
叶色退化的紫叶狼尾草生理变化特征
2019-01-25 04:50章陆杨许志敏刘燕珍丁国昌
福建农林大学学报(自然科学版) 2019年1期
朱 婷, 章陆杨, 许志敏, 郝 亮, 陈 琳, 刘燕珍, 丁国昌
(福建农林大学园艺学院,福建 福州 350000)
颜色对观赏价值的影响较为明显[1].彩叶植物的叶色表现是遗传因素和外部环境共同作用的结果,色素种类、比例和分布是导致叶色变化的直接原因[2].影响叶片呈色的外部因素主要包括光照、温度、水分等,其中光照强度是影响彩叶植物叶色变化最重要的环境条件[3],叶色退化也是彩叶植物的生命现象之一,对植物叶色影响较大.
观赏草是一类株形优美、色彩丰富的草本植物[4].彩叶观赏草不仅继承了观赏草的优良抗性及独特的形态,并且在观赏色彩上有了进一步提升,此类深红、深紫色叶是最引人注目的色调之一,能够营造出令人难忘的效果[5].紫叶狼尾草(Pennisetumsetaceum‘Rubrum’)是典型的深紫色彩叶观赏草,它是禾本科狼尾草的栽培品种[6].其形态飘逸,整株紫红色,穗状花序轻柔,能保持至晚秋或初冬,极具观赏性,成为人们青睐的园林植物之一.但在实际应用中发现,紫叶狼尾草遮阴状态叶色变绿,且有叶色退化为绿色的现象,影响景观效果.因此,本研究以自然光照、长期遮阴和叶色退化的紫叶狼尾草为对象,测定其叶色参数、色素含量、关键酶等指标,作以对比分析,从生理学角度探讨紫叶狼尾草叶色退化生理机制,为后期研究紫叶狼尾草叶色形成及退化机理提供科学依据,并且为其叶色的分子水平研究提供依据.
1 材料与方法
1.1 试验地概况
试验地设在福建省福州市福建农林大学中华园试验基地(26°5′6″N,119°13′52″),位于福建省东部沿海闽江下游.资源丰富,温暖湿润,四季常青,雨量充沛,夏长冬短,霜少无雪,无霜期达326 d[7].
1.2 试验材料
2018年3月春季,从福州乐彩园艺有限公司引进正常和有叶色退化表现的紫叶狼尾草幼苗.将正常生长的紫叶狼尾草幼苗,一部分进行露天盆栽,作为试验对照组;一部分置于遮光率为30%的大棚内盆栽,作为试验遮阴组,叶色退化的幼苗与对照的紫叶狼尾草同样露天盆栽,管理水平一致.7月初,幼苗成株,且叶色退化的紫叶狼尾草完全变绿(图1),于7月4日早9: 00取样本,选取生长健壮、长势一致的植株,叶片均取自植株由上而下的第3~4片叶,用冰盒带回实验室,贮存于-80 ℃备用,随后进行色差及生理指标的测定.共测定18株,每组6株,6次生物学重复.

CK:对照组;TD:叶色退化;TS:长期遮阴.图1 3组紫叶狼尾草叶色Fig.1 Leaf color of 3 groups of P.setaceum ‘Rubrum’
1.3 测定项目与方法
叶色参数测定参照丁廷发[8]的方法但略有改动.采用国产威福WS70型全自动色差计测定叶片颜色,测色光斑直径为8 mm.选取叶色长势相近的3片功能叶进行测定,每个叶片选取上、中、下3个部位,重复3次,记录叶色参数L,a,b值,其中,L表示光泽明亮度,L值越大,亮度越高;a值表示红/绿,a值越大,红色越深,a值越小,绿色越深;b值表示黄/蓝,b值越大,黄色越深,b值越小,蓝色越深.叶片颜色指数(color index)计算[9]和等级划分[10]参照Carreno et al对葡萄果实颜色的研究方法,等级划分为:绿黄(CIRG<2)、粉红(2
叶绿素a、叶绿素b、类胡萝卜素含量的测定参照张秀君等[11]的方法但略有改动,每样品重复3次,计算参照李合生[12]的方法.花色素苷的测定参照Pirie et al[13]的方法,每样品重复3次,计算参照Xu et al[14]的方法.类黄酮、苯丙氨酸解氨酶( PAL)的检测均采用南京建成生物工程研究所研制的试剂盒进行测定,重复3次.查尔酮异构酶(CHI)采用上海仁捷生物科技有限公司研制的试剂盒进行测定,重复3次.查尔酮异构酶(CHI)的提取参照王惠聪[15]的方法,测定参照Lister[16]的方法,重复3次。
1.4 数据处理
采用Excel 2013软件进行数据整理和绘图.采用SPSS 22.0软件进数据分析,其中运用One-Way ANOVA进行单因素方差分析,用Bivariate Correlations进行相关性分析.
2 结果与分析
2.1 叶色参数
按照Carreno et al[9-10]的方法得出CK为红色级别,TS与TD均为黄绿级别,与肉眼观察结果一致.方差分析结果表明(表1),3组紫叶狼尾草的L值差异显著(P<0.05),TD最大,CK最小,TD分别比CK、TS高33.2%、8.0%.TS、TD两组的a值、b值和叶色参数均无显著性差异.CK的颜色指数远大于TD和TS,分别是TD的2.39倍,TS的2.80倍.可知,a值对叶色的影响最大,长期遮阴和叶色退化的紫叶狼尾草颜色接近.
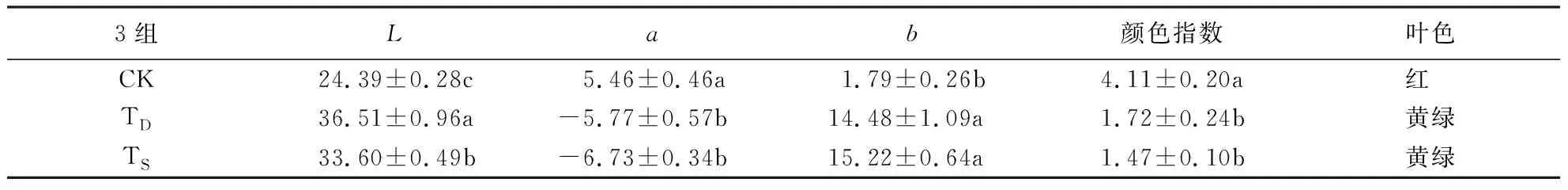
表1 3组紫叶狼尾草叶色参数1)Table 1 Leaf color parameters of P.setaceum ‘Rubrum’
1)表中同列间不同字母表示差异显著(P<0.05).
2.2 光合色素含量
叶绿素和类胡萝卜素是高等植物进行光合作用的主要色素,其中,叶绿素使叶片呈现绿色,类胡萝卜素使叶片呈现橙黄色[2].从图2可知,3组紫叶狼尾草叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量之间均存在显著差异(P<0.05)且变化一致.其中,总叶绿素含量TD比CK、TS分别高36.6%、16.8%,类胡萝卜素含量TD比CK、TS分别高33.2%、22.2%.可见,对照组的光合色素含量最低,长期遮阴和退化的紫叶狼尾草光合色素差异显著(P<0.05)且退化的叶绿素含量最高,两组叶色表现均为绿色但光合色素差异显著,可能是由于叶色退化的紫叶狼尾草处于自然光下,生长状态良好,叶片更加结实饱满,叶肉细胞中含有更多的叶绿体,而导致叶绿素含量更高的结果.
2.3 花色素苷和类黄酮含量及色素含量百分比
花色素苷是高等植物叶片中存在的一种重要的类黄酮色素,与叶绿素、类胡萝卜素共同决定叶片的呈色[17].从图3可知,TS和TD的花色素苷含量无显著差异,与CK间差异显著(P<0.05),CK分别是TD、TS的2.14倍、2.32倍.3组类黄酮含量差异显著(P<0.05),CK最大,TS最小,CK分别是TD、TS的1.97倍、2.70倍.可见,对照组紫叶狼尾草的花色素苷和类黄酮含量都远高于两种叶色变绿的紫叶狼尾草,退化组的花色素苷含量大于长期遮阴且两者无显著差异,这与对叶色参数a值的方差分析结果一致.

CK:对照组;TD:叶色退化;TS:长期遮阴.图中同指标间不同字母表示差异显著(P<0.05).图2 3组紫叶狼尾草光合色素含量Fig.2 Leaf photosynthetic content of 3 groups of P.setaceum ‘Rubrum’
叶色变化的内部因素主要是由光合产物的变化引起植物体内各类色素及其比例的变化,致使叶片呈现不同色彩[18].花色素苷是紫红色叶植物的主要呈色因子[19],因此,花色素苷的占比对于叶片呈现紫红色有较大影响.从图4可知,CK、TD、TS的花色素苷含量分别占27.6%、10.3%、11.5%,CK的含量占比远高于TD、TS.CK、TD、TS的总叶绿素含量分别占53.3%、67.2%、67.4%,类胡萝卜素含量占比分别为19.1%、22.5%、21.1%.可见,对照紫叶狼尾草花色素苷占比大,所以叶片呈紫色,长期遮阴与叶色退化的紫叶狼尾草总叶绿素和类胡萝卜素含量占比大,所以两者呈黄绿色.
2.4 PAL和CHI活性
苯丙氨酸解氨酶(PAL)是花色素苷合成的第1个关键酶[20],而花色素苷是叶色呈现红紫色的关键色素,因此,PAL活性间接决定植物的叶色.PAL也是类黄酮类、木质素等合成过程中起重要调节作用的酶,抵抗外界环境的干扰[21].由方差分析可知(图5),3组紫叶狼尾草的PAL活性均存在显著性差异(P<0.05),CK最大,TS最小,虽然长期遮阴与退化组的紫叶狼尾草叶色相近,但PAL活性存在显著差异(P<0.05).花色素苷和叶色参数的方差分析结果也显示CK最大而TS最小.可见,PAL活性与花色素苷存在紧密联系,叶色越红,PAL活性越高.
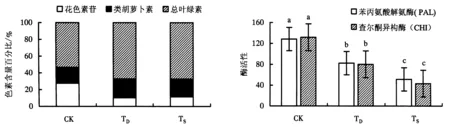
CK:对照组;TD:叶色退化;TS:长期遮阴.图中同指标间不同字母表示差异显著(P<0.05).图4 3组紫叶狼尾草的色素含量百分比Fig.4 Leaf pigment percentage of 3 groups of P.setaceum ‘Rubrum’
有研究表明,CHI酶作为花色素苷合成的关键酶,在黄酮类化合物的转化中起重要作用[22],所以,CHI活性与叶色也可能存在一定的联系.从图5可知,CHI活性与PAL活性变化趋势一致,三组间存在显著差异(P<0.05)。CK组最红,其CHI活性最高,TS组最绿,其CHI活性最低。推断出CHI活性可能在紫叶狼尾草类黄酮化合物转化过程起着重要作用。退化组与遮阴组的叶色无显著差异,但两者的PAL和CHI活性差异显著(P<0.05)且遮阴组较低,可能是因为光胁迫使植物生长受阻,而使遮阴组酶活性降低.
2.5 各指标间相关性分析
各生理指标间的相关性分析如表2.叶色参数与色素含量直接相关,L值和b值与叶绿素a、叶绿素b、类胡萝卜素含量呈显著或极显著正相关,a值与花色素苷和类黄酮含量呈极显著正相关,花色素苷和类黄酮含量与叶绿素a、叶绿素b、类胡萝卜素含量呈显著或极显著负相关.PAL和CHI活性是花色素苷合成过程中的关键酶[21-22],通过分析可得,PAL和CHI活性与L值、b值呈极显著负相关,与a值、花色素苷、类黄酮含量呈极显著正相关,可推断出,叶绿素a、叶绿素b、类胡萝卜素、花色素苷、类黄酮、PAL和CHI活性对紫叶狼尾草叶色有重要影响.
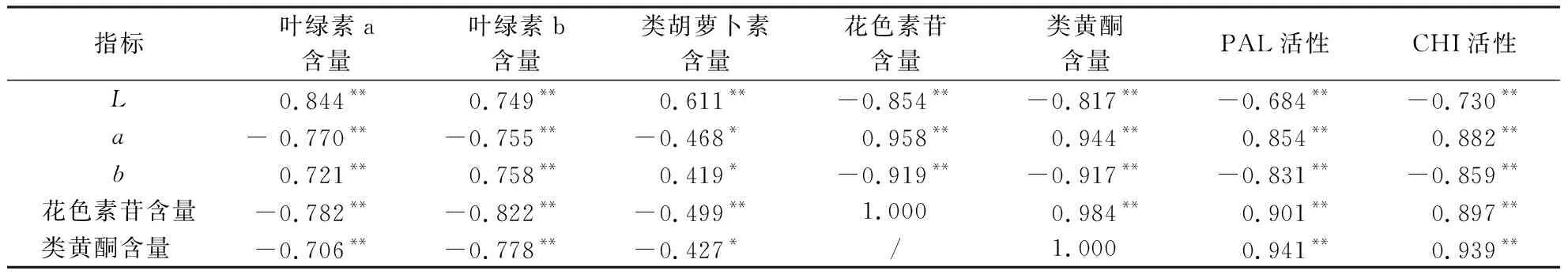
表2 3组紫叶狼尾草的叶色参数及各生理指标相关性系数1)Table 2 Leaf color parameters and physiological indexes of 3 groups of P.setaceum ‘Rubrum’
1)*和**分别表示相关系数达到0.05和0.01的显著水平.
3 结论与讨论
通过叶色参数的测定使叶色量化,可更加精确地分析叶色与各生理指标之间的关系[23].该方法用于植物叶色研究的国内文献较少,葛雨萱等[24]在研究黄栌叶色及朱景乐等[25]在研究杜仲叶色时有涉及.叶色参数计算结果表明,对照组的颜色指数远大于长期遮阴及退化的紫叶狼尾草,遮阴及退化的紫叶狼尾草叶色表现相近,均为黄绿色.
高等植物叶片中的色素主要是叶绿素a、叶绿素b、类胡萝卜素和花色素苷[26].对光合色素含量的测定表明,退化组的紫叶狼尾草叶绿素含量最高,并分析得出,可能是由于遮阴使植物受到光胁迫,叶片生长状态不佳,叶肉细胞中的叶绿体数量减少,使得遮阴组较退化组叶绿素含量低.通过相关性分析得出,光合色素与叶色参数a值呈极显著负相关(P<0.01).这与朱倩玉等[27]对4种彩叶灌木的研究结果一致.
花色素苷是植物呈现紫色的重要色素[19],类黄酮可促进花色素苷的合成.本研究结果表明,花色素苷和类黄酮含量与叶色参数a值均呈极显著正相关(P<0.01),对照组花色素苷含量最高且与其他两组差异显著(P<0.05),这与聂庆娟等[20]对红栌的研究结果一致.长期遮阴与叶色退化的花色素苷含量无显著差异,与叶色参数a值的结果相互验证.通过叶片色素含量百分比得出,对照组花色素苷占比远大于其他两组紫叶狼尾草,且这两组的色素含量百分比接近,这也是对照呈现紫色而遮阴与退化组呈现绿色的主要原因,植物体内色素含量的比例决定叶片的呈色[18].通过植物叶片各色含量素与颜色指数的分析表明,对于紫叶狼尾草叶片呈色起决定性作用的为花色素苷.
PAL活性间接决定着植物叶色的形成,它的作用是催化苯丙氨酸形成肉桂酸[28].但也有学者指出,PAL与植物花色素苷的生物合成无关,更不是限制花色素苷生成的关键酶[15]。本研究结果表明,PAL活性与叶色参数a值、花色素苷含量、类黄酮含量呈极显著正相关,与吴驭帆等对鹅耳枥[29]的研究结果一致,说明PAL是紫叶狼尾草叶色形成的关键酶。对照组的PAL活性显著高于其他两组的活性(P<0.05),虽然遮阴和退化的紫叶狼尾草花色素苷含量相近,但退化的PAL活性显著高于遮阴的活性(P<0.05).
CHI是植物花色素苷合成过程中的重要酶,在类黄酮类化合物转化过程中起着关键作用[22],也有报道指出CHI活性与花色素苷的积累在不同植物中的相关性不一致[15],该研究表明CHI活性与叶片参数a值、花色素苷含量及类黄酮含量均显著正相关,说明CHI促进了紫叶狼尾草类黄酮的转化,进而合成花色素苷,这与王庆菊等[30]对紫叶稠李叶片的研究结果类似。叶色退化的PAL和CHI活性均显著高于长期遮阴的活性,这可能是由于紫叶狼尾草受到光胁迫后,生长较弱而导致的酶活性降低。
综上可得,虽然叶色退化和长期遮阴的紫叶狼尾草在颜色上无差异,但根据两者叶绿素含量和酶活性的规律分析得出,可能由于生长环境而导致的生理指标存在差异,且光合色素、花色素苷、类黄酮和PAL和CHI活性是影响紫叶狼尾草叶色的主要生理指标。其呈色机理在分子水平上的研究有待进一步深入.
猜你喜欢
中国饲料(2022年5期)2022-04-26
广东农业科学(2018年11期)2019-01-09
现代园艺(2017年13期)2018-01-19
中国绿色画报(2017年11期)2018-01-04
河北科技师范学院学报(2017年4期)2017-03-08
广东饲料(2016年3期)2016-12-01
衡阳师范学院学报(2016年3期)2016-07-10
花卉(2016年1期)2016-02-17
作物研究(2014年6期)2014-03-01
食品工业科技(2014年6期)2014-02-27
