
应用神经突方向分散度和密度成像技术研究rs747302基因单核苷酸多态性对青春期儿童脑白质微结构的影响
2019-02-14 09:43李霄扬黄明珠张旭于兵
磁共振成像 2019年11期
李霄扬,黄明珠,张旭,于兵
作者单位:中国医科大学附属盛京医院放射科,沈阳,110001
近年来的一系列研究发现,多巴胺受体D4基因启动子区-616 位点(rs747302)的单核苷酸多态性(single nucleotide polymorphism,SNP)可能与多种儿童精神异常,如注意缺陷多动障碍,原发性夜间遗尿症[1-2]等的发病具有相关性,但是rs747302基因多态性对正常青春期儿童脑微结构,特别是白质微结构的研究,截至目前未见相关报道。
扩散磁共振成像技术通过施加不同参数的梯度脉冲,以测量微米数量级的水分子扩散差异,从而计算出细胞完整性和组织微结构等信息,是目前唯一有能力在活体无创地检测脑白质微结构的影像学手段。扩散张量成像是目前最经典且临床应用最广泛的技术,它对于脑白质微组织结构异常具有较好的敏感性,但是特异性不够强,脑白质微观结构的某些特性有时会被掩盖,例如,扩散张量成像反映的微结构的各向异性差异往往被方向分散所掩盖。只有区分出这些特性,才能够对树突和轴突的微结构进行深入的无创性的探索。
Zhang等[3]基于磁共振扩散成像扫描开发的神经突方向分散度和密度成像(neurite orientation dispersionand density imaging,NODDI)技术,为临床实现神经突(树突和轴突)的微结构检测提供了全新的渠道。因而,本研究计划使用NODDI技术,探索rs747302基因对正常青春期儿童脑白质微结构的影响。
1 材料与方法
1.1 研究对象
本研究根据rs747302基因型检测结果以及患者的性别、年龄状况,在2016年1月至2018年12月期间,选取182名年龄11~13岁,神经系统发育正常的青春期儿童。该研究计划已经获得我院伦理委员会审查批准(批准号2015PS54),所有参与研究的被试者本人及其父母(父母不具备民事行为能力时由其法定监护人代理)均签署了相应的知情同意书。
所有参与研究的青春期儿童入组标准如下:神经和精神系统问诊及查体正常,无血液系统、神经系统疾病及精神疾病病史;常规MRI检查脑实质未见异常;无睡眠呼吸异常病史;无典型精神及精神药物服用史,由具备资质的心理科医师进行韦氏智力测验后确认智商(intelligence quotient,IQ)>8。
1.2 数据采集
1.2.1 rs747302基因型分析
所有受试儿童常规采集5 ml外周静脉血,并采用EDTA抗凝,用碘化钾法从外周血淋巴细胞中提取基因组DNA用于后续检测。rs747302 SNP分析采用PCR技术,使用 ABI 9700 PCR扩增仪检测rs747302基因C/G多态性,PCR反应的引物设计及PCR反应条件详见表1。由于人群中rs747302基因的CC纯合子分布频率很低,因而本研究中将CC和CG基因型的被试合并为C等位基因携带组(C组),与G纯合子被试儿童(G组)进行组间对比。
1.2.2 韦氏智力测验
韦氏智力测验采用湖南医科大学龚耀先等修订的韦氏儿童智力量表(C-WISC)[4]。所有受试者均完成11项分测验,其中言语分测验6项包括知识(I)、领悟(C)、分类(S)、算术(A)、数字广度(D)、词汇(V);操作分测验5项包括填图(PC)、图片排列(PA)、木块图(BD)、图形拼凑(OA)、编码(Cd)。从分测验中分别获得言语量表分(V)、操作量表分(P)及总量表分(F)。进一步计算言语智商(VIQ)、操作智商(PIQ)、总智商(IQ),3个因子智商:言语理解因子(VC)=I+V+C+S、知觉组织因子(PO)=PC+PA+BD+OA、记忆/不分心因子(M/C)=A+D+Cd。
1.2.3 磁共振扫描数据处理
NODDI扫描采用Philips Ingenia 3.0 T超导型MRI扫描仪,使用64通道头线圈进行扫描。NODDI 扫描采用多层同时成像(Multi-Band SENSE)自旋平面回波(SEEPI)序列,扫描参数:TR 4000 ms,TE 85 ms,反转角90°,SENSE加速因子为P=2.5,Multi-Band加速因子为2,FOV 220 mm×220 mm,扫描矩阵110,层厚2 mm,层间距0 mm,层数54层,b值为0、1000 s/mm2及2000 s/mm2,每个b值下扩散敏感梯度方向32个。NODDI序列扫描时间为7 min。
从MRI扫描仪导出NODDI扫描的原始图像,在由专业的神经组放射科医师目视观察去除形变和头动明显的被试数据后,首先应用FSL (https://fsl.fmrib.ox.ac.uk/fsl/)软件包对NODDI数据进行涡流头动校正和剥脑预处理,然后应用伦敦大学Hui Zhang教授团队编写,的NODDI Matlab Toolbox (V0.9)(http://mig.cs.ucl.ac.uk/index.php?n=Download.NODDI)软件包,计算得到突触体积分数(neurite density index,NDI)、神经突方向分散系数(orientation dispersion index,ODI)等NODDI指标图。
为进一步计算白质NODDI指标的组间差异,本研究采用FSL基于纤维束示踪的空间统计分析(Tract-Based Spatial Statistics,TBSS)组件,对NODDI指标图(NDI和ODI图)进行深入分析,具体步骤如下:首先应用FSL DTIFIT组件,根据预处理后的NODDI数据中的b=700s/mm2,32个弥散梯度方向的扫描数据,计算每名被试的部分各向异性图;然后应用FMRIB Non-linear Registration Tool工具包,将各向异性图配准到蒙特利尔神经研究所(Montreal neurological institute,MNI)空间;之后应用TBSS软件包根据配准后的部分各向异性图得到平均部分各向异性图纤维骨架图像;最后,将所有被试的NODDI指标图(NDI和ODI图)映射到平均部分各向异性图纤维骨架上,得到每个被试的ODI纤维骨架图和ODI纤维骨架图。
1.2.4 统计学分析
应用SPSS 17.0软件包(SPSS Inc.,Chicago,IL,U S A)对被试数据进行分析。首先采用Kolmogorov–Smirnov检验对于所有的定量数据进行正态性分布检验,符合正态分布的数据用均值±标准差表示,不符合正态分布的数据则以中位数(四分位间距)表示。采用Student’st检验或Mann-Whitney U检验对于年龄、受教育程度进行各组间比较,采用Chi-square检验比较性别的各组间差异,以P<0.05作为组间差异具有统计学意义。
由于韦氏智力测验的部分结果为非正态分布,所以我们采用Mann-Whitney U检验对智力测验的结果进行组间比较,并采用Bonferroni法进行多重比较校正,以校正后P<0.05作为差异有统计学意义。
为了探索不同基因型被试白质微结构的组间差异,本研究应用FSL的Randomize组件,将性别、年龄受教育程度和IQ作为协变量,采用置换检验(置换次数5000次)分别对于NDI和ODI的纤维骨架图进行基于纤维束示踪的组间分析,组间分析结果采用参考簇水平(threshold-free cluster enhancement,TFCE)方式进行多重比较校正,将校正后P<0.05的白质区域作为组间差异显著。根据约翰霍普金斯大学ICBM-DTI-81白质纤维图谱定位组间差异显著的白质纤维束。
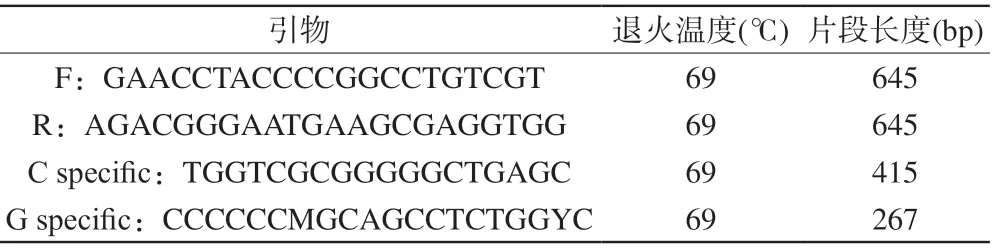
表1 rs747302基因型分析引物及反应条件Tab. 1 Protocol of rs747302 genotyping

表2 所有被试DRD4 -616位点基因检测结果Tab. 2 The results of genotyping of DRD4 -616 C/G SNP
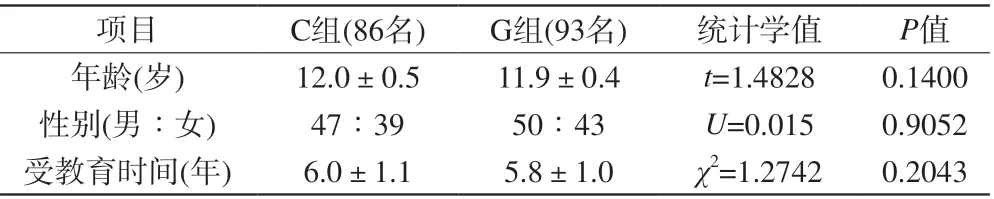
表3 所有被试基本情况Tab. 3 Demographic data of the participants
为了深入分析NODDI指标的组间比较结果,我们分别将NDI和ODI组间差异显著的白质区域作为感兴趣区,提取所有被试感兴趣区内的NODDI指标进行组间比较,并且采用Spearman相关分析的方法,探索感兴趣区内NODDI指标与韦氏智力测验的相关性。
2 结果
4名被试儿童的NODDI影像数据形变或头动明显,其基因型数据和影像数据被排除。列入统计的被试基因检测结果见表2,他们的年龄和接受教育时间未见组间差异,但是C等位基因携带者的记忆/不分心因子显著低于G纯合组,详见表3、4。
基于TBSS组间分析结果显示,rs747302基因C/G SNP导致的被试白质组间差异主要位于双侧丘脑前辐射(anterior thalamic radiation,ATR),表现为ODI的升高及NDI的减低(P<0.05,TFCEcorrected)(图1)。分别将NDI和ODI组间差异显著的ATR白质区域作为感兴趣区,提取所有被试感兴趣区内的NODDI指标进行组间比较,发现C等位基因携带者的ATR的ODI值显著高于G纯合组,而且NDI显著低于G纯合组(U=120,P<0.001;U=890,P<0.001)。Spearman相关分析的结果显示,C等位基因携带者的NDI值与记忆/不分心因子正相关(r=0.6804,P<0.001) (图2)。

表4 韦氏智力测验结果Tab. 4 Intelligence testing results of the participants

图1 rs747302基因多态性对白质NODDI参数的影响 图2 rs747302对ATR NODDI参数的影响极其与注意/不分心因子的联系 Fig. 1 Effect of rs747302 gene on NODDI parameters of white matter. Fig. 2 Effect of rs747302 gene on NODDI parameters of ATR and its relationship with M/C factor.
基于纤维束示踪的组间分析结果显示,rs747302基因C/G SNP导致的被试白质组间差异主要位于双侧丘脑前辐射(anterior thalamic radiation,ATR),表现为ODI的升高及NDI的减低。
组间分析结果显示:(A) C等位基因携带者ATR的NDI值显著低于G纯合组;(B) C等位基因携带者ATR ODI显著高于G纯合组;(C) C等位基因携带者记忆/不分心因子显著低于G纯合组;(D) C等位基因携带者的NDI值与记忆/不分心因子正相关(r=0.6804;P<0.001)。
3 讨论
NODDI建立了一个可以区分细胞内间隔、细胞外间隔和脑脊液等三种微结构环境的组织结构模型:细胞内间隔指神经突膜环绕的空间。在脑组织内,细胞内间隔模拟的神经方向的范围包括方向高度一致的白质结构(如胼胝体)、弯曲或者呈辐射状的白质结构(如半卵圆中心)和向各个方向蔓延为特征的皮层和皮层下灰质结构;细胞外间隔)细胞外间隔是指神经周围的空间,它被各种类型胶质细胞和灰质中的细胞体所占据。在这个空间里,水分子的扩散被神经突起所阻碍,而不是仅仅受限。因此,用简单的各向异性扩散模拟细胞外间隔。CSF(脑脊液)是指脑脊液所占据的空间。这个空间用各向同性的符合高斯分布的扩散来描述。现有的研究表明,相对于传统的扩散张量成像模型,NODDI模型于白质的微结构有着更高的灵敏性和特异性[5]。
rs747302基因位于DRD4基因启动子区, 存在转录调控蛋白AP-2的结合位点,而该位点C、G等位基因的多态性[6],可能影响AP22结合位点的活性,从而影响了DRD4基因的诱导及转录,进而造成一些DRD4蛋白结构与功能的变化[4,7]。
本研究结果显示,rs747302基因主要造成了双侧ATR的白质微结构改变,而且ATR的NDI改变与C等位基因携带者的注意/不分心因子相关。ATR结构主要连接丘脑以及额叶,是额叶-丘脑神经环路的重要联络纤维,而额叶-丘脑神经环路对于注意力的维持、睡眠/觉醒状态的切换以及排尿行为的控制都起着重要的作用[8]。考虑已有研究证实ADHD和遗尿症的额叶-丘脑结构连接及功能连接均存在异常[9-11],笔者推测C等位基因携带者的ATR微结构改变可能造成了他们ADHD和遗尿症发病的风险相对升高;考虑到ATR的NDI改变与C等位基因携带者的注意/不分心因子减低具有相关性,可能表明rs747302基因的多态性,与ADHD患者的注意力不集中以及以前研究中发现的PNE患儿注意缺陷可能存在一定的关联性[12]。
本研究还表明C等位基因携带者的ATR ODI值升高且NDI减低,这表明C等位基因携带者的ATR部分区域神经突总体积减小,而且神经突的排列比较分散。基于以往的动物实验研究结果,笔者推测rs747302基因的C等位基因影响了DRD4蛋白的表达,从而干扰了脑发育过程中突触连接修剪的过程,进而影响了C等位基因携带者ATR白质的神经突微结构,造成了他们ATR的部分区域白质神经突减少,由于ATR神经突的总体积减小,所以这些神经突在相对大的空间内可能分布排列也较G组分散[6]。
本研究还存在较多局限性,首先,因为样本量及C等位基因比率所限,我们在研究中对CC和CG基因型的合并可能造成一定程度的遗传效应混淆;其次,考虑到青春期儿童的生长发育变化比较剧烈,我们将被试的年龄限制在较小的范围内,因此我们还需要在以后对更大年龄范围内进行深入研究和长期随访,以验证rs747302基因SNP对于不同生长发育阶段儿童脑白质微结构的影响。
利益冲突:无。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
中老年保健(2021年12期)2021-08-24
昆明医科大学学报(2021年3期)2021-07-22
陶瓷学报(2021年2期)2021-07-21
中西医结合肝病杂志(2020年2期)2020-10-27
医学新知(2019年4期)2020-01-02
制造技术与机床(2019年6期)2019-06-25
制造技术与机床(2018年11期)2018-11-23
故事作文·高年级(2017年3期)2017-04-12
中国运动医学杂志(2016年3期)2016-07-10
