
c-Jun蛋白对A型流感病毒H1N1感染小鼠体内CD4+和CD8+T细胞增殖的调节作用
2019-01-25 11:34:02张守平申汶涛王丽荣张百重3
浙江大学学报(农业与生命科学版) 2018年6期
张守平,申汶涛,王丽荣,张百重3*
(1.河南科技学院动物科技学院,河南新乡453003;2.中国农业大学动物医学院,北京100192;3.河南科技学院资源与环境学院,河南新乡453003)
A型流感病毒是引发流感的重要病原,可以感染多种动物,引起发热、咳嗽、呼吸困难等呼吸系统疾病以致全身严重败血症等多种症状。经A型流感病毒感染后的复杂致病机制一直是科学家们研究的热点。杀伤性T细胞(CD8+T细胞)在病毒的清除中发挥重要作用,其作用方式为直接裂解被感染细胞或释放γ-干扰素(interferon-γ,IFN-γ)和肿瘤坏死因子-α(tumor necrosis factor,TNF-α)等促炎性细胞因子,有效控制流感病毒感染。CD8+T缺乏的小鼠感染流感病毒后呈现出更高的易感性和病毒复制量[1]。辅助性T细胞(CD4+T细胞)通过分泌IFN-γ、白介素-2(interleukin-2,IL-2)和TNF-α等来促进B细胞产生抗体和维持CD8+T细胞功能,而缺乏CD4+T细胞的小鼠呈现出流感病毒清除延迟的现象[2]。因此,CD4+T和CD8+T细胞在抗流感感染免疫中发挥着重要的作用。
c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)信号通路对CD4+和CD8+T细胞的增殖、分化等过程具有重要的调控作用[3]。c-Jun是一种原癌蛋白,也是JNK通路发挥作用的一种下游转录因子。在流感病毒感染的早期,可以检测到c-Jun活化蛋白,而其在流感病毒感染过程中是否对CD4+和CD8+T细胞发挥作用还不清楚。核酶Dz13是一种可以特异性切割并降解c-Jun蛋白的DNA分子,即一种脱氧核酶,目前已经很成熟地应用在抗肿瘤、细胞迁移等方面的研究中。有研究显示,使用Dz13阻断c-Jun蛋白可以在体外抑制单核-内皮细胞聚集,在大鼠炎症模型中阻止白细胞黏附、渗出[4]。但是关于Dz13应用于病毒感染所引起的细胞增殖或分化等的报道则很少。在本研究中,通过建立H1N1流感病毒感染模型,使用Dz13抑制c-Jun蛋白的表达,从而探讨c-Jun蛋白对病毒感染后小鼠体内CD4+和CD8+T细胞的增殖及细胞因子的调节作用。
1 材料与方法
1.1 材料
动物:6~8周龄无特定病原体(specific pathogen free,SPF)级雌性BALB/c小鼠,体质量(20±1.5)g,购自北京维通利华实验动物技术有限公司,饲养于独立通风笼盒(individually ventilated cages,IVC)系统内。
病毒:H1N1(A/PR/8/34)流感病毒毒株,由中国农业大学动物医学院国家动物寄生原虫实验室保存,于9日龄SPF级鸡胚尿囊腔传代后,测定对小鼠的半数致死量(median lethal dose,LD50),分装后于-80℃冰箱中储存,备用。
核酶Dz13和对照序列Dz13scr:核酶Dz13的序列(5′-CGGGAGGAAGGCTAGCTACAACGAGA GGCGTTG-Ti-3′)和Dz13scr的序列(5′-GCGACG TGAGGCTAGCTACAACGAGTGGAGGAG-Ti-3′)由华大基因合成。用无菌磷酸盐缓冲液(phosphate buffer solution,PBS)溶解至质量浓度为2 mg/mL,于-20℃冰箱中储存,备用。
试剂:FITC Rat Anti-mouse CD4和PE Antimouse CD8抗体购自美国eBioscience公司;红细胞裂解液购自中科迈晨科技有限公司;TRIzol为美国Invitrogen公司产品;p-c-Jun和c-Jun蛋白抗体为美国CellSignalTechnology(CST)公司产品;FuGENE®6为美国Promega公司产品。其余常规试剂为实验室内储存的药品。
1.2 方法
小鼠攻毒:将小鼠随机分为Dz13+H1N1、DZ13scr+H1N1、PBS+H1N1、PBS 4组,每组 9只。肌内注射Zoletil®(16.5 mg/kg)以麻醉小鼠,随后将3 LD50/只H1N1流感病毒溶于50 μL PBS中,以滴鼻方式给予小鼠。
小鼠给药:以感染病毒时计为0 d,在感染的0和 3 d,将 Dz13 和 Dz13scr(2.5 mg/kg)溶于 50 μL PBS(含2.5 μL FuGENE6和1 mmol/L MgCl2)中,以滴鼻方式给予小鼠。
外周血内CD4+、CD8+T细胞的检测:于感染后3和6 d时,每组小鼠各取3只,摘取眼球采血并处死。收集经抗凝剂处理的外周血200 μL,1 000 r/min离心3 min,弃上清液;加入1 mL红细胞裂解液,轻轻吹打均匀,室温静置10 min,1 000 r/min离心5 min,弃掉上层红色液体;收集沉淀部分,加入1 mL PBS洗涤,1 000 r/min离心5 min,弃上清液;加入0.5 mL PBS重悬,加入FITCAnti-mouse CD4抗体和PE Anti-mouse CD8抗体,混匀,避光静置约20 min;用BDAccuri®C6 Plus流式细胞仪检测。
用蛋白质印迹法(Western-blotting,WB)检测肺内c-Jun蛋白的表达:在取下来的肺组织(大约全肺的1/3)中加入1 mL RIPA蛋白裂解液[含10 mmol/L苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)],在低温条件下用玻璃研磨器将组织匀浆。在冰上作用30 min后,以1.2×104r/min离心10 min,取上清液。按比例加入上样缓冲液,煮沸10 min。用二喹啉甲酸(BCA)法测定蛋白质量浓度后,将各组蛋白样品质量浓度调整为2 μg/μL,取18 μL样品进行聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,PAGE),用湿转法将蛋白胶样品于60 V电压下2 h内转膜至聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)上。用5%脱脂奶粉室温封闭l h。一抗用p-c-Jun(1∶1 000)抗体、c-Jun(1∶1 000)抗体和β-tubulin(1∶1 000)抗体孵育过夜(4℃),用0.5%PBST溶液洗膜后,用HRP标记的羊抗兔二抗(1∶2 000)室温孵育1 h。经0.5%PBST溶液洗膜后,用ECL化学发光试剂盒显色并进行结果分析。
用实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)检测肺内细胞因子的表达:取100 μL外周血,用红细胞裂解液裂解后,1 000 r/min离心10 min,收集沉淀细胞。按照TRIzol说明书提取各组细胞总RNA。取2 μg总RNA,按照全式金反转录试剂盒操作说明将RNA反转录为cDNA,以20 μL反应体系进行PCR扩增。每个样本设3个重复。反应程序:95℃预变性90 s;95 ℃退火5 s,58 ℃延伸30 s,共30个循环。获得各样本待测基因的扩增循环阈值(CT)值,采用 2-△△CT
法进行数据处理。引物序列见表1。
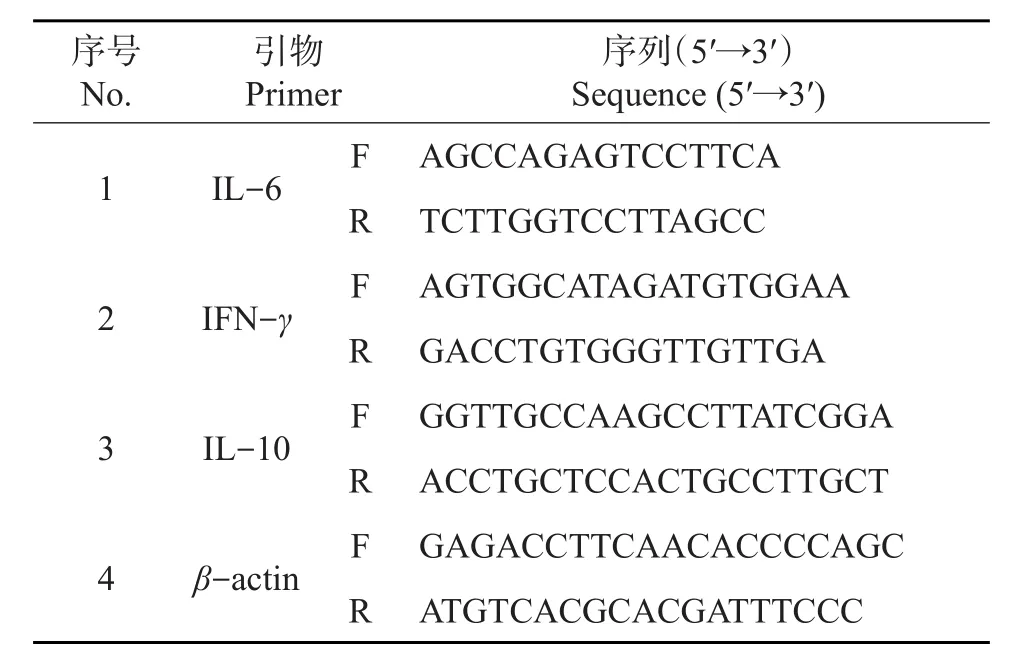
表1 细胞因子的引物序列Table1 Primer sequences of the target cytokines
1.3 数据分析
采用Graphpad Prism 6.0软件作图。采用SPSS 17.0软件中的单因素方差分析及Tukey事后检验对试验所得数据进行统计学分析。
2 结果与分析
2.1 Dz13对c-Jun蛋白表达的抑制作用
用蛋白质印迹法检测肺组织内c-Jun蛋白的表达量,结果(图1)表明:小鼠攻毒后,与未感染组(PBS)相比,肺内c-Jun(p-c-Jun)蛋白的活化水平明显上调,随着感染后时间的递增,在6 d时肺内c-Jun蛋白的活化程度明显低于3 d时的水平。Dz13用药组(Dz13+H1N1)c-Jun蛋白(t-c-Jun)的表达受到抑制,在3 d时的抑制水平高于6 d,随着t-c-Jun蛋白表达受到抑制,活化的c-Jun蛋白表达量也随之降低。说明病毒感染后可以引起c-Jun蛋白的活化,核酶Dz13可以特异性地抑制c-Jun蛋白表达及其活化。
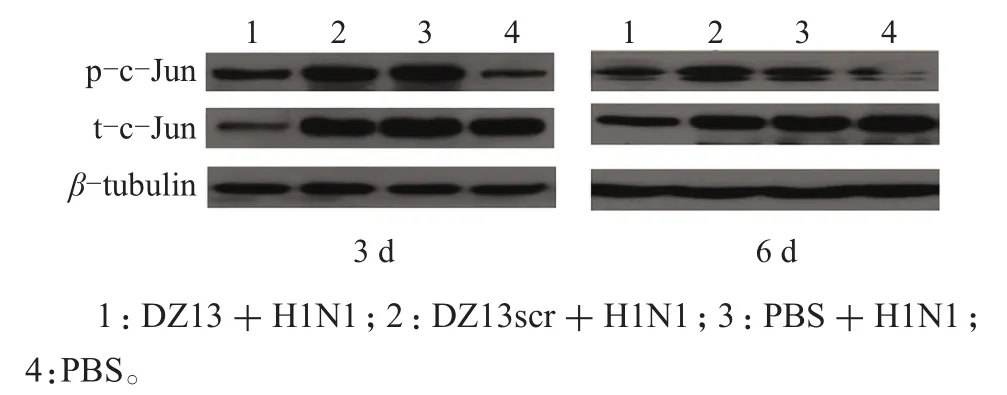
图1 Dz13对病毒感染小鼠肺内c-Jun蛋白表达的影响Fig.1 Effect of Dz13 on c-Jun protein expression in lungs of infected mice
2.2 c-Jun蛋白对病毒感染引起的CD4+和CD8+T细胞增殖的影响
病毒感染机体后,可以迅速激起机体的免疫反应。使用流式细胞术检测感染后3和6 d小鼠外周血内CD4+和CD8+T细胞的增殖,结果(图2)表明:小鼠感染病毒后,外周血内CD4+和CD8+T细胞的数量与对照组相比明显增多;而核酶Dz13用药组与Dz13scr相比,CD4+和CD8+T细胞的增殖受到抑制(P<0.05)。说明c-Jun蛋白参与调节流感病毒感染后小鼠体内CD4+和CD8+T细胞的增殖。
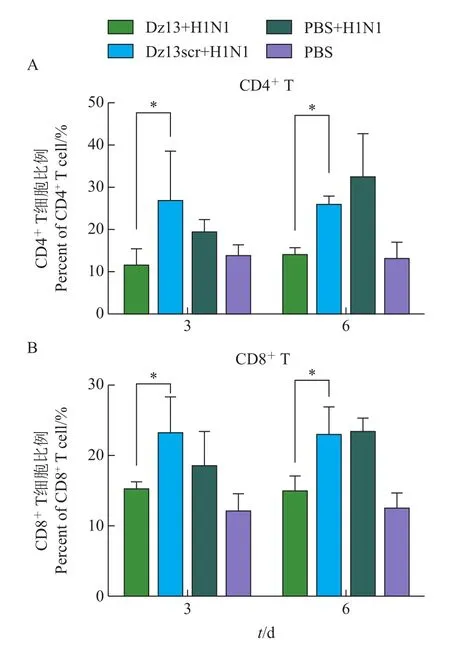
图2 Dz13对病毒感染小鼠外周血内CD4+(A)和CD8+(B)T细胞增殖的影响Fig.2 Effect of Dz13 on proliferation of CD4+(A)and CD8+(B)T cells in peripheral blood of infected mice
2.3 c-Jun蛋白对病毒感染后引起的细胞因子表达的调节作用
流感病毒感染后一个特征性变化就是细胞因子表达量的上调。在本研究中,为测定c-Jun蛋白对病毒感染后细胞因子表达量的调节作用,我们使用qRT-PCR测定感染后3和6 d小鼠外周血内细胞因子的表达情况。从图3中可见:病毒感染后,细胞因子IFN-γ、IL-6和IL-10明显地上调表达;Dz13用药后,Dz13+H1N1组细胞因子IFN-γ、IL-6和IL-10表达量明显低于Dz13scr+H1N1组和PBS+H1N1组(P<0.05)。说明c-Jun蛋白参与调节流感病毒感染后细胞因子的表达。
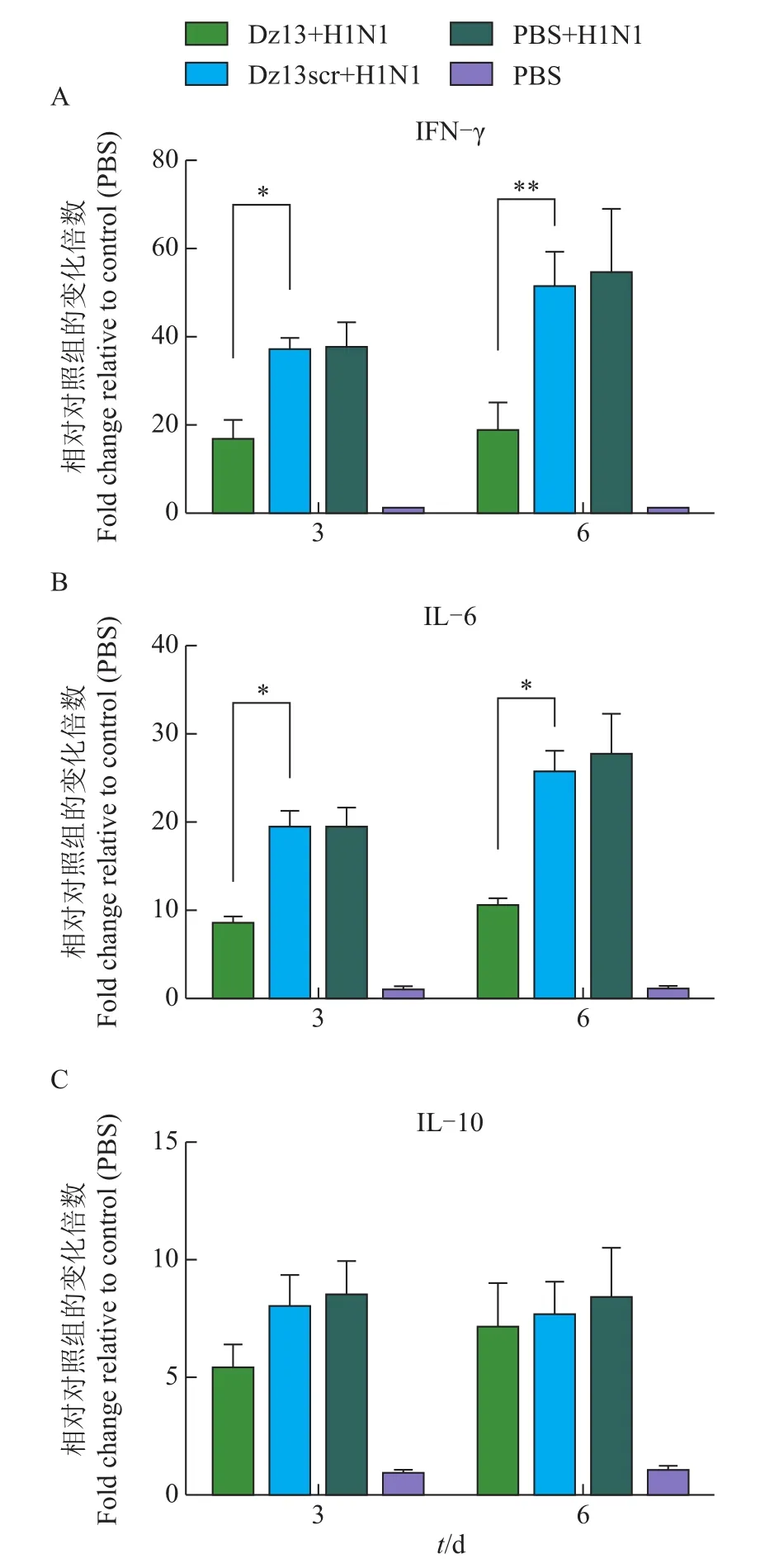
图3 Dz13对病毒感染小鼠外周血内细胞因子表达量的影响Fig.3 Effect of Dz13 on cytokine expression in peripheral blood of infected mice
3 讨论
原癌蛋白c-Jun和c-fos以异二聚体的形式组成转录因子AP-1(activator protein-1),AP-1可以调控多种基因的转录,是JNK通路中发挥生物学效应的主要分子,参与细胞增殖、分化、凋亡等的调节过程。大量研究证实:在流感病毒感染的早期可以检测到c-Jun活化蛋白(p-c-Jun),并启动下游IFN-β等一些抗病毒细胞因子的转录;抑制c-Jun蛋白表达后能够导致IFN-β转录水平降低,病毒繁殖率升高[5-6]。同时,有研究发现,JNK-/-小鼠可抑制感染后引起的CD8+T细胞的增殖[7]。而c-Jun蛋白作为JNK通路的效应因子是否参与流感病毒感染后CD4+和CD8+T细胞的增殖还不清楚。在本研究中,阻断小鼠体内c-Jun蛋白的表达后,流感病毒感染后c-Jun蛋白的活化明显降低,外周血内CD4+和CD8+T细胞的增殖均受到抑制。进一步证实了c-Jun蛋白作为JNK通路的效应分子,在流感病毒感染后所引起c-Jun蛋白的活化,参与了T细胞增殖和炎性反应等的调节。
脱氧核酶DRz是人工合成的具有酶活性的DNA分子,能在一定条件下识别并结合靶底物mRNA于配体结合部位形成螺旋,催化切割后与之分离,再结合其他底物,从而持续地抑制底物基因的表达。因其廉价、低毒、高稳定性、高特异性等优点,目前已成为分子生物医药等多个领域的研究热点。c-Jun蛋白在肺癌、肝癌等多种肿瘤中异常表达,在肿瘤的发生发展中发挥着重要作用,因此,c-Jun蛋白在肿瘤及抗感染治疗中可以作为一个有效靶点而存在。Dz13作为脱氧核酶的一种,通过与c-Jun核酸序列的特异识别,发挥基因表达抑制效应和高效的剪切活性。目前已经广泛应用在抗皮肤癌、血管形成、细胞增殖迁移等方面的研究中[8-9]。将Dzl3体外转染至人类微血管内皮细胞中,发现c-Jun蛋白表达减少,裸鼠皮肤鳞状细胞癌、黑色素瘤等皮肤癌细胞的迁移、侵袭受到抑制[10]。在本研究中,Dz13作用于流感病毒感染小鼠后,可以明显抑制流感病毒感染引起的c-Jun蛋白的活化和表达及CD4+和CD8+T细胞的增殖和炎性因子的上调表达。此研究结果提示我们,在流感病毒感染后可以通过使用特异性Dz13阻断c-Jun蛋白表达来抑制感染后的过度炎性反应。结合Dz13的诸多优势,其在抗病毒感染应用中将具有重要的潜能,可以为抗病毒药物的筛选提供参考。
猜你喜欢
中老年保健(2022年1期)2022-08-17 06:14:30
材料与冶金学报(2022年2期)2022-08-10 09:15:38
作文成功之路·小学版(2020年6期)2020-07-27 01:48:28
广东医科大学学报(2020年6期)2020-02-06 06:00:40
兽医导刊(2019年1期)2019-02-21 01:14:20
猪业科学(2018年8期)2018-09-28 01:27:52
现代检验医学杂志(2016年3期)2016-11-15 01:59:50
现代检验医学杂志(2016年1期)2016-11-12 13:19:42
分析测试学报(2015年6期)2016-01-13 06:19:03
无机化学学报(2014年10期)2014-02-28 17:33:13
