
不同干扰等级下常绿阔叶次生林林分结构及树种多样性
2019-01-24 01:04:52郑小贤
浙江农林大学学报 2019年1期
左 政,郑小贤
(北京林业大学森林资源与环境管理国家林业和草原局重点实验室,北京100083)
生物多样性是人类赖以生存及经济发展的基础[1-2]。然而,随着经济的发展,高强度的森林采伐,导致生物多样性减少,水资源和非木质产品不断损失[3]。生物多样性的损失或许是影响人类生存最重要的因素。林分结构是林分特征中最重要的内容,林分结构的研究对充分发挥森林多项功能,科学经营森林具有重大的意义。因此,开展人为干扰下林分结构及生物多样性的研究具有重要的理论和现实意义。干扰是自然界中普遍存在的、不同时空尺度上偶然发生的不可预知的事件,是自然生态系统演替过程中一个重要的组成部分,直接影响着森林物种组成、群落结构和生物多样性。不同干扰类型和强度对森林群落结构特征有着显著的影响。随着人类活动的加剧,人为干扰对森林生态系统的影响也日益受到重视。国内外关于人为干扰对森林影响做过大量研究,主要集中于物种组成和多样性研究[4-12],并认为中度干扰强度下的物种多样性有增加的趋势,即 “中度干扰假说”,然而干扰与生物多样性的关系非常紧密,也非常复杂[13],并不是所有的干扰对物种多样性的影响都符合该假说所描述的规律[14-18]。常绿阔叶次生林作为中国南方典型的地带性植被类型,具有显著的水源涵养、多样性保护等多项生态功能[19-20]。然而,自20世纪50年代以来,由于森林资源不合理开发利用,人为干扰加剧,南方大部分常绿阔叶林不断减少,退化严重,生态环境问题日益突出。目前,国内外关于常绿阔叶次生林的研究主要集中于林分结构[21-24]、 更新演替[25]、 森林功能[19,26-27]、 群落特征[28-30]、 更新评价[31]、 健康评价[32-33]和经营模式[34-36]等方面,有关人为干扰对林分结构影响研究较少。本研究通过对不同干扰等级下常绿阔叶次生林林分结构及多样性变化的研究,探讨林分结构及多样性变化规律,了解人类活动对其的影响,以期寻求合理的森林经营模式,为常绿阔叶次生林的植被恢复和生物多样性保护提供科学依据。
1 研究地概况
福建省将乐林场位于福建省三明市西北部将乐县城区,处武夷山脉东南麓,扼闽江支流金溪中下游, 介于 26°26′~27°04′N, 117°05′~117°40′E, 东临顺昌, 西接泰宁, 南连明溪, 北毗邵武, 东南与沙县接壤,境内山峰林立,溪流密布,最高峰海拔为1 640.2 m。将乐林场土地总面积为7 113.6 hm2,森林覆盖率为94.8%,有林地面积为6 568.9 hm2。将乐林场属亚热带季风气候,具有海洋性和大陆性气候特点,年平均气温为18.7℃,年平均降水量为1 703.7 mm,最高年度为2 460.4 mm,最低年度仅1 007.8 mm,年际差异较大。将乐林场土壤以红壤分布面积最大,土壤质地一般为沙壤土或轻壤土,块状结构,紧实度适中,比较潮湿。
将乐林场常绿阔叶次生林主要树种为栲树Castanopsis fargesii,赤杨叶Alniphyllum fortune,木荷Schima superba, 甜槠 Castanopsis eyrei, 米槠 Castanopsis carlesii, 青冈 Cyclobalanopsis glauca, 枫香Liquidamba formosana,苦槠Castanopsis sclerophylla,南酸枣Choerospondias axillaria,黄樟Cinnamomum porrectum,冬青Ilex chinensis,光叶山矾Symplocos lancifolia,杨梅Myrica rubra,檵木Loropetalum chinense,杉木Cunninghamia lanceolata,檫木Sassafras tzumu,山矾Symplocos sumuntia等。
2 研究方法
2.1 样地调查
在福建将乐县林场全面踏查的前提下,采用典型样地调查的方法,选取具有代表性地段设置样地25块。这些林分均是1958年皆伐后自然生长起来的常绿阔叶次生林。本研究仅局限于人为干扰,不包括自然干扰。样地形状为矩形,面积为0.04~0.12 hm2,记录样地的基本情况,包括经纬度、海拔、坡度、坡向等,并画出样地的方位示意图。采用相邻格子法将每块样地分成10 m×10 m的调查单元,在每个调查单元内进行每木检尺;在样地的中心和四角分别设置5个5 m×5 m小样方,调查并记录样方内幼苗幼树的树种、直径(树高>1.3 m的测胸径,树高<1.3 m的只测地径)、树高、株数等。样地基本情况见表1。

表1 样地基本情况表Table 1 Basic situation of stand
2.2 干扰评价
在调查样地的同时,对每个样地的干扰情况进行调查分析。根据将乐林场常绿阔叶次生林的实际调查情况,选取相对郁闭度、幼树相对密度、受害木(压弯、折断或掘根)相对密度、受害木相对断面积、伐倒木相对密度和伐倒木相对断面积6个干扰影响因素对常绿阔叶次生林受干扰程度进行定量评价。采用相对影响方法[1],计算6个干扰因素对常绿阔叶次生林干扰程度的相对值。如伐倒木密度中2号样地密度最小,为17株·hm-2,1号,3号和4号样地伐倒木株数分别为50,50和33株·hm-2,则以伐倒木密度最小的2号样地产生干扰的相对值为1,1号、3号和4号样地相对值分别为50/17=2.9,50/17=2.9和33/17=1.9。其余5个干扰影响因素对常绿阔叶次生林产生干扰的相对值依此计算(表2);然后,将6个干扰因素对常绿阔叶次生林干扰强度的相对值的总和作为人为干扰强度值;最后,参考生态干扰度划分系统[37],结合将乐林场常绿阔叶次生林的实际情况,采用等距法依据常绿阔叶次生林干扰影响因素值即干扰度值(FDI),将25块常绿阔叶次生林样地划分为Ⅰ~Ⅴ等5个干扰等级(表3)。

表2 常绿阔叶次生林干扰等级Table 2 Disturbance gradient of evergreen broad-leaved secondary forests

表3 干扰度指数分级Table 3 Hierarchical list of index of disturbance
2.3 数据分析
2.3.1 空间结构 采用体现树种空间隔离程度的树种混交度(Mi),反映林木个体大小的大小比数(Ui),描述林木个体在水平地面上分布格局的角尺度(Wi)3个参数对林分空间结构进行分析[38]。本研究选定相邻木n=4,标准角为α0=72°。计算公式如下:

2.3.2 多样性计算 采用物种丰富度,Margalef丰富度指数,Shannon-Wiener指数,Pielou均匀度指数作为物种α多样性测度指标,用Jaccard相似指数来评价β多样性,Simpson优势度指数作为树种优势度测量指标。①物种丰富度:S=出现在样地内的物种数量。②Margalef物种丰富度指数(DM):DM=(S-1)/lnN。其中:S为样方中物种的数目,N为样方中所有物种个体的数目。③Shannon-Wiener指数(H):其中:Pi为物种i的相对重要值,S为样地的物种总数。④Pielou均匀度指数(E):E=H/lnS。 其中:H为Shannon-Wiener指数,S为样地的物种总数。 ⑤Jaccard相似指数(IS):IS={c/ (a+b+c )}×100。其中:c为2个样方共有的物种数,a为样地1特有的物种数,b为样地2特有的物种数。⑥Simpson优势度指数(C):其中:Pi为物种i的相对重要值,S为样地的物种总数。
3 结果与分析
3.1 不同干扰等级下林分结构特征
不同干扰等级下常绿阔叶次生林林分结构不同(表4)。随着干扰等级的增加,林分平均胸径和平均树高逐渐减小。林分平均密度和幼树密度随着干扰等级的增加先增大后减小,均在干扰等级Ⅳ达到最高值,干扰等级V与其他干扰等级相比,林分密度和幼树密度均为最小。
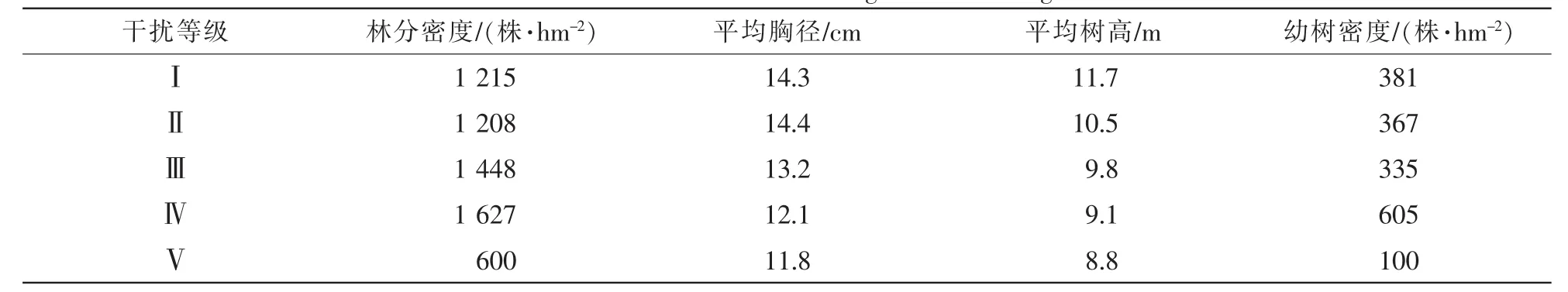
表4 不同干扰等级林分基本情况Table 4 Basic situation of forest along a disturbance gradient
3.2 不同干扰等级下胸径、树高结构
不同干扰等级下常绿阔叶次生林的直径结构差异显著(P<0.05)(图1A),乔木个体在各径阶的数量分布不同。不同干扰等级的径阶分布均为倒 “J”型曲线,即不同干扰等级的直径分布均是随着径级的增大,株数随之减少。在同一干扰等级下,小径阶(DBH<12 cm)个体数显著大于中径阶(12 cm≤DBH<24 cm)和大径阶(DBH≥24 cm)个体数(P<0.05),约占63.89%~78.72%,说明该林分具有潜在的天然更新能力。在同一径阶结构下,干扰等级Ⅴ的小径阶个体数、中径阶个体数和大径阶个体数显著小于其他干扰等级(P<0.05),表明重度干扰等级下,人为的选择性采伐不仅造成大径级树木个体的减少,还因小径阶和中径阶个体数量的减少而降低了林分天然更新能力和生产力。

图1 不同干扰等级下胸径和树高分布Figure 1 Distribution of DBHand tree height along a disturbance gradient
干扰等级Ⅰ~Ⅳ常绿阔叶次生林的树高结构差异显著(P<0.05)(图1B),乔木个体在各高度级的数量分布不同。干扰等级Ⅰ~Ⅳ均为单峰山状曲线,乔木层的高度级峰值均出现在高度级5~7级,并向两侧递减,低高度级(树高<9 m)个体数显著多于中高度级(9 m≤树高<15 m)和高高度级(树高≥15 m)个体数(P<0.05),表明各干扰等级林分垂直结构趋向于稳定。干扰等级Ⅴ各高度级个体数量差异不显著(P>0.05),其分布有2个峰值(5~7级和15~17级),并且缺失最大树高级(高度级19),说明干扰等级Ⅴ树高结构受人为干扰影响较大,林分垂直分布结构不稳定。
3.3 空间结构
不同干扰等级的常绿阔叶次生林随着干扰等级的增加,角尺度逐渐增大(表5)。依据平均角尺度(W)的计算标准,当W>0.517时为团状分布,因此5种干扰类型均属于团状分布,且随着干扰程度的加重,林分越趋向于团状分布。

表5 不同干扰等级空间结构特征Table 5 Structural characteristics of spatial structure along a disturbance gradient
不同干扰等级的常绿阔叶次生林随着干扰等级的增加,大小比数逐渐减小。不同干扰等级的常绿阔叶次生林均处于中庸状态,即每株林木的4株相邻木中有一半比参照树的胸径大。
不同干扰等级的常绿阔叶次生林随着干扰等级的增加,混交度逐渐减小。不同干扰等级的常绿阔叶次生林均趋向于强度混交的状态,即每株林木周围均有2~3株其他树种。一般认为,混交度越大,林分相对越稳定[39]。随着干扰等级的增加,林分稳定性不断降低,不利于林分的健康稳定生长。
3.4 树种多样性
从表6可以看出:不同干扰等级下的多样性指数存在显著差异。中度干扰林分(干扰等级Ⅱ)具有最高的物种丰富度,重度干扰林分物种丰富度最低。轻度干扰林分的Margalef指数,Shannon-Wiener指数及个体物种数均高于其他干扰等级,重度干扰林分Margalef指数,Shannon-Wiener指数及个体物种数最低,其他干扰强度各指数相差不大。Pielou指数随着干扰强度的增大,指数不断减小。总体来看,随着干扰等级的增大,α多样性指数逐渐减小,轻度干扰和中度干扰林分的α多样性指数均高于重度干扰林分。Simpson优势度指数在不同干扰等级下显著变化,重度干扰森林优势度指数最高,轻度干扰森林优势度指数最低。
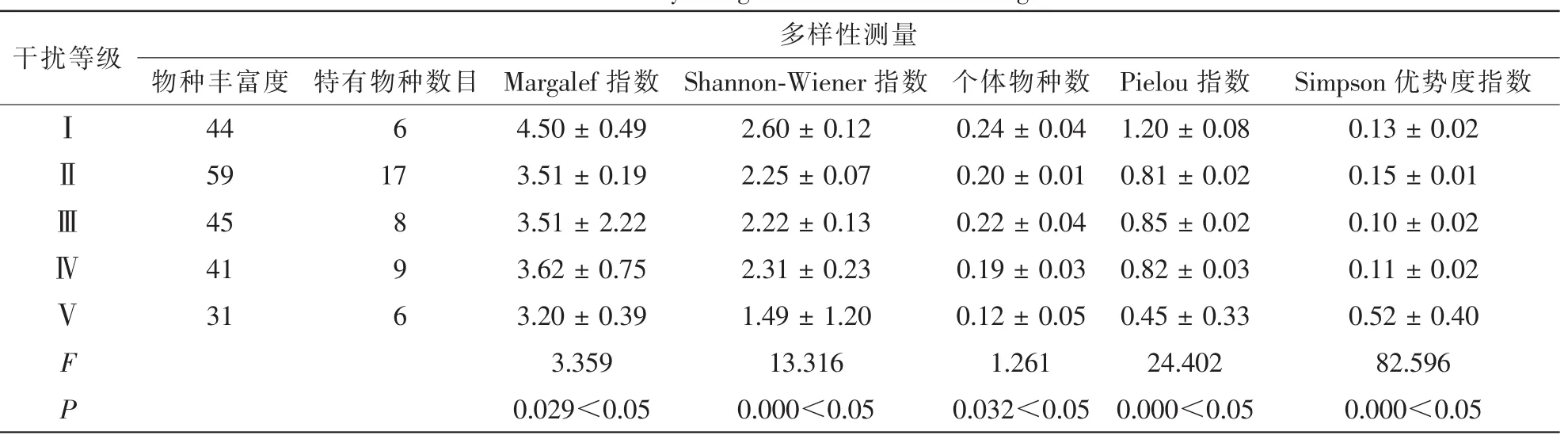
表6 不同干扰等级下α多样性Table 6 α-diversity along a different disturbance gradient
随着干扰等级提升,Jaccard相似指数呈降低的趋势(表7),干扰等级Ⅰ,Ⅱ,Ⅲ,Ⅳ与干扰度等级Ⅴ的Jaccard相似指数都偏低,其中轻度干扰和重度干扰之间物种相似性较低,中度干扰林分之间物种相似性最高。说明干扰强度越大,常绿阔叶次生林物种异质性越强,破碎化也越严重。
4 结论与讨论
干扰是普遍存在于森林生态系统中的正常现象[40-41]。常绿阔叶次生林随着干扰强度的增加,不同干扰等级直径结构和树高结构之间差异显著。不同干扰等级直径分布均趋向于倒 “J”型,且随着干扰等级的增大,森林潜在更新能力和生产力皆随之降低。干扰等级Ⅰ~Ⅳ树高分布均为单峰山状曲线,垂直结构较为稳定,干扰等级Ⅴ有2个峰值且各高度级之间个体数差异不显著,垂直结构不稳定。从林分空间结构分析结果来看,随着干扰强度的增加,林分角尺度逐渐增大,林分均属于团状分布;大小比数逐渐减下,林分均处于平庸状态;混交度逐渐减小,林分均趋向于轻度混交的状态。林分受外界干扰的影响,导致幼树幼苗被压、天窗形成、地形破坏、大径级立木缺失等情况发生,改变了原有群落的组织结构,丧失了这些优势木的庇护作用,扰动林内微环境,使林分处于暂时不稳定状态,影响群落的稳定性和演替。这些同时表明,不同干扰强度对林分结构产生最直接的影响。随着干扰强度的加剧,林分结构退化,森林生态功能衰退,甚至导致物种濒危甚至灭绝,从而间接影响到物种组成。
人为干扰对常绿阔叶次生林的物种多样性产生负面影响。随着干扰等级的增大,林分物种数减少,各多样性指数和均匀度指数均呈现递减的趋势。这与认为适度干扰能够增加物种多样性的 “中度干扰假说”的结论不大一致[42],但与很多其他森林群落乔木物种多样性研究结果一致。这可能与当地民众滥砍盗伐具有较高经济价值的壳斗科Fagaceae优势树种有关,尤其以栲树为主。
重度干扰林分的Simpson优势度指数高于轻度干扰林分。在所有干扰等级林分中,栲树均为优势树种,在重度干扰林分的重要值最高。这与栲树自身耐荫性有关,有利于早期幼苗生长。随着干扰等级的增强,留下的伴生树种多为耐荫树种,能够忍受优势树种与它们进行光照、水分和养料的竞争,同时也与现阶段林场采用封山育林的阔叶林经营措施相关。因此,不同人为干扰强度下呈现不同的物种多样性特征与人为干扰作用范围和物种耐受性、群落繁殖策略等生态习性有关。
由于人为干扰的复杂性导致干扰强度的量化问题研究一直十分困难。本研究加入林分干扰因素,综合建立适当的干扰体系,定量干扰强度,与当地的实际情况较为符合,能够客观反映出常绿阔叶次生林的林分结构及多样性在不同干扰程度下呈现出的规律性。

表7 不同干扰等级下物种相似值Table 7 Jaccard’s coefficient along a disturbance gradient
猜你喜欢
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
