
天然植物提取物替代饲用抗生素的应用研究进展
2019-01-16 03:45:36唐茂妍陈旭东
饲料博览 2018年12期
唐茂妍,陈旭东
(1.上海正正生物技术有限公司,上海 200433;2.北京爱普莱生物科技有限公司,北京 100193)
天然植物提取物在人类药物或营养补充剂中的使用已有数千年历史,中国对药用植物的研究也有悠久的历史。西方把植物作为香料和药物使用开始于两千多年前的罗马和希腊时期[1]。但是天然植物提取物作为饲料添加剂使用是在2000年以后才开始深入研究,特别是在抗生素禁止或限制在饲料中使用之后,欧洲、美国及日本等国家对于天然植物提取物的抗菌作用进行了深入研究。本文综述了天然植物提取物添加剂的抗菌作用、作用机理以及在动物饲料中替代抗生素的应用研究。
1 天然植物提取物的定义及分类
1.1 定义与分类
一直以来植物提取物的概念宽泛而混乱,但作为饲料添加剂使用的植物提取物应符合国家的相关规定和标准。在国内关于植物提取物的标准GB/T 19424-2003中对于天然植物饲料添加剂的定义如下:以一种或多种天然植物全株或其部分为原料,经物理提取或生物发酵法加工,具有提高饲料利用率和改善动物产品品质等功效的饲料添加剂。本文中还沿用习惯说法“植物提取物”,特指从植物中提取(非化工合成)的活性成分明确且可以测定的、含量稳定的、对动物和人类没有任何毒副作用,并已通过动物试验验证可以提高动物生产性能的饲料添加剂。植物提取物按照物理形态可分为固体和液体两种,按照有效成分不同可分为多酚、多糖、萜类、生物碱、黄酮等。
1.2 常见植物提取物
近10年来,植物提取物及其精油在畜牧业生产的应用研究日渐深入,目前已有许多试验数据证明牛至油、肉桂精油、丁香油、迷迭香油、茶树油等,均具有较好的抑制或杀灭细菌、真菌、曲霉菌和球虫的作用。主要植物提取物的活性成分见表1。

表1 植物提取物的活性成分
1.2.1 牛至与牛至油
牛至在植物分类学中属于唇形科、牛至属,又名止痢草、野荆芥,是一种对寒冷敏感的多年生香草,带有甜松和甜橘的香味。牛至含粗蛋白质、多种维生素、微量元素、芳香味挥发油等。从牛至中提取的牛至油含有多种芳香味化合物,其中酚类化合物>50%,主要有效成分为百里香酚、百里香素、香芹酚等。
1.2.2 肉桂与肉桂醛
肉桂属于樟科、樟属,生于亚热带地区,喜温暖潮湿气候的常绿乔木。树皮灰褐色,具有芳香味。肉桂醛存在于肉桂等植物体内,为黄色黏稠液体,是肉桂精油的主要活性成分。自然界中天然存在的或人工合成的肉桂醛均为反式结构。因其分子为一个丙烯醛链接一个苯基,因而可被认为是一种丙烯醛的衍生物。
1.2.3 迷迭香与迷迭香油
迷迭香属唇形科灌木,原产欧洲和非洲北部地中海沿岸,喜温暖气候。迷迭香的花和叶子中能提取优良的抗氧化剂和迷迭香精油。不同来源的迷迭香精油组分大致相同,包括α-蒎烯、1,8-桉叶素、莰烯、樟脑、龙脑和β-蒎烯,但不同来源的迷迭香精油各组分的含量不同[2]。
1.2.4 丁香与丁子香酚
丁香是指桃金娘科蒲桃属的热带植物,原产于印度尼西亚的群岛上。丁子香酚又名丁香酚、丁香油酚,是丁香花蕾中含有的挥发油的主要成分,常温下为无色至淡黄色油状液体,在空气中易变棕色、变稠。几乎不溶于水,溶于大多数有机溶剂。
2 天然植物提取物抑菌机理
迄今为止,植物提取物在抗菌、抗氧化方面的研究取得诸多进展。特别是近年来,植物提取物在畜禽饲料中的应用越来越广泛,其抑菌特性的研究也越来越深入。但因植物提取物的活性成分种类繁多,其抗菌作用也可能有多种机制[4]。不同作用机制可能不是单独进行,相互之间是有影响的;一种机制的反应物或生成物可能会影响另一种机制的运行。
植物提取物的抗菌作用机制包括破坏/降解细胞壁结构、破坏细胞膜蛋白质结构、破坏细胞质膜、使细胞内容物外泄、凝聚细胞质、减弱质子运动力等。
2.1 牛至提取物及其活性成分的抗菌机理
研究发现,具有抑菌活性的植物提取物中酚类化合物如香芹酚、百里香酚和丁子香酚的含量比较高[3-4]。而这些化合物中香芹酚和百里香酚的作用机制研究的最为深入。Stiles等认为,二者的主要作用模式是膜穿孔和膜黏合,细胞膜通透性改变和胞内物质外泄,导致重要的酶系统受到损害,从而引起微生物死亡[5]。研究表明,香芹酚会作用于细胞膜上的磷脂脂肪酸链,从而使细胞膜损伤,引起细胞质内容物外泄造成细胞死亡[6-7]。
植物提取物的抗菌活性与其主要成分的化学结构有关联。Dorman等认为含有酚结构的化合物的抑菌活性高于不含酚结构的化合物[4]。百里香酚和香芹酚的结构相似,化学结构中均含有酚基,只是羟基在苯环上的位置不同。酚羟基是极度活跃基团,极易与相关酶的活性位点形成氢键,从而影响酶的作用。
另外,试验表明植物提取物中不同成分之间具有协同抗菌作用,非主要成分也起重要作用。已经证明香芹酚和百里香酚以及香芹酚和其前体物p-百里香素在抗菌方面存在协同叠加效应[6,8]。完整的植物提取物的抗菌效能明显优于主要成分混合得到的混合物的抗菌效能[9-10]。
2.2 肉桂提取物及其活性成分的抗菌机理
肉桂因其独特的气味和防腐效用很早就被应用到食品、医药和化妆品领域中。后因肉桂活性成分在抑菌方面的高效性和广谱性(细菌、真菌和曲霉菌等),使之在饲料中的应用日渐广泛[11]。
肉桂醛分子中存在一个与苯环共轭的醛基,醛基是亲水基团,易被细菌表面的亲水基吸附而穿入细胞壁,破坏细菌、真菌的细胞壁多糖结构[12]。肉桂醛的醛基还可与细菌中的蛋白及酶作用,破坏细菌正常的生理代谢,抑制细菌生长。通过电镜扫描、胞膜通透性、膜完整性、膜电位实验,发现肉桂醛可破坏菌体细胞的形态,使之出现形变,导致胞膜通透性、完整性受到破坏,影响细胞的代谢活动从而抑制细菌生长[13]。
肉桂醛能作用于葡聚糖合酶和几丁质合酶,特异性抑制真菌细胞壁中葡聚糖和几丁质的合成[14]。谢小梅等研究认为,肉桂醛可通过影响黄曲霉细胞膜麦角甾醇的生物合成,使细胞膜受损而影响其正常功能;同时霉菌细胞不能维持正常的细胞形态,胞内外物质交换受阻,胞内生物大分子合成减少,从而抑制黄曲霉的生长繁殖[15-16]。另外,肉桂醛可作用于黄曲霉的细胞膜,造成膜脂质过氧化,干扰黄曲霉细胞的有序级联反应,破坏线粒体内膜上的氧化还原系统[17]。
2.3 迷迭香提取物及其活性成分的抗菌机理
迷迭香的化学成分包括单萜、二萜、三萜、黄酮等,目前的研究主要集中在二萜类化合物。二萜化合物主要以二萜酚类和二萜醌类为主。二萜酚类包括鼠尾草酸、鼠尾草酚、迷迭香酸、迷迭香酚等化合物,这些物质除具有较强的抗氧化性之外,大部分还具有显著的抑菌作用[18-21]。
关于迷迭香提取物的抑菌机理报道不多,现在的观点认为,二萜和单萜化合物可改变革兰氏阳性菌和阴性菌的细胞膜通透性,引起胞内物质泄漏,使得细胞无法正常代谢,进而减弱了DNA聚合酶活性,影响DNA复制,从而发挥抑菌作用[22-23]。
3 植物提取物抑菌作用及替代饲用抗生素的应用效果
3.1 体外抑菌效果
Hammer等和金立志研究了不同天然植物提取物对有害菌的最小抑菌浓度,见表2[24-25]。研究结果表明,茶树、芫荽、马郁兰和止痢草的提取物对3种致病性细菌(大肠杆菌、沙门氏菌和金黄色葡萄球菌)有较强的抑制效果,尤其是止痢草/牛至提取物的抑菌性最强。
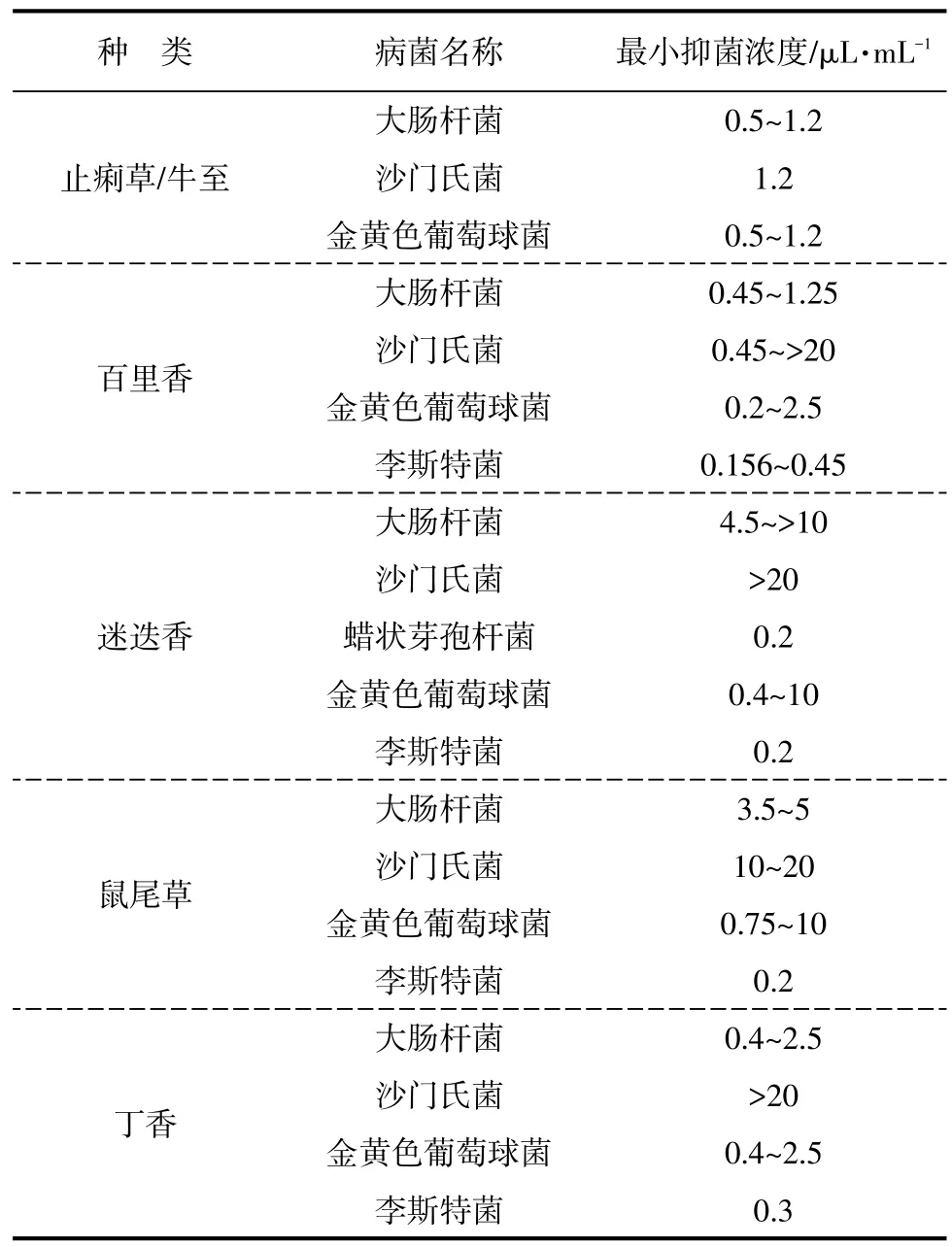
表2 天然植物提取物的最小抑菌浓度
韩乾杰等采用牛津杯法测定肉桂醛、百里香酚和香兰素对于4种细菌(大肠杆菌、沙门氏菌、产气荚膜梭菌和乳酸菌)的抑制效果[26]。结果表明,3种植物精油均对大肠杆菌、沙门氏菌、产气荚膜梭菌有明显抑制作用,而对乳酸菌无抑制作用。在相同浓度下,抑菌效果为肉桂醛>香兰素>百里香酚。肉桂醛与香兰素复合植物精油抑菌效果优于单一植物精油,并且二者质量比为6∶1时抑菌效果最佳。单一丁酸钠没有表现出明显抑菌效果,但丁酸钠与植物精油复合可显著降低植物精油的用量。
3.2 植物提取物替代饲用抗生素的应用进展
虽然2006年起欧盟已禁止抗生素作为生长促进剂添加于饲料中,然而当时国内对于抗生素替代品在实际养殖生产中的应用研究并不多见。2017年硫酸粘杆菌素禁止作为促生长剂在饲料中使用,才使得一系列的抗生素替代品研究逐步深入。这些替代品的研究主要集中于植物提取物、微生态制剂等。
3.2.1 植物提取物在猪饲料中的应用
近年来关于植物提取物替代或部分替代抗生素的试验研究中,大多数表现为正效果,即所用植物提取物能替代特定剂量的某种或某些抗生素,并且不影响或改善动物的生产性能,提高机体免疫力。
纪少丽等在妊娠-哺乳期母猪饲料中添加止痢草精油(OE),发现母猪日粮中添加OE可显著提高仔猪的日增重和日采食量,断奶仔猪到屠宰全程添加OE能显著改善肉猪的日增重。而且母猪日粮添加OE可显著增加断奶至肥育期间的猪血清IGF-Ⅰ水平,并且提高了肉猪屠宰性能[27]。
肖传明等试验证明,在28 d断奶仔猪基础日粮中添加新型猪用植物提取物500 g·t-1可显著降低仔猪腹泻率,提高日增重和日采食量;同时通过测定血液生化指标说明该植物提取物可显著促进断奶仔猪免疫器官发育,增强动物免疫力,可以取代饲料中抗生素的使用[28]。王亮等比较了牛至油提取物、苦木甘草提取物、植物精油复合物和抗菌肽(添加量均为1 000 g·t-1)对断奶仔猪(284 d断奶,大约克夏纯种猪)生产性能、血液生化及免疫性能的影响。以抗生素组作为对照组,试验结果表明,抗生素替代产品能够促进仔猪生长、促进机体蛋白质沉积、降低脂肪沉积并提高机体抵抗力,其中牛至油提取物作用效果最佳[29]。范觉鑫等用几种抗生素替代产品(酸化剂、酶制剂、微生态制剂、植物提取物、中草药添加剂等)组合使用配制无抗日粮,研究在21 d断奶的杜×长×大仔猪(平均体重6.25±0.30 kg)饲料中替代抗生素的效果[30]。发现不同替代品的组合方案对于仔猪的影响差异显著,或者提高了生产性能,或者改善了腹泻率和腹泻指数。但如何兼顾生产性能和腹泻率的配方组合还需进一步研究。田冬冬等也用21 d断奶仔猪(平均体重6.5 kg)试验了几种添加剂组合(复合有机酸+复合酶制剂+微生态制剂+植物提取物)对于仔猪生长性能和抗腹泻效果的影响,结果在整个试验期内,试验组生产性能和腹泻率与抗生素组均无显著差异,表明在该试验条件下,上述组合能替代抗生素在断奶仔猪日粮中使用[31]。杨斌研究发现,饲喂25 d断奶的三元仔猪(7.32±1.16 kg)黄芪提取物(黄芪多糖、黄芪皂苷)42 d,结果与添加抗生素(金霉素+粘杆菌素)对照组相比,日粮中添加黄芪提取物能够显著改善断奶仔猪的饲料转化率,在一定程度上提高断奶仔猪日增重[32]。刘艳霞用品种相同的断奶猪80头(体重8.0±0.5 kg)试验植物源性添加剂艾可特肠佳(主要成分为鞣花酸、木桔素、飞燕草色素-3,5-二糖苷等肠道上皮细胞更新促进剂)和艾可美天然杀菌剂(主要成分是由植物提取物高浓度、高纯度的有机分子单体Cimenol环及柠檬酸组成)对保育猪生长的影响,发现与空白对照组相比,试验组仔猪生长速度、饲料利用率提高,粪便颜色等得到改善,同时保育猪腹泻率大幅度降低(11%和3%)。经济效益分析,保育猪饲料添加艾可特肠佳+艾可美天然杀菌剂,收入增加22.41元·头-1[33]。梁梦玫等用31~33 d三元(杜×大×长)杂交保育猪135头试验不同植物提取物(牛至油、肠佳+艾可美)与抗生素的互作效应,发现降低抗生素用量添加肠佳与艾可美组合产品,可显著提高仔猪日增重并降低料重比[34]。
3.2.2 天然植物提取物在禽饲料中的应用
Zhang等试验表明,常山提取物可有效对抗艾美耳球虫感染,特别是添加20 mg·kg-1时,可显著提高肉鸡体增重,降低出血性腹泻、病变评分和虫卵排出量,效果与抗球虫药地克珠利相当[35]。Liu等发现,肉鸡饲料添加植物提取物(辣椒素、大蒜素和姜黄素)均可降低大肠杆菌攻毒时腹泻评分和腹泻频率,并且攻毒后当天的TNF-α、结合珠蛋白、回肠巨噬细胞、中性粒细胞数量以及攻毒11 d的白细胞和中性粒细胞数量均显著降低[36]。
史东辉等研究了止痢草提取物对肉鸡血清的抗氧化作用和鸡肉脂类氧化影响,结果表明,整个试验期添加止痢草提取物的各试验组肉鸡血清总抗氧化能力(T-AOC)均显著高于空白对照组和抗生素组[37]。所有试验各组整个贮藏期间肌肉中丙二醛浓度均显著低于空白对照组和抗生素组,可见,日粮中的止痢草植物提取物可提高肉鸡血清中抗氧化酶的活性,有效延缓鸡肉中脂类的氧化。肖传明等比较了植物提取物(含蒲公英等提取物,富含黄酮、生物碱等有效成分)和抗生素(10%硫酸粘杆菌素200 g·t-1+10%金霉素预混剂400 g·t-1)对Cobb商品代肉鸡生长性能和血清生化指标的影响,结果表明,与抗生素组相比,植物提取物组显著提高肉鸡血清总蛋白含量、谷草转氨酶、淀粉酶、脂肪酶活力,以及钙磷、粗蛋白质、粗脂肪消化率,并显著降低死淘率[38]。
杨静静等发现,对于经口感染柔嫩艾美耳球虫孢子卵囊的肉鸡,复合植物提取物与阳性对照组(地克珠利)相比疗效显著,且在饲料中添加复合植物提取物Ⅰ的增重效果优于地克珠利,是替代抗球虫、促生长抗生素的良好药物选择[39]。Abudabos等研究结果表明,在鼠伤寒沙门氏菌攻毒的肉鸡饲料中添加复合植物提取物,与抗生素组相比改善了饲料利用率,其他生产性能指标没有显著差异;血液生化指标和肝脏酶活均无显著差异,表明此提取物可在肉鸡日粮中替代抗生素[40]。
3.2.3 天然植物添加剂在水产饲料中的应用
刘泓宇等以基础饲料为空白对照,添加氟苯尼考(Florfenicol)0.3%为抗生素对照组,其余4组分别添加RapXOL(植物提取物组,主要成分为牛至油、肉桂醛等)200 mg·kg-1以及不同梯度含量益生菌,发现日粮中添加植物提取物可明显提高凡纳滨对虾的成活率、全虾蛋白含量。植物提取物RapXOL能提高对虾的成活率和免疫能力,可作为良好的免疫增强剂[41]。
4 植物提取物应用中存在的问题
虽然植物提取物作为一种新型高效的饲料添加剂越来越多的应用于畜禽养殖中,并且多数试验报告表明其在替代饲料抗生素促生长剂方面有巨大潜力,但是在实际应用中也存在问题,影响了植物提取物在养殖业中的使用效果和范围。
4.1 产品质量不稳定
虽然我国天然植物种类十分丰富,但是植物品种、种植地、收获时间、使用部位等不同会导致其活性成分差异大,即使植物名称相同,其主要活性成分的含量也可能不同[1,25]。因而对于同种来源的植物提取物其质量差异可能很大。
4.2 产品标准不统一
在植物提取物方面的国家标准和行业标准缺失。主要原因在于:天然植物成分复杂,不同地域的植物有效成分含量不同,因而对于原料本身的标准难以制定;不同厂家设计植物提取物配方时参考的饲养标准也不同;植物提取物活性物质不明确或含量不稳定,导致政府部门无法监督,用户也无法进行检测,限制了植物提取物的评估和应用。
4.3 实际应用效果不确定
虽然很多植物提取物在体外对大多数消化道有害菌均有抑制作用,但在动物养殖中使用时,效果往往千差万别,并且有时体外抑菌剂量并不能在体内达到同样的效果。另外,不同植物提取物产品的有效成分和含量不同,对于有害菌的抑制效果差异很大。因而,针对性使用植物提取物能降低抗生素生长促进剂在动物饲料中的应用,但还需要更深入研究。
猜你喜欢
源流(2023年5期)2023-07-01 14:07:24
今日农业(2022年2期)2022-11-16 12:29:47
中老年保健(2022年2期)2022-08-24 03:20:44
中老年保健(2021年5期)2021-12-02 15:48:21
现代畜牧科技(2021年2期)2021-03-19 07:48:26
河南畜牧兽医(2020年11期)2020-01-11 05:31:49
现代营销(创富信息版)(2018年2期)2018-08-15 00:45:27
中成药(2017年10期)2017-11-16 00:50:27
广东饲料(2016年4期)2016-12-01 03:43:18
广西林业科学(2016年1期)2016-03-20 05:33:05
