
杭州市树附生苔藓植物分布特征及其与环境的关系
2019-01-25 11:34莫亚鹰胡忠健茹雅璐吴玉环
浙江大学学报(农业与生命科学版) 2018年6期
盛 威,莫亚鹰,胡忠健,茹雅璐,吴玉环*
(1.杭州师范大学生命与环境科学学院,生态系统保护与恢复杭州市重点实验室,杭州310036;2.中国科学院沈阳应用生态研究所,森林生态与管理重点实验室,沈阳110016;3.杭州植物园,杭州310007)
树附生苔藓植物是指生长在活的乔灌木树皮上的苔藓植物种类[1],是许多生态系统的重要组成成分,在生态系统功能维持中具有重要作用[2-4]。由于树附生苔藓植物的营养物质主要来自雨水、露水及大气尘埃的沉积物[5],致使其对环境因子的敏感度可达到种子植物的10倍[6],是一类被世界各国广泛应用于监测环境质量的生物指示物[7-10],也是森林完整性的指示物[11-13]。基于它们的附生特性,树附生苔藓植物与树种、树龄、树体大小、树皮理化性质及它们在树干附生高度、位置等各种与附主相关的要素之间关系复杂[14-17],其种类、群落类型和分布格局呈现多样化,受到苔藓植物学家的普遍关注[18-19]。SLACK[20]研究发现树附生苔藓植物与附生位置高度密切相关,而TRYNOSKI等[21]认为多数树附生苔藓植物偏向于树干的某一高度,不同朝向也会对树附生苔藓植物的分布产生影响。也有不少研究者[22-27]认为树附生苔藓植物的分布受附生树种类的影响最为显著。
国内对城市生态系统中树附生苔藓植物群落的调查主要集中在长江三角洲地区,以上海市[28-29]、杭州市[30-31]为主。刘艳[32]曾对杭州市的树附生苔藓植物进行过标本采集和简单普查,但未进行样点的样方调查和数量生态学研究。官飞荣等[30-31]在进行较大范围的生态调查后,对杭州市树附生苔藓植物的多样性和附生专一性进行了相关报道。本研究进一步通过对杭州市树附生苔藓植物多样性与分布规律进行分析,填补了杭州市树附生苔藓植物分布特征研究的空白,从而为合理保护和开发杭州市苔藓植物资源及其生境提供基础数据和参考策略。
1 研究方法
1.1 调查区域
于2011年7月、9月及2012年7月,以杭州西湖为中心,选取杭州市区的公园、景点、大学校园及湿地共21个样点[30-31](图1)(附表1,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2017.07.241)进行调查。
样点分别为杭州植物园(1)、西湖一公园(14)、杭州市少年宫(15)、西湖六公园(16),共4个公园;浙江大学玉泉校区(2)、浙江大学之江校区(4)、杭州师范大学玉皇山校区(11)、浙江大学华家池校区(17)、杭州师范大学文一路校区(18)、浙江大学紫金港校区(19),共6个大学校园;五云山(3)、栖霞岭(5)、曲院风荷(6)、九溪(7)、龙井(8)、虎跑(9)、玉皇山(10)、柳浪闻莺(12)、吴山天风(13),共9个旅游景点;和睦湿地(20)和西溪湿地(21),共2个湿地。
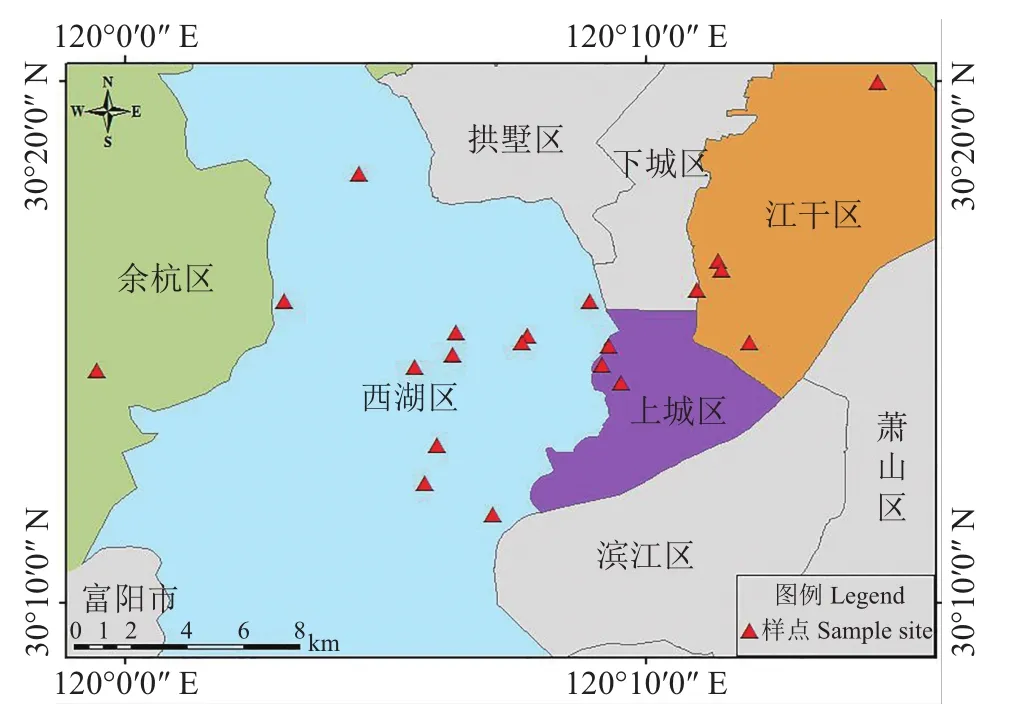
图1 杭州市树附生苔藓植物样点分布Fig.1 Sampling sites of epiphytic bryophytes in Hangzhou City
1.2 野外调查
野外样方调查包括对树附生苔藓植物的种类调查、盖度调查、标本采集等。本次调查的研究对象是21个样点内胸径大于15 cm的树木,共178棵,在距离地面30、110、150、180 cm的树干高度处,以及树干的东、南、西、北4个面分别设立10 cm×10 cm的样方,用网格法测定每个样方内每种树附生苔藓植物的盖度,得到所有调查树木在16个样方上的总盖度,作为分析所需的基础数据[29-31]。采集样方内苔藓植物标本带回实验室,先用解剖镜和显微镜观察,再参考《中国苔藓志》等文献资料及馆藏标本对其逐一鉴定到种,凭证标本存放于杭州师范大学植物标本馆(HTC)里。
1.3 数据处理与分析
在野外调查获取原始数据的基础上,用Microsoft Excel 2003和SPSS 19.0对2 848个树附生苔藓植物样方进行统计。对21个样点的不同高度位置和朝向的数据按最大值法进行标准化处理,最终得到1个21×48的样方-物种矩阵,2个21×4的样方-环境因子矩阵。然后采用PCORD 5.0的双向指示种分析法(two-way indicator species analysis,TWINSPAN)[33]完成群落的分类,采用CANOCO 4.5的除趋势对应分析法(detrended correspondence analysis,DCA)[34-35]完成群落排序,并通过CanoDraw 4.0完成相应排序图。根据数据矩阵,采用CANOCO 4.5的典型相关分析(canonical correlation analysis,CCA)完成盖度数据计算,并由CanoDraw 4.0生成物种-环境因子和样方-环境因子的排序图。在运行CANOCO 4.5的DCA和CCA程序时,对种类盖度数据进行开平方处理。
2 结果与分析
2.1 杭州市树附生苔藓植物种类调查
对杭州市21个样点内的178棵树上的2 848个样方的树附生苔藓植物进行物种鉴定,共发现树附生苔藓植物48种,隶属于22科37属,其中藓类植物18科32属42种,苔类植物4科5属6种(表1)。
2.2 杭州市各样点树附生苔藓植物种类分布
树附生苔藓植物种类最多的样点为西湖一公园(14),共24种;苔藓植物种类最少的样点为浙江大学紫金港校区(19),只有1种,为中华细枝藓(Lindbergia sinensis)(表2)。其中,所含苔藓植物种类≥20的有8个样点,分别是杭州植物园(1)、浙江大学玉泉校区(2)、栖霞岭(5)、虎跑(9)、西湖一公园(14)、杭州市少年宫(15)、浙江大学华家池校区(17)和杭州师范大学文一路校区(18)。
2.3 杭州市各样点的分类结果与分析
将杭州市21个样点用TWINSPAN法进行等级分类(图2),其聚类分组结果与DCA的排序结果(图3)基本一致。DCA排序图以二维图的形式直观反映了各样点间树附生苔藓植物在种类组成上的关系及分布格局。沿着第1轴从左到右,适宜树附生苔藓植物分布的生境情况逐渐恶化,样点组Ⅳ的生境最适宜树附生苔藓植物生长,样点组Ⅰ和Ⅱ的生境较适宜,样点组Ⅲ的生境较不适宜,而样点组Ⅴ的生境最不适宜树附生苔藓植物的生存。DCA排序图的第2轴主要反映了各样点组在海拔高度上的变化,即沿着第2轴从下到上,样点的海拔逐渐降低,样点组Ⅱ的2个样点均为海拔超过50 m的山林,明显不同于其他4个样点组,因而较为独立地分布在第2轴的底线位置。说明DCA排序轴能较好地反映杭州市树附生苔藓植物群落的分布格局及生态关系[36]。
样点组Ⅰ:包括浙江大学之江校区(4)、曲院风荷(6)、龙井(8)、杭州师范大学玉皇山校区(11)、九溪(7)、玉皇山(10)、和睦湿地(20)、西溪湿地(21),共8个样点。除和睦湿地和西溪湿地外,它们均位于西湖风景名胜区内,毗邻杭州西湖,远离市区交通主干道,历史悠久,生境普遍良好,植被多样性高且有多株树龄超百年的古树,为树附生苔藓植物的生长提供了适宜的生境。苔藓植物多为常见的城市树附生种类,共25种,总盖度为11.496 1%,在5个样点组的总盖度中排名第2,其中柱蒴绢藓(Entodon challengeri)的平均盖度最大,为2.775 4%(附表2,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2017.07.241)。
样点组Ⅱ:包括五云山(3)、吴山天风(13),共2个样点。两者均属丘陵地区,植被资源丰富且保护良好,林冠郁闭度较高,其中五云山海拔超过300 m,在西湖群山中排列第3;吴山天风景区位于西湖东南面,海拔为94 m。本次调查在两山所取的样点海拔均超过50 m,其他19个样点平均海拔为23 m,有明显的区别,可能是海拔的梯度变化形成了独特的立体小气候,为树附生苔藓植物提供了多样化的小生境,引发了其生态分布的分化[37]。该样点组以灰藓科、锦藓科等生态幅宽广的种类为主,共14种。树附生苔藓植物的总盖度为10.232 4%,在5个样点组的总盖度中排名第3,东亚金灰藓(Pylaisia brotheri)的平均盖度最大,为4.478 3%(附表2)。
样点组Ⅲ:包括浙江大学玉泉校区(2)、柳浪闻莺(12)、西湖一公园(14)、杭州市少年宫(15)、西湖六公园(16)、浙江大学华家池校区(17)、杭州师范大学文一路校区(18),共7个样点。各样点均毗邻市中心的交通主干道,人类活动频繁,人为干扰强烈,大气污染较重,空气较干燥,综合环境条件较差。该样点组树附生苔藓植物以碎米藓科、羽藓科、牛舌藓科、锦藓科为主,共计31种,总盖度为8.429 6%,在5个样点组的总盖度中排名第4,其中八齿碎米藓(Fabronia ciliaris)的平均盖度最大,为1.428 6%(附表2)。狭叶小羽藓(Haplocladium angustifolium)和细叶小羽藓(H.microphyllum)是典型的抗旱、抗污染、抗干扰能力突出的藓类,对大气和土壤污染物的富集能力明显,是一种能有效监测环境污染的生物指示剂[39-41]。

表1 杭州市树附生苔藓植物种类Table1 Species of epiphytic bryophytes in Hangzhou City
样点组Ⅳ:包括杭州植物园(1)、栖霞岭(5)和虎跑(9),共3个样点。各样点均位于西湖西南方的丘陵地区,属于在原生林基础上人工改造的公园和景点,落成时间久远,植被多样性较高且树龄较长,林冠郁闭度和空气湿度较大。该样点组树附生苔藓植物共21种,其中苔类植物5种。树附生苔藓植物的总盖度为21.985 2%,其中苔类的总盖度为12.426 0%,芽胞裂萼苔(Chiloscyphus minor)的平均盖度最大,为11.716 6%(附表2)。该样点组的树附生苔藓植物总盖度、苔类种类数和苔类总盖度在5个样点组中均属最高,对生境要求的苛刻程度高于藓类的苔类植物较多地分布在该样点组的树木上,从很大程度上说明样点组Ⅳ的综合环境条件最适宜树附生苔藓植物分布。
样点组Ⅴ:仅有浙江大学紫金港校区(19)1个样点。该样点始建于2001年,落成时间在各样点中最短,人为活动干扰严重,森林稀疏,空气湿度较低,树木虽然胸径较大但均为近期移栽,难以有苔藓植物附生。该样点组仅有中华细枝藓,总盖度也仅为1.368 8%(附表2),物种数和总盖度在5个样点组中均最少,仅分布于紫金港校区建设过程中原生残留下来的树木上。相对而言,样点组Ⅴ的综合环境条件最不适宜树附生苔藓植物的生长。
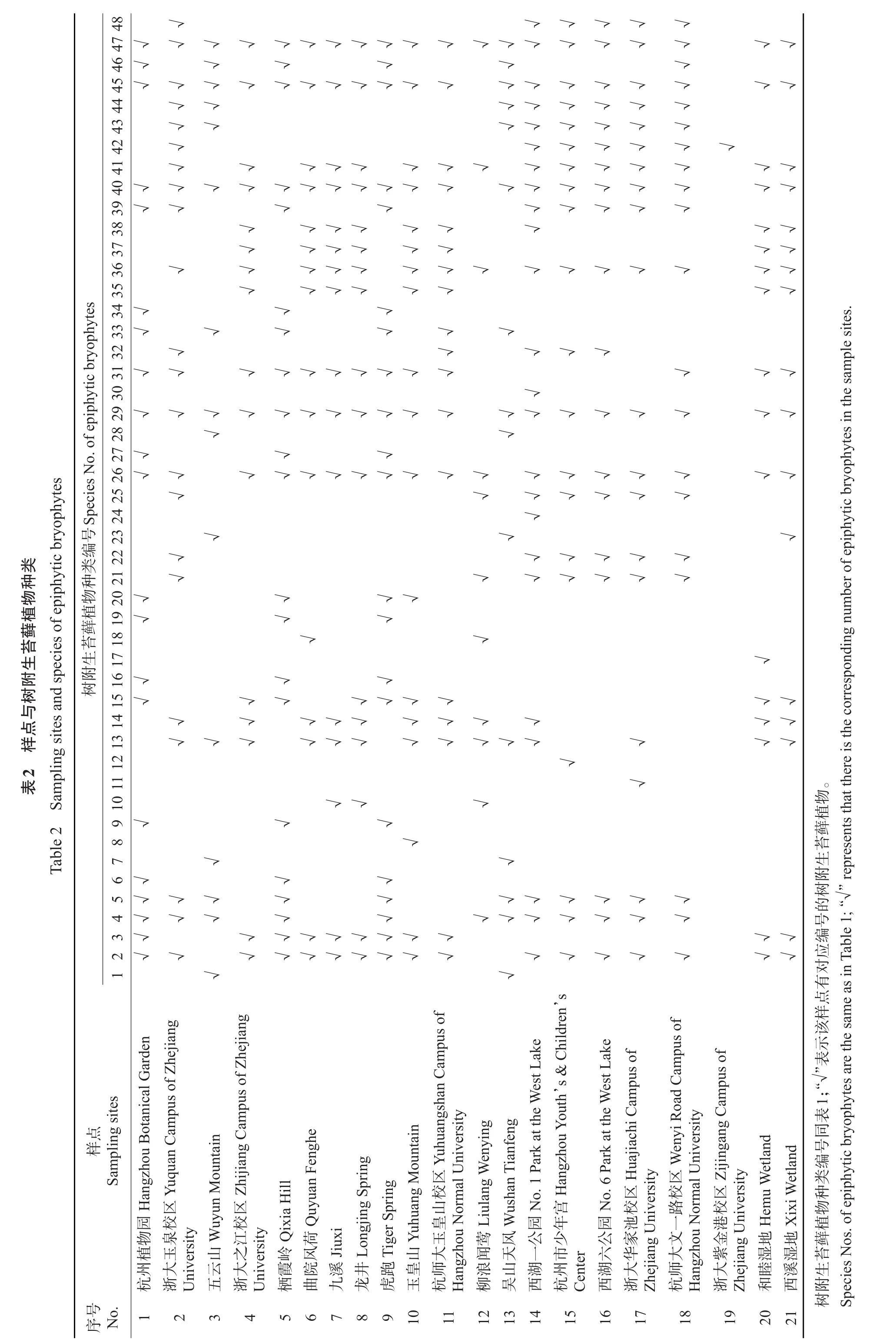

图2 杭州市树附生苔藓植物分布格局的TWINSPAN图Fig.2 TWINSPAN diagram of distribution patterns of the epiphytic bryophytes at 21 sampling sites in Hangzhou City
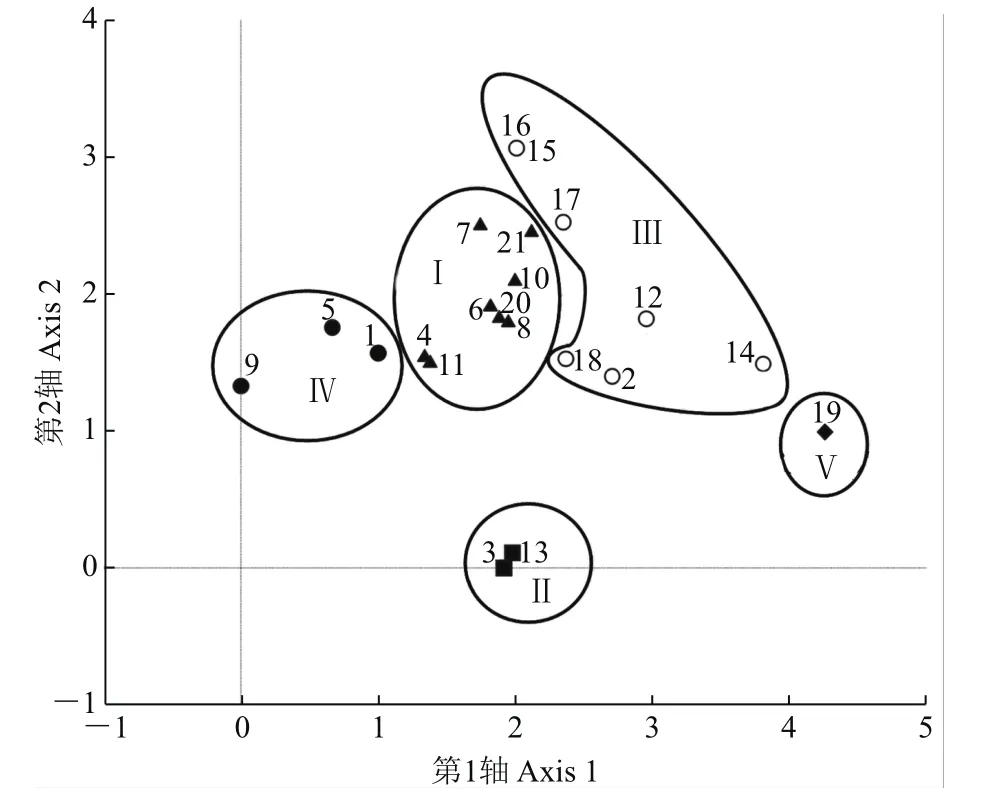
图3 杭州市21个样点树附生苔藓植物分布格局的DCA排序图Fig.3 DCAordination diagram of distribution patterns of epiphytic bryophytes at 21 sampling sites in Hangzhou City
2.4 树附生苔藓植物分布与环境的关系
相比于地面生苔藓植物,树附生苔藓植物对生境的依赖性更高,对环境因子变化的敏感度也相应增高。通过分析不同树干朝向和高度因子对树附生苔藓植物的影响可用来说明树附生苔藓植物分布特征与环境之间的关系。
2.4.1 树附生苔藓植物分布与树干朝向之间的关系
48种树附生苔藓植物在4个树干朝向上的盖度(附表3,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2017.07.241),清楚地反映出南面朝向树干上的树附生苔藓植物的盖度和种类数均最小,而北面朝向的树干上的树附生苔藓植物的盖度、种类数和特有种数均最多,另外,芽胞异萼苔、柱蒴绢藓、东亚金灰藓、华东附干藓(Schwetschkea courtoisii)、橙色锦藓(Sematophyllum phoeniceum)和东亚碎米藓(Fabronia matsumurae)在树干4个朝向分布上均占优势。
对21个样点的树干朝向因子及48种树附生苔藓植物盖度数据进行典型相关分析(CCA),可以看到:第1排序轴向左主要反映了树附生苔藓植物倾向于分布在树干西面和南面的变化趋势;第2排序轴向上主要反映了树附生苔藓植物倾向于分布在树干南面而远离树干西面的变化趋势(表3)。同时表明,4种树干朝向因子间均呈较高的正相关关系,其中东面因子与北面因子间的相关系数最大(0.888 9)。4种树干朝向因子间的相似性较高,说明树附生苔藓植物在树干朝向上的分布没有明显差异,苔藓植物在南面树干和北面树干的相似性最弱,这与前文4个朝向树干上的树附生苔藓植物的盖度和多样性情况相吻合,由树干朝向引发的光照和水分条件的变化可能是影响苔藓植物分布的因素之一,且该因素的作用比较有限。
应用CCA分别对树附生苔藓植物种类与朝向因子、样点与朝向因子进行排序(图4、图5)。4个朝向因子均指向CCA排序图第1轴正方向,4个朝向因子间的夹角均为锐角,说明它们之间有较强的正相关关系,其中东面因子与西面因子的相似性最强。结合苔藓植物种类在排序图上的分布情况,大致可以将48个苔藓植物种类分为3组。反映出树附生苔藓植物的种类分布在4个树干朝向上有一定的差异,南面因子较独立于其他3个朝向因子,但超过半数的树附生苔藓植物种类远离该4个朝向因子,大多数苔藓植物种类的排布比较分散且对朝向因子没有严格要求。说明绝大多数树附生苔藓植物种类对附生树干的朝向没有明显的专一性,仅是少数种类存在对树干朝向的一定的倾向性。
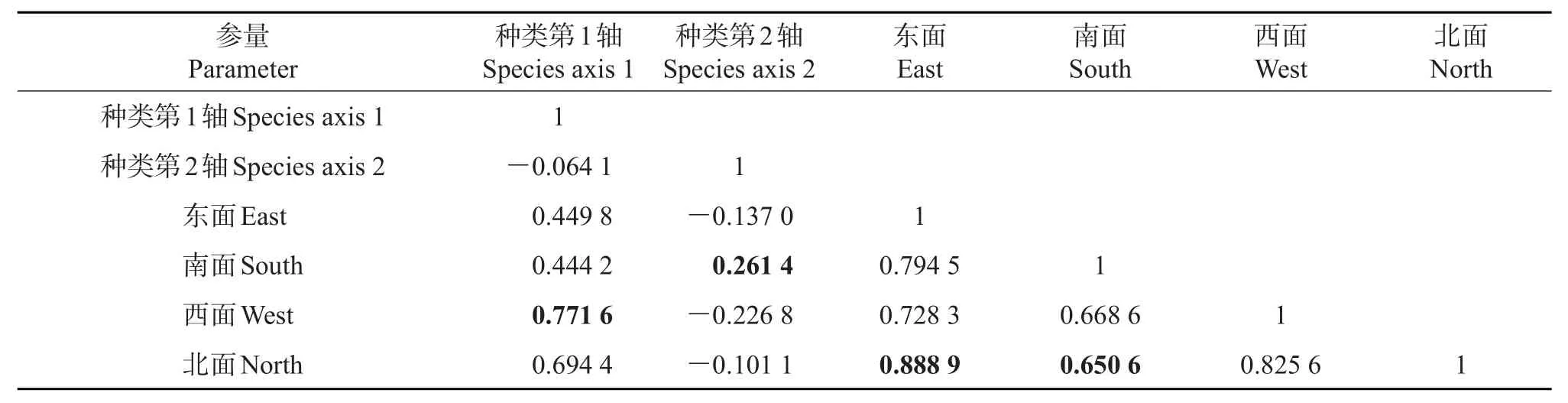
表3 树附生苔藓植物种类排序轴间、朝向因子间、朝向因子与排序轴间的相关系数Table3 Correlation coefficients of epiphytic bryophyte species axis 1 and axis 2,four orientation factors,orientation factors and two axes
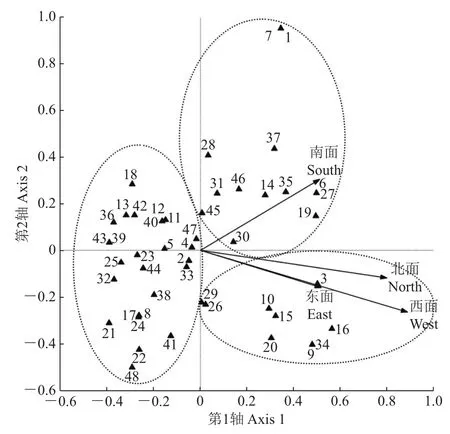
图4 树附生苔藓植物种类-朝向因子的CCA排序图Fig.4 CCA ordination diagram of epiphytic bryophyte species and four orientation factors
结合样点在排序图上的分布情况,大致可以将21个样点分为3组(图5)。样点与朝向因子的CCA排序图表明,21个样点的树附生苔藓植物的分布在树干朝向上有较弱的分化差异,仅有3个样点的苔藓植物倾向于分布在树干南面,另3个样点倾向于分布在树干北面、东面和西面,绝大多数的样点远离4个朝向因子而呈散点分布。说明树附生苔藓植物的分布与树干朝向没有明显的相关性。也说明在生境错综复杂的城市生态系统里,树干朝向对树附生苔藓植物分布的影响比较微弱。
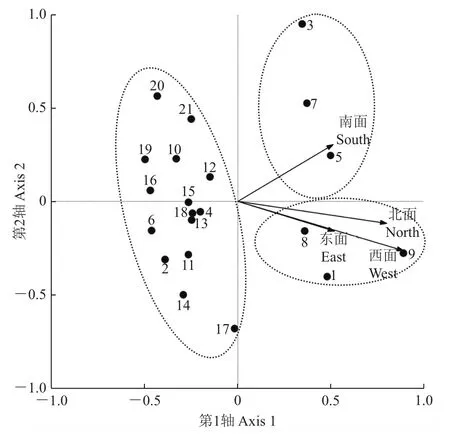
图5 样点-朝向因子的CCA排序图Fig.5 CCA ordination diagram of sampling sites and four orientation factors
2.4.2 树附生苔藓植物分布与树干高度之间的关系
树附生苔藓植物在树干顶部的盖度最大,而在树干中部的盖度最小;在树干上部的种类数最大,而在树干基部的种类数最小(附表4,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2017.07.241)。对21个样点的高度-朝向因子及48种树附生苔藓植物盖度数据进行典型相关分析(CCA)。结果(表4)表明:第1排序轴向左主要反映了树附生苔藓植物倾向于分布在树干基部的变化趋势;第2排序轴向上主要反映了树附生苔藓植物倾向于分布在树干基部而远离树干顶部的变化趋势。4种树干高度因子间均呈较高的正相关关系,上部因子与顶部因子间的相关系数最大(0.955 4);基部因子与顶部因子间的相关系数最小(0.576 7)。4种树干高度因子间相似性较高,表明树附生苔藓植物在4个树干高度上的分布差别不大,其中树干上部和树干顶部的相似性最强,而树干基部和树干顶部的相似性最弱,说明树附生苔藓植物在4个树干高度上的分布情况有一定分化,但没有显著性差异。
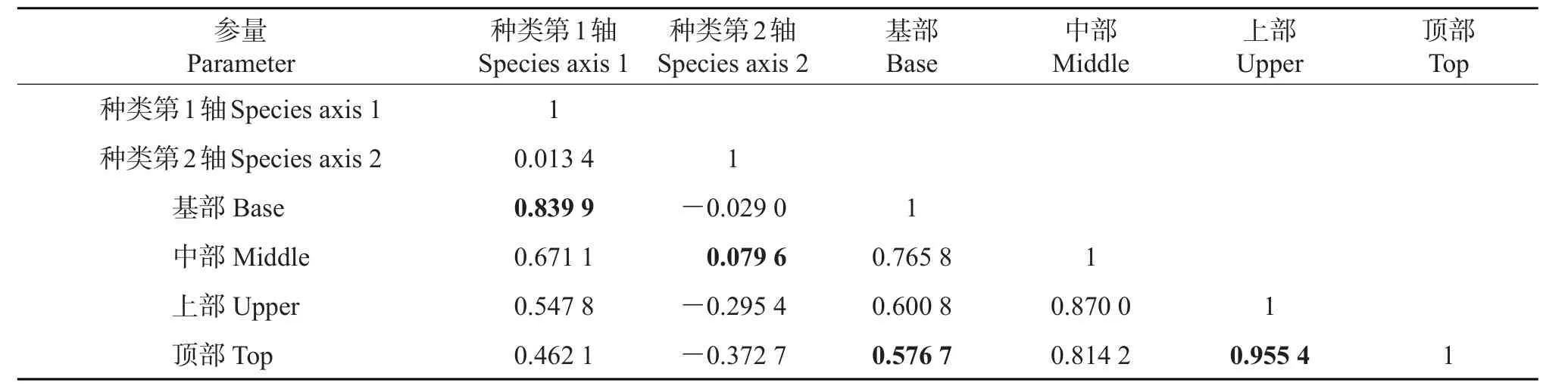
表4 树附生苔藓植物种类排序轴间、高度因子间、高度因子与排序轴间的相关系数Table4 Correlation coefficients of epiphytic bryophyte species axis 1 and axis 2,four height factors,height factors and two axes
应用CCA分别对树附生苔藓植物种类与高度因子、样点与高度因子进行排序(图6、图7)。4个高度因子彼此相关性较高,绝大多数(共计33种,占总种数的68.75%)树附生苔藓植物种类远离4个高度因子而呈散点分布,表明受调查的树附生苔藓植物种类对附生树干的高度位置没有明显的专一性。样点与高度因子的CCA排序图显示,4个高度因子间有较强相关性,绝大多数(15个样点,占总样点数的71.43%)样点远离4个高度因子而呈散点分布。只有西溪湿地(21)的树附生苔藓植物倾向于分布在树干基部。该样点作为水网密布、规划良好的城市边缘湿地公园,与其他样点的生境有明显的不同,附生树干基部的空气湿度较大、空气质量较好,适合树附生苔藓植物的生长。
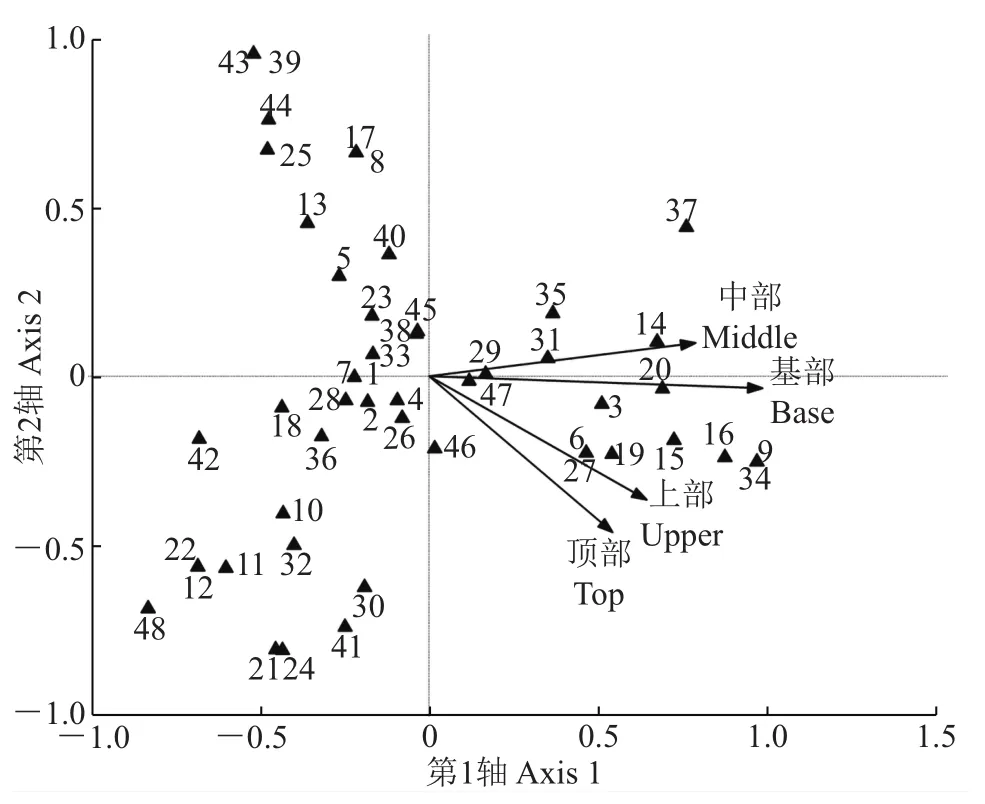
图6 树附生苔藓植物种类-高度因子的CCA排序图Fig.6 CCA ordination diagram of epiphytic bryophyte species and four height factors
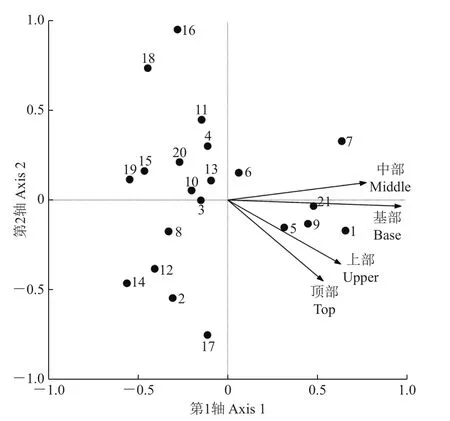
图7 样点-高度因子的CCA排序图Fig.7 CCAordination diagram of sampling sites and four height factors
3 讨论
本文通过TWINSPAN数量分类法和DCA排序法,对21个样点进行定量分析并形成聚类图和排序图,发现杭州市树附生苔藓植物可以划分为5个样点组。通过比较各样点组的物种组成和盖度情况,再结合各样点的实际生境状况,可以有效地反映树附生苔藓植物在杭州市的分布格局及与环境的关系。
就苔藓植物对各样点组生境的适宜度而言,样点组Ⅳ相对最适宜,样点组Ⅰ次之,样点组Ⅱ再次,样点组Ⅲ不适宜,样点组Ⅴ最不适宜。就各样点组的树附生苔藓植物的多样性情况而言,排列顺序同生境的适宜度相同。说明通过分析树附生苔藓植物的种类、盖度等多样性指标及分布情况,可以大致推断各样点乃至整个城市适宜苔藓植物生存的环境状况,尤其是反映环境污染状况、空气质量和人为干扰程度。徐晟翀等[28]、安丽等[39]、曹同等[42]的研究发现,上海地区苔藓植物的多样性水平和分布格局与环境状况有密切的关系,从环境污染和人为干扰严重的市中心到相对清洁的郊区,苔藓植物尤其是树附生苔藓植物的多样性水平呈明显递增的趋势,其种类组成和生长状况也有明显的差异。通过分析杭州市树附生苔藓植物分布格局与样点生境的关系,发现树附生苔藓植物倾向于生长在建成时间久远、附生树木树龄较长、人为干扰程度较小、空气湿度较高、环境污染较少、林冠郁闭度较大的样点中。树附生苔藓植物对具体生境的依赖性比较高,生境异质性的作用十分显著,多种多样的环境因子在相互影响、相互制约中对树附生苔藓植物产生复杂的综合作用。
本研究发现样地历史和树龄较长的树木适宜于树附生苔藓植物的分布。QUARTERMAN[43]和SONG等[44]对森林生态系统的研究也认为,林龄、树龄对树附生苔藓植物的相对优势度影响明显,苔藓植物定居树干的机会随着树龄的增加而增加。STUDLAR[45]也认为,树附生苔藓植物的群落组成(物种丰富度和生活型等)会随着树龄的增加而发生变化,树木生长会改变树皮的性质和树干的小生境,继而引起苔藓植物群落的演替。郭水良等[46]发现,森林郁闭度不同引起的空气湿度差异是造成不同森林类型树附生苔藓植物组成和盖度差异的一个重要原因。本研究也表明,城市树附生苔藓植物一般倾向于生长在林冠郁闭度和空气湿度较大的生境中,但如羽藓科小羽藓属等部分抗旱、抗干扰能力强的苔藓植物种类,依然能分布在空气湿度较低、人为干扰较严重的市中心附近,这些生态幅较宽、对环境条件要求较低的树附生苔藓植物便更能发挥监测城市环境质量和变化的作用。
此外,本文利用定量方法分析了城市树附生苔藓植物分布与树干空间位置(朝向和高度)的关系。结果显示,树附生苔藓植物的种类组成、盖度等多样性水平在不同朝向和不同高度上都有一定的分化差异,但不存在占绝对优势的树干朝向或树干高度。CCA排序结果也表明,绝大多数的树附生苔藓植物种类和样点与朝向因子、高度因子之间没有紧密联系,城市树附生苔藓植物在附生树干的定居对树干空间位置的要求并不严格。总之,树干朝向和高度对杭州市树附生苔藓植物的分布没有显著影响,这与有关学者对森林生态系统中的树附生苔藓植物的研究结果不同。刘蔚秋等[17]、PATRICIA等[19]、TRYNOSKI等[21]、曹同等[47]、刘冰等[48]的野外调查显示,树附生苔藓植物的生态分布与树干高度有较显著的关系,树附生苔藓植物的物种丰富度和多样性一般沿垂直高度从下往上呈现递减趋势,在空气湿度较大的树干基部最高。有学者[21,49]也发现,树干朝向虽然对树附生苔藓植物的分布有一定影响,但效果远没有其他主导性环境因子明显。与自然的森林生态系统不同,在人为因素占主导的城市生态系统中,树附生苔藓植物主要受小生境、微气候的影响,人为干扰活动、环境污染、空气湿度、光照、树皮性质、树龄等环境因子的综合作用明显[50],城市中复杂的生境异质性会对树附生苔藓植物产生重大的影响,使它们的分布一般不会单一地倾向于某几个环境因子或某一种生境。
因此,未来在开展城市树附生苔藓植物生态研究中应重点关注小生境,扩大调查范围,兼顾不同生境类型的样点,对环境因子的计测应更加多元化、全面化,以期更加客观、综合地反映树附生苔藓植物分布与环境因子的关系,为保护树附生苔藓植物多样性及其生境提供科学依据。
猜你喜欢
湖北植保(2022年4期)2022-08-23
辽河(2022年1期)2022-02-14
辽河(2022年1期)2022-02-14
安徽农业科学(2021年16期)2021-08-30
土壤(2021年1期)2021-03-23
南方农业·中旬(2020年9期)2020-11-30
中国农村水利水电(2020年4期)2020-06-12
读者·校园版(2018年24期)2018-12-04
绿色科技(2017年15期)2017-09-01
福建农业学报(2016年6期)2016-11-01
