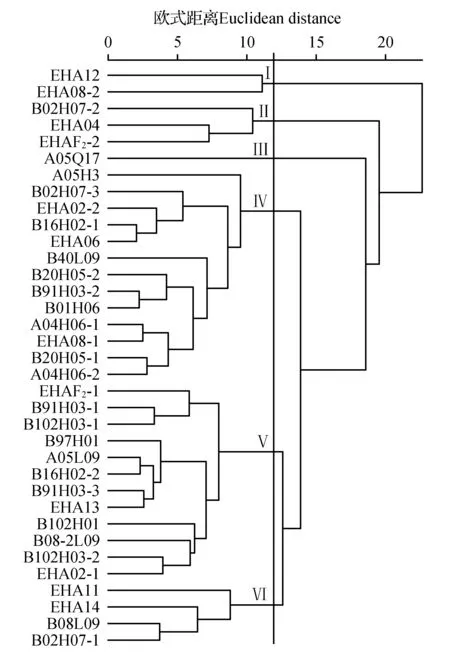
图1 35个湿加松无性系的系统聚类图Fig.1 Hierarchical Cluster Map of 35 Clones of P.elliottii×P.caribaea Morelet var. hondurensis
2.4 聚类分析
采用R软件对35个湿加松无性系进行了系统聚类分析(图1)。在欧氏距离为12时,可将35个无性系分为6大类群,每个类群的份数、表型性状的平均值以及变异系数(表4)。
第Ⅰ类:有2个无性系,均来源于澳洲全同胞家系,共8分株,占供试材料总数的5%。主要表现为冠幅较小,可为培育窄冠型良种提供种质。冠幅和分枝角(<60°)的均值在六大类群里均最小,变异系数均较小。
第Ⅱ类:有3个无性系,1个来源于台山全同胞家系、1个来源于澳洲全同胞家系、1个来源于澳洲半同胞家系F2,共13分株,占总材料的8.125%。主要表现为树体高大,树冠宽,粗皮且材积大。树高、胸径、材积、冠幅、轮枝距和皮厚的均值在六大类群里均最大,且材积的变异系数最大。综合分析可知,该类群长势最优,可为培育速生良种提供种质材料,有较高的用材经济价值。
第Ⅲ类:只有1个无性系,来源于台山全同胞家系,该无性系长势良好,但分枝角的均值最大(80°),侧枝直径与轮枝距的均值最小,变异系数都较低,仅3分株,占总材料的1.875%。
第Ⅳ类:共13个无性系,分别有3个来源于澳洲全同胞家系、10个均来源于台山全同胞家系,共64分株,占总材料的40%。其整体长势较差,且树高和侧枝直径的变异系数均最大,需进一步培育从中选取优良品种。
第Ⅴ类:包括了12个无性系,有1个来源于澳洲半同胞家系F2、2个来源于澳洲全同胞家系、9个来源于台山全同胞家系,共55分株,占总材料的34.375%。其侧枝直径的均值最大,分枝角及轮枝距的变异系数最大,说明选择范围较宽,整体上的长势优于第Ⅳ类,适合后期大量种植。
第Ⅵ类:包括4个无性系,2个来源于澳洲全同胞家系、2个来源于台山全同胞家系,共17分株占总材料的10.625%。其树高、材积与皮厚的均值最小,胸径的变异系数最大,说明该类群长势最差,但皮薄,胸径选择范围较宽,可为培育薄皮型良种提供种质。
2.5 主成分分析
本研究针对8个性状采用R软件的程序包psych进行了主成分分析(表5)。结果表明,前3个主成分的特征值均大于1,固选为主成分,3个主成分的累积贡献率达86.00%,基本上可表现6个性状的综合变异情况。第1主成分贡献率为39.36%,树高、胸径和皮厚的系数明显高于其它性状,说明了湿加松表型遗传多样性的重要变异因子为树高、胸径和皮厚三个表型性状;第2主成分贡献率为30.32%,冠幅、侧枝直径的特征向量值均较高,说明第2主成分是冠幅、侧枝直径的综合反映;第3主成分贡献率为16.33%,系数最大的是分枝角(0.92),说明第3主成分主要反映了分枝角因子。
表535个湿加松无性系基于8个表型性状的5个主成分的特征向量和贡献率
Table5Eigenvectorandcontributionrateoffiveprincipalcomponentsbasedoneightphenotypictraitsin35clonesofP.elliottii×P.caribaeaMoreletvar.hondurensis

表型性状Phenotypic traits主成分Principal componentRC1RC2RC3树高Tree height0.790.19-0.11胸径Breast diameter0.70.50.05材积Volume0.690.58-0.11冠幅Crown width0.130.850.12分枝角Branching angle-0.12-0.020.92侧枝直径Lateral branch diameter0.160.85-0.15轮枝距Wheel pitch0.63-0.010.42皮厚Skin thickness0.80.11-0.19特征值Characteristic value2.72.081.12方差贡献率Variance contribution rate(%)39.3630.3216.33累积方差贡献率Cumulative variance contribution rate(%)39.3669.6786.00
注:下划线表示每个主成分较高的特征向量。
Note:Underline represents the higher eigenvector for each principal component.
3 讨论
3.1 湿加松无性系的遗传多样性分析
遗传多样性作为种质资源的重要评价因子,也是培育优良品种的研究提供重要依据[16]。调查表型性状的遗传多样性是衡量表型变异最直观最简易的途径,但易受环境、个体差异或标记性状较少等诸多外在因素影响。前人在表型性状对松属植物遗传多样性上的研究[17]较少,仅在贺兰山杜松(Juniperusrigida)[18]、欧洲赤松(PinussylvestrisL.)[19]等少数松树上有过相关报道,大多采用RFLP标记[20]、RAPD标记[21]、SSR标记[22~24]、ISSR[25]等分子标记法研究松树遗传变异规律。本文通过对35个湿加松无性系的8个表型性状进行遗传多样性分析,结果发现8个表型性状的平均遗传多样性指数较高,表明其有着较高的丰富度与均匀度,丰富了我国湿加松无性系的遗传背景。另外,从计算结果可以看出大部分性状的变异系数和遗传多样性基本统一,材积均较高,分枝角和树高均较低,说明其离散程度相似。影响湿加松变异的有遗传与非遗传两大因素,各占一半,但研究表明家系的不同对遗传影响最强,其次是无性系的不同[26],所以该研究的遗传多样性分析结果可能受到了不同来源的无性系影响。
3.2 湿加松无性系的相关性分析
通过相关性分析,9对表型性状之间存在着较高的相关性,其中有11对极显著正相关,5对显著正相关。研究表明树体越高,皮较厚,胸径、材积、侧枝直径、轮枝距也较大,这与梁德洋等[27]对红松无性系的研究结果相似,树高、胸径与材积之间均呈极显著正相关。李义良等[12]研究表明湿地松、加勒比松的树高、胸径、材积、皮厚及冠幅两两间均呈极显著正相关,本研究中除胸径与材积、胸径与皮厚及冠幅与皮厚均呈不显著相关其它性状也呈显著正相关。综上所述,在选育湿加松时应优先参考材积为第一指标,同时应参考树高、皮厚、胸径、冠幅及侧枝直径等综合指标,为今后的选育工作给予理论指导。本研究中参试无性系数量不同可能对试验结果产生一定的误差,后期应增加材料的数量,另外应增加产脂力、木材密度、种子量等性状的收集,进一步分析更具说服力[28]。此外,用R软件进行相关性分析要比IBM SPSS Statisticsr类似软件更为直观,其中极显著相关分成0~0.001和0.001~0.01两个级别,更有利于最优指标的提取。
3.3 湿加松无性系的聚类分析
利用湿加松无性系的8个表型性状相关数据进行系统聚类分析,将表型差异较小的分为一个类群,把35个无性系分为6大类群。第Ⅰ类为选育窄冠型优良基因的材料;第Ⅱ类为后期选育速生良种的材料,木材经济价值较高;第Ⅲ类虽长势良好,但分枝角较大,应缩短生长周期;第Ⅳ类长势均较差,需进一步具体分析再选育良种;第Ⅴ类长势优,为后期选优的主要良种材料;第Ⅵ类为选育薄皮型良种的材料。通过对各类群的具体分析,部分来源相同的聚在一类,如来源于澳洲全同胞家系的EHA12与EHA08-2、EHA11与EHA14分别聚为一类,来源于台山全同胞家系的父本为H01与H03、H05与H06的大多聚在一类,说明它们在表型上的变异较小。大部分来源相同的种质聚在不同类群,说明无性系在表型上存在了较大的差异,与无性系的来源没有必然的联系,这与万述伟等[4]和王丽侠等[29]的研究结果相同。在育种时可依据表型性状的聚类结果,同时结合分子标记法,去合理分配种质,可有效避免近交,减少控制授粉的工作,大大缩短育种周期,提高湿加松无性系的改良效率。
3.4 湿加松无性系的主成分分析
本研究对湿加松的8个表型性状进行主成分分析,提取了3个主成分,反映了6个性状的大部分信息,累积贡献率高达86.00%,与相关性分析结果相似。第1主成分主要代表了树高、胸径和皮厚三个生长性状的综合情况,体现出湿加松在树高、胸径、皮厚三方面明显的杂种优势,这与李义良等[30]对湿加松杂种优势的研究结果相似。此外,第2主成分主要反映了冠幅、侧枝直径的综合情况;第3主成分主要反映了分枝角的情况,所以这些性状也可评价湿加松的优劣。
综上所述,对不同来源湿加松无性系的表型性状进行分析,初步了解了湿加松种质的表型遗传多样性情况,为今后种质利用及良种选育奠定了理论基础。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29
广西林业科学(2022年3期)2022-07-08
绿色科技(2022年11期)2022-07-02
绿色天府(2022年2期)2022-03-16
林业科技情报(2021年3期)2021-09-01
山东林业科技(2020年1期)2020-05-18
蔬菜(2019年9期)2019-01-05
农村农业农民·B版(2018年2期)2018-03-22
绿色科技(2016年23期)2017-03-15
卷宗(2012年5期)2012-10-21
